
Abstract
Steele, Andrew R., Philip N. Ainslie, Rachel Stone, Kaitlyn Tymko, Courtney Tymko, Connor A. Howe, David MacLeod, James D. Anholm, Christopher Gasho, and Michael M. Tymko. Global REACH 2018: characterizing acid–base balance over 21 days at 4,300 m in lowlanders. High Alt Med Biol. 23:185–191, 2022.
Introduction:
High altitude exposure results in hyperventilatory-induced respiratory alkalosis, followed by metabolic compensation to return arterial blood pH (pHa) toward sea level values. However, previous work has limited sample sizes, short-term exposure, and pharmacological confounders (e.g., acetazolamide). The purpose of this investigation was to characterize acid–base balance after rapid ascent to high altitude (i.e., 4,300 m) in lowlanders. We hypothesized that despite rapid bicarbonate ([HCO3−]) excretion during early acclimatization, partial respiratory alkalosis would still be apparent as reflected in elevations in pHa compared with sea level after 21 days of acclimatization to 4,300 m.
Methods:
In 16 (3 female) healthy volunteers not taking any medications, radial artery blood samples were collected and analyzed at sea level (150 m; Lima, Peru), and on days 1, 3, 7, 14, and 21 after rapid automobile (∼8 hours) ascent to high altitude (4,300 m; Cerro de Pasco, Peru).
Results and Discussion:
Although reductions in [HCO3−] occurred by day 3 (p < 0.01), they remained stable thereafter and were insufficient to fully normalize pHa back to sea level values over the subsequent 21 days (p < 0.01). These data indicate that only partial compensation for respiratory alkalosis persists throughout 21 days at 4,300 m.
Introduction
There are several important physiological changes that occur to facilitate successful acclimatization in individuals of lowland descent (i.e., born <1,500 m) to high altitude. Among the first and most significant responses to high altitude is an increase in alveolar ventilation. Although this hypoxia-induced hyperventilation attenuates the decrease in arterial blood oxygenation, it causes a concomitant decrease in the partial pressure of arterial carbon dioxide (PaCO2). Respiratory-induced alkalosis results in an obligatory excretion of bicarbonate ([HCO3−]) following hypocapnia to “normalize” arterial blood pH (pHa) back to sea level values.
Several studies have assessed acid–base balance at high altitude; however, the timeline of this response during early acclimatization is not well understood, and only few studies have assessed acid–base balance after several weeks of high altitude exposure (i.e., 3,000–5,000 m) (Severinghaus et al., 1963; Forster et al., 1975; Dempsey et al., 1978; Krapf et al., 1991; Lundby et al., 2004; Somogyi et al., 2005; Subudhi et al., 2014; Steele et al., 2020; Bird et al., 2021).
Early accounts of incomplete metabolic compensation in response to respiratory alkalosis have been reported since 1963 where four healthy participants were taken rapidly to 3,800 m (Severinghaus et al., 1963). Arterial pH rose from 7.42 to 7.49 after 2 days and remained elevated to a similar extent after 8 days (Severinghaus et al., 1963). There have also been a number of other studies that have reported arterial (or arterialized venous blood) acid–base balance after rapid ascent to high altitude with a minimum of 5 days of altitude exposure, but typically limited to <10 days at high altitude (Forster et al., 1975; Dempsey et al., 1978; Krapf et al., 1991; Lundby et al., 2004; Somogyi et al., 2005; Subudhi et al., 2014; Steele et al., 2020; Bird et al., 2021). Consistent across these studies, as outlined in Table 1, there is a decrease in partial pressure of arterial oxygen (PaO2), PaCO2, and [HCO3−] and an increase in pHa. However, a temporal characterization of acid–base balance over several weeks at high altitude is yet to be established.
Summary of Studies That Investigated Acid–Base Balance at High Altitude in Lowlanders
All information was taken from the reported mean data. These delta changes are from baseline.
Denotes that pHa was not reported in this article. We calculated pHa based on the physiological principles of the Henderson–Hasselbalch equation.
Denotes that [HCO3−] was not reported in this article. We calculated [HCO3−] based on the physiological principles of the Henderson–Hasselbalch equation.
[HCO3−], bicarbonate; F, female; NR, not reported value; PaCO2, partial pressure of arterial carbon dioxide; PaO2, partial pressure of arterial oxygen; pHa, arterial blood pH; RR, renal reactivity.
The purpose of this investigation was to fully characterize acid–base balance at high altitude by taking serial measurements of arterial blood on days 1, 3, 7, 14, and 21 following rapid ascent to 4,300 m in healthy lowlanders. We hypothesized that we would observe a rapid reduction of [HCO3−] within 1 week of acclimatization; however, incomplete metabolic compensation would still be apparent as reflected in elevations in pHa compared with sea level after 21 days of acclimatization to 4,300 m.
Materials and Methods
Ethical approval
The data presented were collected in accordance with the Declaration of Helsinki (except registry in a database). The study was approved by University of British Columbia Clinical Research Ethics Board (H17-02687), and local Peruvian ethics committee for the Universidad Peruana Cayetano Heredia (No. 101686). All participants signed the approved consent form after the study was thoroughly explained and were free to withdraw from experimentation without justification or penalty at any time. An overview of our research teams high altitude expedition to Peru can be found elsewhere (Tymko et al., 2020), and the blood gas data at sea level and on day 14 of high altitude exposure have been previously published (Tymko et al., 2020). However, the a priori, primary research question addressed in the current study is novel and exclusively dealt within this study alone.
Participants
All participants included in this investigation were born and currently reside below 1,500 m and avoided visiting altitudes >1,500 m for at least 6 months before the expedition. Before experimentation, participants were asked to refrain from exercise and caffeine for a minimum of 12 hours, and to be fasted for a minimum of 2 hours. Study participants were included if they were between the ages of 18 and 60 years without any medical history of cardiovascular, cerebrovascular, pulmonary, or metabolic disease. Participants were studied at 150 m (Lima, Peru; sea level), and on days 1, 3, 7, 14, and 21 at 4,300 m (Cerro de Pasco, Peru) after rapid transport (∼8 hours) by automobile.
Participants were allowed to drink ad libitum, and no dietary restrictions were implemented for this specific experiment. In addition, all participants were asked to refrain from taking acetazolamide (i.e., diamox) and nonsteroidal anti-inflammatory agents throughout both the ascent and stay at high altitude. However, a few participants required acetazolamide treatment early into our high altitude exposure to reduce severe symptoms of high altitude illness; however, these participants were omitted from data collection until they ceased taking medication for at least 3 days.
Blood sampling and analysis
Participants were supine breathing quietly for a minimum of 10 minutes before experimentation. Arterial blood was drawn from the radial artery directly into a safePico auto-fill syringe (Radiometer, Copenhagen, Denmark) for immediate blood gas analysis, and into lithium–heparin containing vacutainers for blood viscosity measurements on day 14 of acclimatization. In addition, on day 14, blood samples were analyzed using a stationary blood analyzer to measure hematocrit (Hct) and hemoglobin ([Hb]) (ABL90; Radiometer, Mississauga, Ontario).
On each testing day, arterial blood samples were analyzed by a portable blood analyzer for arterial oxygen saturation (SaO2), pHa, PaCO2, PaO2, and [HCO3−] (CG4+ Cartridge; i-STAT, Abbot Point of Care, Princeton, NJ), which has an internal barometric sensor to correct for prevailing atmosphere pressure. Following collection into lithium–heparin vacutainers, whole blood viscosity was measured in most, but not all, samples in duplicate at a shear rate of 225 seconds−1 and standardized temperature of 37°C using a cone and plate viscometer (DV2T Viscometer; Brookfield Amtek, USA) (Gnasso et al., 2001; Baskurt et al., 2009).
Statistics
All statistical analyses were performed using GraphPad Prism V8.0 (San Diego, CA) and were reported as mean ± standard deviation. Statistical significance was set at p < 0.05. One-way repeated measures mixed effects model with the Geisser–Greenhouse correction were used to detect statistical differences within participants. When significant F ratios were detected, post hoc comparisons were made using the Tukey test. The following calculations were also used to estimate hemoglobin–oxygen affinity (i.e., p50) (Hill, 1910), arterial oxygen content (CaO2), calculated alveolar ventilation, hydrogen ion concentration [H+], and renal reactivity (RR):
Results
Arterial blood data in lowlanders after ascent to 4,300 m
Table 2 and Figure 1 display arterial blood data collected in lowlanders after rapid ascent to 4,300 m from 150 m. At 4,300 m, PaO2 and SaO2 were reduced (p < 0.01 all time points), and pHa was elevated on days 1, 7, 14, and 21 compared with sea level (all p < 0.02). Calculated alveolar ventilation was elevated at all high altitude time points compared with low altitude (all p < 0.01); however, calculated alveolar ventilation on day 1 was lower compared with days 7, 14, and 21 at high altitude (p = 0.032, p = 0.018, and p < 0.01, respectively). The PaCO2 was reduced at all high altitude time points compared with low altitude (all p < 0.01) but was higher on day 1 compared with days 7, 14, and 21 (all p < 0.02).
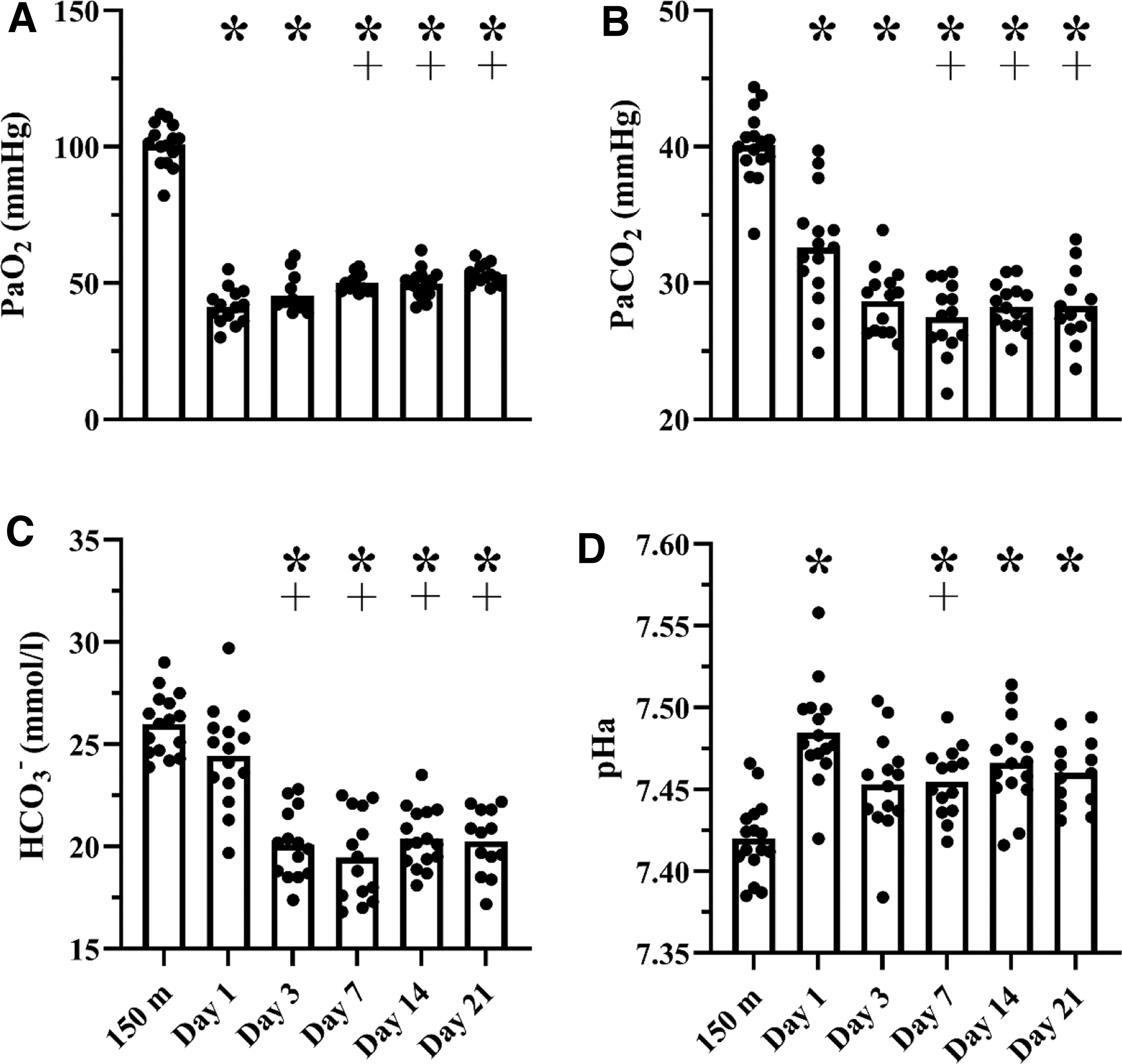
Arterial blood data in lowlanders after rapid ascent to 4,300 m. Individual data in lowlanders displaying the effect of high altitude exposure to 3,800, 4,300, and 5,000 m on arterial blood gases (panels A+B), [HCO3−] (Panel C), and pHa (Panel D). *p < 0.05 versus 150 m; ┼p < 0.05 versus day 1 at 4,300 m. One-way repeated measures mixed effects model with the Geisser–Greenhouse correction were used to detect statistical differences within participants. When significant F ratios were detected, post hoc comparisons were made using the Tukey test. [HCO3−], bicarbonate; PaCO2, partial pressure of arterial carbon dioxide; PaO2, partial pressure of arterial oxygen; pHa, arterial blood pH.
Arterial Blood Data in Lowlanders After Rapid Ascent from 150 to 4,300 m
One-way repeated measures mixed effects model with the Geisser–Greenhouse correction were used to detect statistical differences within participants. When significant F ratios were detected, post hoc comparisons were made using the Tukey test.
p < 0.05 versus low altitude.
p < 0.05 versus day 1.
p < 0.05 versus day 3.
[H+], hydrogen concentration; [Hb], hemoglobin; BMI, body mass index; CaO2, arterial oxygen content; Hct, hematocrit; p50, partial pressure of oxygen to achieve 50% hemoglobin saturation; PATM, atmospheric pressure; SaO2, arterial oxygen saturation.
The observed respiratory alkalosis was not fully compensated by the rapid reduction in [HCO3−] that was stable by day 3 at high altitude (all p < 0.01). [Hb], Hct, CaO2, and blood viscosity were elevated on day 14 compared with sea level (all p < 0.05), and p50 remained unchanged at high altitude (p = 0.51). In addition, RR was lower on day 1 compared with days 3, 7, 14, and 21 (all p < 0.01). Figure 2 displays the Davenport diagram illustrating acid–base buffering at high altitude. The initial rightward shift on day 1 (i.e., increase in pHa) had not fully returned pHa to sea level values despite substantial renal [HCO3−] excretion by day 3.
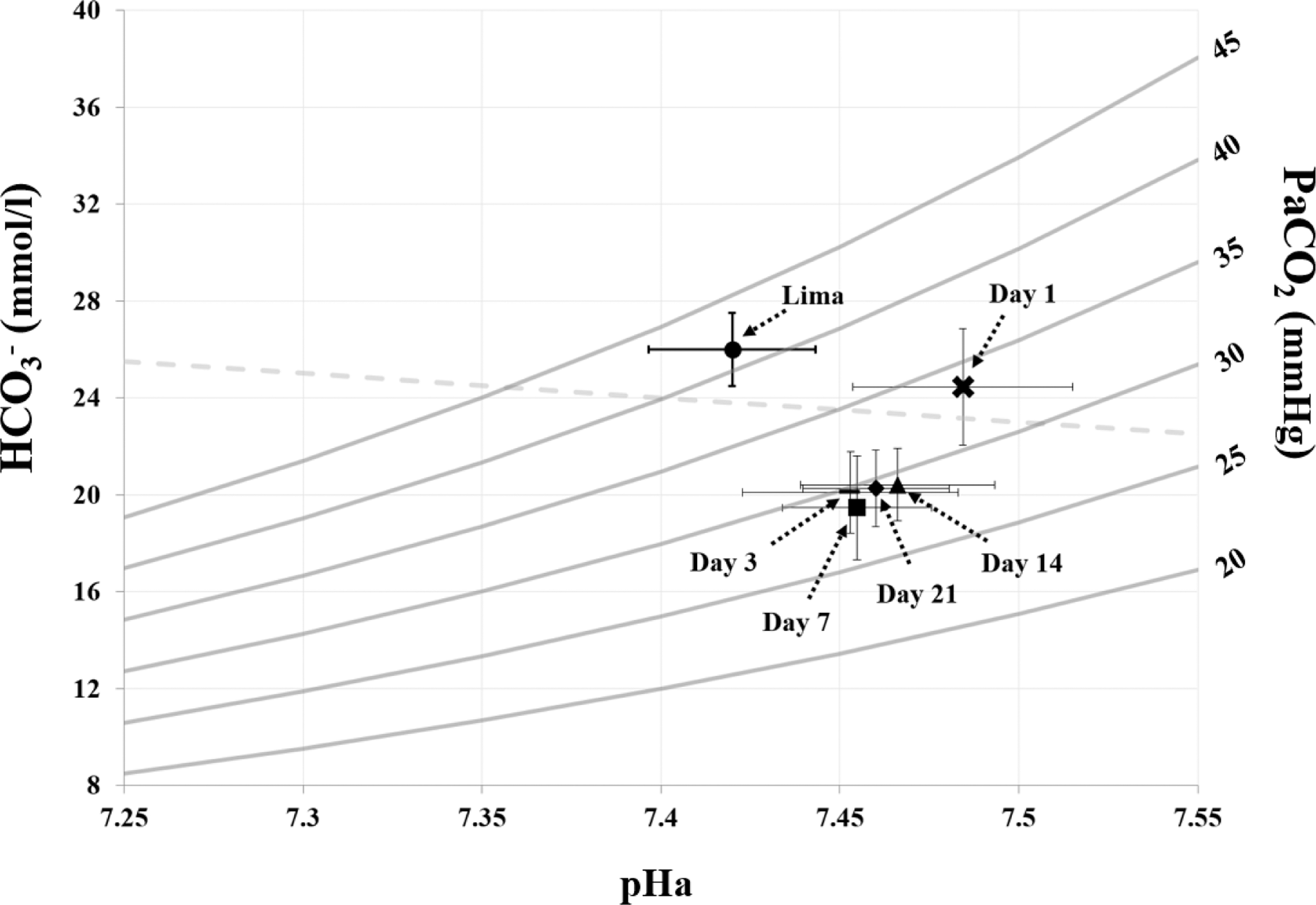
The Davenport diagram in lowlanders after rapid ascent to 4,300 m. Mean ± standard deviation are represented for data collected in Lima and on days 1, 3, 7, 14, and 21 at 4,300 m. The respiratory alkalosis on day 1 resulted in a rightward shift in acid–base equilibrium, which was corrected (i.e., downward and leftward shift) via reductions in arterial [HCO3−], and acid–base balance remained stable between days 3–21 at 4,300 m.
Discussion
The purpose of the current investigation was to characterize acid-base regulation in lowlanders in the absence of high altitude medications at 4,300 m by taking serial measurements of arterial blood over 21 days. We found that pHa was elevated immediately on day 1 of arrival to 4,300 m and remained elevated above sea level values despite rapid metabolic compensation (i.e., [HCO3−] excretion) by day 3 of acclimatization. The clinical relevance of sustained elevated pHa at high altitude remains unclear, and the following discussion outlines the implications of these finding in the context of acid–base physiology, and acclimatization to high altitude.
Acid–base balance in lowlanders acclimatizing to high altitude
We have demonstrated similar findings compared to the majority of rapid ascent high altitude studies investigating acid–base balance for a minimum of 5 days (average duration at high altitude = 13.3 ± 14.6 days) (Severinghaus et al., 1963; Forster et al., 1975; Dempsey et al., 1978; Krapf et al., 1991; Lundby et al., 2004; Somogyi et al., 2005; Subudhi et al., 2014; Steele et al., 2020; Bird et al., 2021). Similarly, our data demonstrated that at high altitude, pHa was elevated early during acclimatization and remained elevated for 21 days, suggesting an incomplete metabolic compensation. Also, we observed that PaCO2, [HCO3−], and pHa were stable after only 3 days of high altitude exposure in lowlanders (Table 2; Figs. 1 and 2). Whether pHa remains elevated with longer exposure in lowlanders (i.e., months) or is reflective of a new stable acid–base balance remains to be elucidated.
RR, an index of acid–base compensation (Δ[HCO3−]/Δ[PaCO2]), reflects the relationship between the renal stimulus (i.e., PaCO2) and the physiological correction [HCO3−] (Zouboules et al., 2018). We found that after 3 days of high altitude there was no change in RR. This would be expected since we observed no further changes in [HCO3−] or PaCO2 beyond day 3 at high altitude.
Despite the several strengths of the current investigation, there are some methodological considerations. First, the Davenport diagram (Fig. 2) only provides basic insight into acid–base derangements at high altitude. Therefore, one of our research team's future goals is to collect additional data for more sophisticated models (e.g., Stewart's model, base excess, and ion equilibrium) to better understand acid–base balance at high altitude (Stewart, 1983; Corey, 2003). Second, high altitude causes a decrease in plasma volume and an increase in erythrocyte proliferation (D'Alessandro et al., 2016; Young et al., 2019), which was supported by the observed increases in hematocrit (Table 2). These changes in hydration, plasma, and extracellular volume status would undoubtedly affect renal function and metabolic compensation.
Third, our study cohort did not consist of enough female participants to make meaningful sex-related comparisons, however, we acknowledge that acid–base regulation between males and females may be different due to ovarian hormones and iron status (Cárdenas-Santamaría et al., 2018; Ryan et al., 2021). Fourth, salt and fluid intake was not controlled for during testing. We acknowledge that changes in fluid and salt may have contributed to changes in renal function and electrolyte balance (Swenson et al., 1995). Fifth, the most novel aspect of this study is the high frequency of arterial blood samples taken while at high altitude; however, based on these data, more rapid measurements (e.g., every 6–12 hours) of arterial blood during the first 3 days of altitude exposure would give a more detailed timeline on acute acid–base acclimatization to high altitude.
Several additional questions remain after completing this study. Based on the current presented data and a report by Lundby et al. (2004) that assessed acid–base balance after 8 weeks at a similar altitude (Lundby et al., 2004), pHa does not return to sea level values. However, whether pHa returns to sea level values over a longer time period in lowlanders (e.g., 4–6 months) remains to be tested, and whether sustained elevations in pHa are adaptive or maladaptive remains to be elucidated. Furthermore, it is possible that an altitude threshold exists for acid–base balance, where above a certain altitude pHa can no longer return to sea level values. More laboratory-based and field-based experiments are required to address these current gaps in the literature.
Conclusions
We found that pHa in lowlanders was elevated and full metabolic compensation was absent even after 3 weeks residing at high altitude. These data provide unique insight into acute and chronic respiratory and metabolic adaptation to hypoxemia.
Footnotes
Authors' Contributions
A.R.S., M.M.T., and P.N.A were responsible for the concept of the study. All authors contributed to the analysis, interpretation of the data, along with drafting the article or critically revising it for important intellectual content. All authors approved the final version of the article, and all persons designated as authors qualify for authorship, and all those who qualify for authorship are listed.
Availability of Data and Material
The data sets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Author Disclosure Statement
We have no competing interests to declare.
Funding Information
These research expeditions were funded, in part, by a Canada Research Chair (CRC) and the Natural Sciences and Engineering Research Council (NSERC) to P.N.A. M.M.T. was funded by the NSERC graduate award.