
Abstract
The lentiviral vector system based on human immunodeficiency virus type 1 (HIV-1) is used extensively in gene therapy trials of neurological and neurodegenerative diseases. Retrograde axonal transport of viral vectors offers a great advantage to the delivery of genes into neuronal cell bodies that are situated in regions distant from the injection site. Pseudotyping of HIV-1-based vectors with selective variants of rabies virus glycoprotein (RV-G) increases gene transfer via retrograde transport into the central nervous system. Because large-scale application for gene therapy trials requires high titer stocks of the vector, pseudotyping of a lentiviral vector that produces more efficient retrograde transport is needed. In the present study, we developed a novel vector system for highly efficient retrograde gene transfer by pseudotyping an HIV-1 vector with a fusion envelope glycoprotein (termed FuG-B) in which the cytoplasmic domain of RV-G was substituted by the corresponding part of vesicular stomatitis virus glycoprotein. The FuG-B pseudotype shifted the transducing property of the lentiviral vector and enhanced the retrograde transport-mediated gene transfer into different brain regions innervating the striatum with greater efficiency than that of the RV-G pseudotype in mice. In addition, injection of the FuG-B-pseudotyped vector into monkey striatum (caudate and putamen) allowed for highly efficient gene delivery into the nigrostriatal dopamine system, which is a major target for gene therapy of Parkinson's disease. Our strategy provides a powerful tool for the treatment of certain neurological and neurodegenerative diseases by promoting retrograde gene delivery via a lentiviral vector.
Introduction
In the present study, we generated two kinds of fusion envelope glycoproteins composed of different combinations of the N-terminal and C-terminal segments derived from RV-G and VSV-G and tested them for the production of an HIV-1-based vector. The glycoprotein containing the extracellular and transmembrane domains of RV-G fused to the cytoplasmic domain of VSV-G (termed FuG-B) increased the vector titer in different cell lines and resulted in highly efficient gene transfer via retrograde transport into various brain regions in the rodent and primate. Our lentiviral system with FuG-B can enhance retrograde gene delivery in gene therapy trials for various neurological and neurodegenerative diseases.
Materials and Methods
Plasmid construction
The envelope plasmid (pCAGGS-RV-G) contained RV-G cDNA under control of the cytomegalovirus enhancer/chicken β-actin promoter (Niwa et al., 1991). RV-G cDNA was isolated from the challenge virus standard strain of rabies virus passaged in suckling mouse brain (Morimoto et al., 1998). The domain structures of VSV-G and RV-G were predicted by a computer algorithm (Rose et al., 1982). Envelope plasmids encoding two kinds of fusion glycoproteins (FuG-A and FuG-B) were constructed. FuG-A cDNA encoded a chimeric protein containing the extracellular domain of RV-G fused to the transmembrane and cytoplasmic domains of VSV-G, whereas the FuG-B cDNA encoded a protein containing the extracellular and transmembrane domains of RV-G fused to the cytoplasmic domain of VSV-G. The RV-G cDNA in the pCAGGS-RV-G was exchanged for the fusion glycoprotein cDNAs, resulting in the envelope plasmids pCAGGS-FuG-A and pCAGGS-FuG-B.
Viral vector production
DNA transfection and viral vector preparation were performed as described previously (Hanawa et al., 2002, 2004) with some modifications. The transfer plasmid (pCL20c-MSCV-GFP) contained the cDNA encoding enhanced green fluorescent protein (GFP) downstream of the murine stem-cell virus (MSCV) promoter. Human embryonic kidney (HEK) 293T cells were transfected with transfer, envelope, and packaging plasmids by the calcium phosphate precipitation method. Viral vector particles were pelleted by ultracentrifugation at 50,000 g for 90 min and resuspended in phosphate-buffered saline (PBS). For injection, vector particles were centrifuged at 10,000 g for 16–18 hr and resuspended in PBS. The particles were then applied to a Sepharose Q FF ion-exchange column (GE Healthcare, Little Chalfont, UK) in PBS and eluted with a linear 0.0–1.5 M NaCl gradient. The fractions were monitored for absorbance at 260/280 nm. The peak fractions containing the particles were collected and concentrated by centrifugation through a Vivaspin filter (Vivascience, Lincoln, UK).
For measurement of the titer of vector preparations, HEK293T, Neuro2A, and N1E-115 cells were seeded into 10-cm-diameter tissue culture dishes and transduced with proper concentrations of viral vectors, and the functional titer was estimated by flow cytometry (FACSCalibur, Nippon Becton Dickinson Co., Tokyo, Japan). In addition, viral RNA in vector stock solution was isolated with a NucleoSpin RNA virus kit (Clontech, Mountain View, CA), and the copy number of the RNA genome was determined using a Lenti-X quantitative reverse transcription–polymerase chain reaction (PCR) titration kit (Clontech). PCR amplification was performed on duplicate samples using the StepOne real-time PCR system (Applied Biosystems, Tokyo) under the following conditions: one cycle of 95°C for 3 min; and 40 cycles of 95°C for 15 sec and 54°C for 1 min.
Vector injection
Animal care and handling procedures were conducted according to the guidelines established by the Animal Research Committee of Fukushima Medical University, Fukushima, Japan, and by the Animal Care and Use Committee of Primate Research Institute, Kyoto University, Inuyama, Japan. Thirty-eight C57BL/6J mice (12 weeks old) and four crab-eating monkeys (Macaca fascicularis, weighing 3.2–4.0 kg) were used for the present study.
For stereotaxic surgery, mice were anesthetized with sodium pentobarbital (50 mg/kg, i.p.), and vectors were introduced into the dorsal region of the striatum (1.0 μl per site, two sites on a track), nucleus accumbens (0.5 μl per site, one site), or medial prefrontal cortex (mPFC) (0.5 μl per site, one site) through a glass microinjection capillary connected to a microinfusion pump. The anteroposterior/mediolateral/dorsoventral coordinates, respectively, from bregma and dura were 0.5/2.0/2.5 mm and 0.5/2.0/3.2 mm for intrastriatal injection, 1.2/1.0/4.2 mm for intra-accumbens injection, and 2.1/0.2/1.7 mm for intra-mPFC injection, according to an atlas of the mouse brain (Paxinos and Franklin, 2001).
Monkeys were sedated with ketamine hydrochloride (5 mg/kg, i.m.) and xylazine hydrochloride (0.5 mg/kg, i.m.) and then anesthetized with sodium pentobarbital (20 mg/kg, i.v.). With the aid of a magnetic resonance imaging-guided navigation system (StealthStation TRIA [Medtronic, Minneapolis, MN] and Brainsight Primate [Regue Research, Montreal, Canada]), viral vectors were injected through a 50-μl Hamilton microsyringe into the striatum (7.5 μl per site, two sites per track, seven tracks per monkey: three tracks for the caudate nucleus and four tracks for the putamen) according to an atlas of the monkey brain (Szabo and Cowan, 1984).
Histological analysis
Four weeks after the injection, mice were anesthetized with sodium pentobarbital (50 mg/kg, i.p.) and perfused transcardially with 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) for mice or with a mixture of 10% formalin and 15% picric acid in 0.1 M phosphate buffer for monkeys. For immunostaining by the avidin–biotin–peroxidase complex method, transverse sections (30 μm thick for mice, 60 μm thick for monkeys) were incubated with rabbit polyclonal anti-GFP antibody (Molecular Probes, Eugene, OR) at a 1:2,000 dilution and then with biotinylated goat anti-rabbit IgG antibody (Vector Laboratories, Burlingame, CA) at a 1:500 dilution. The immunoreactive signals were visualized with a Vectastain Elite ABC kit (Vector Laboratories). For double immunofluorescence histochemistry, sections were incubated with rabbit polyclonal anti-GFP antibody (1:2,000 dilution) and one of the following mouse monoclonal antibodies: anti-NeuN antibody (1:400 dilution, Chemicon, Temecula, CA), anti-glial fibrillary acidic protein (GFAP) antibody (1:400 dilution, Sigma, St. Louis, MO), or anti-tyrosine hydroxylase (TH) antibody (1:100 dilution, Chemicon). Sections were then incubated with fluorescein isothiocyanate–conjugated goat anti-rabbit IgG and Cy3-conjugated donkey anti-mouse IgG (1:500 dilution, Jackson ImmunoResearch Laboratories, West Grove, PA). Fluorescent images were taken under a confocal laser scanning microscope (model LSM510, Zeiss, Thornwood, NY) equipped with proper filter cube specifications for fluorescein isothiocyanate and Cy3 fluorescence channels.
For cell counts, a series of sections through the forebrain and midbrain were used for immunostaining by the avidin–biotin–peroxidase complex method. The number of immunostained cells in each brain region was counted with a computer-assisted imaging program (NIH Image version 1.62, National Institutes of Health, Bethesda, MD). For characterization of striatal cells around vector injection sites, representative sections were used for double immunofluorescence histochemistry. In individual animals, the number of immunostained cells in the regions of interest (0.2 × 0.2 mm) was counted with the imaging program. Eight to 10 sections obtained from each brain region of individual animals were used for cell counts, and the average per section was calculated.
Statistical analysis
Analysis of variance (ANOVA) and post hoc Tukey's HSD test were used for statistical comparisons. Data are expressed as mean ± SEM values.
Results
Creation of lentiviral vectors pseudotyped with fusion envelope glycoproteins
We constructed the envelope plasmids encoding two kinds of fusion glycoproteins, designated FuG-A and FuG-B (Fig. 1A). FuG-A consisted of the extracellular domain of RV-G fused to the transmembrane and cytoplasmic domains of VSV-G, whereas FuG-B contained the extracellular and transmembrane domains of RV-G fused to the VSV-G cytoplasmic domain. We produced HIV-1-based lentiviral vectors carrying the GFP transgene by pseudotyping with distinct glycoproteins RV-G, FuG-A, or FuG-B, transduced vector preparations into HEK293T cells, and then measured the functional titer (transducing units) by flow cytometry (Fig. 1B). The functional titer of the FuG-B pseudotype was increased to 13 times as many as that of the RV-G pseudotype (ANOVA, Tukey's HSD test, p < 0.001), whereas the titer of the FuG-A pseudotype was moderately higher but not significantly different from the RV-G vector titer.
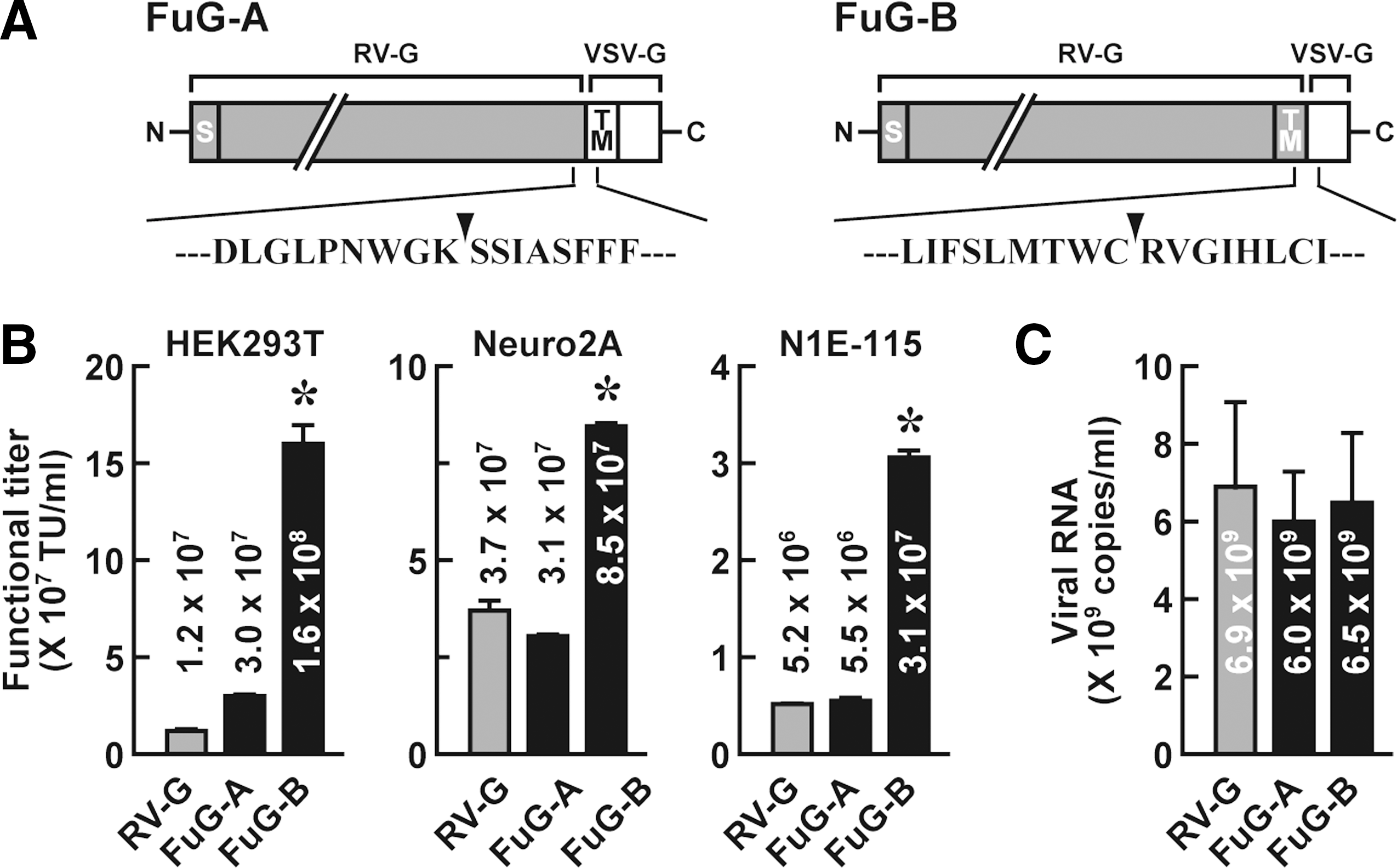
Production of HIV-1-based lentiviral vectors pseudotyped with chimeric envelope glycoproteins. (
To evaluate the functional titer in neuronal cell lines, the vector preparations were transduced into Neuro2A and N1E-115 cells (Fig. 1B). The functional titers of the FuG-B pseudotype were 2.3- and 6.0-fold higher, respectively, than that of the RV-G pseudotype (ANOVA, Tukey's HSD test, p < 0.001). The functional titer of the FuG-A pseudotype was comparable to the RV-G vector titer in these cell lines.
To estimate the concentration of physical particles of the vector, the copy number of viral RNA in vector preparations was determined by quantitative reverse transcription–PCR analysis (Fig. 1C). RNA copy number showed no significant difference among the three viral vectors (ANOVA), suggesting similarity among these pseudotypes of the production efficacy of the physical particles.
Gene transfer through retrograde transport of viral vectors pseudotyped with fusion glycoproteins
To characterize the expression pattern of the transgene, we injected the HIV-1 lentiviral vector pseudotyped with RV-G, FuG-A, or FuG-B with an equivalent RNA titer of 1.2 × 1010 copies/ml (1.0 μl × two sites) into the dorsal striatum of mice. Brain sections prepared from the injected mice were immunostained with anti-GFP antibody for histological examination. Injection of the vector pseudotypes caused transduction in a large area in the dorsal striatum, and the range and extent of transduction were similar among the three pseudotypes (Fig. 2A). To investigate transgene expression through retrograde axonal transport, we examined sections made through the primary motor cortex (M1), primary somatosensory cortex (S1), parafascicular thalamic nucleus (PF), and substantia nigra pars compacta (SNc), each of which innervates the dorsal striatum (Fig. 2B). The number of GFP-positive cells in these four brain regions was counted (Table 1). The cell numbers in the four regions of the FuG-B vector-injected mice were significantly greater than those in the corresponding regions in the RV-G vector-injected animals (ANOVA, Tukey's HSD test, p < 0.001 for M1, S1, and SNc and p < 0.01 for PF); the increases were 12-, 12-, 8.0-, and 14-fold in the M1, S1, PF, and SNc regions, respectively. Although the cell numbers in the four regions of the FuG-A vector-injected mice tended to be slightly higher than those of the RV-G vector-injected controls, there was no significant difference between the two pseudotypes. These results suggest that the FuG-B pseudotype enhances retrograde gene transfer into the brain regions innervating the dorsal striatum.
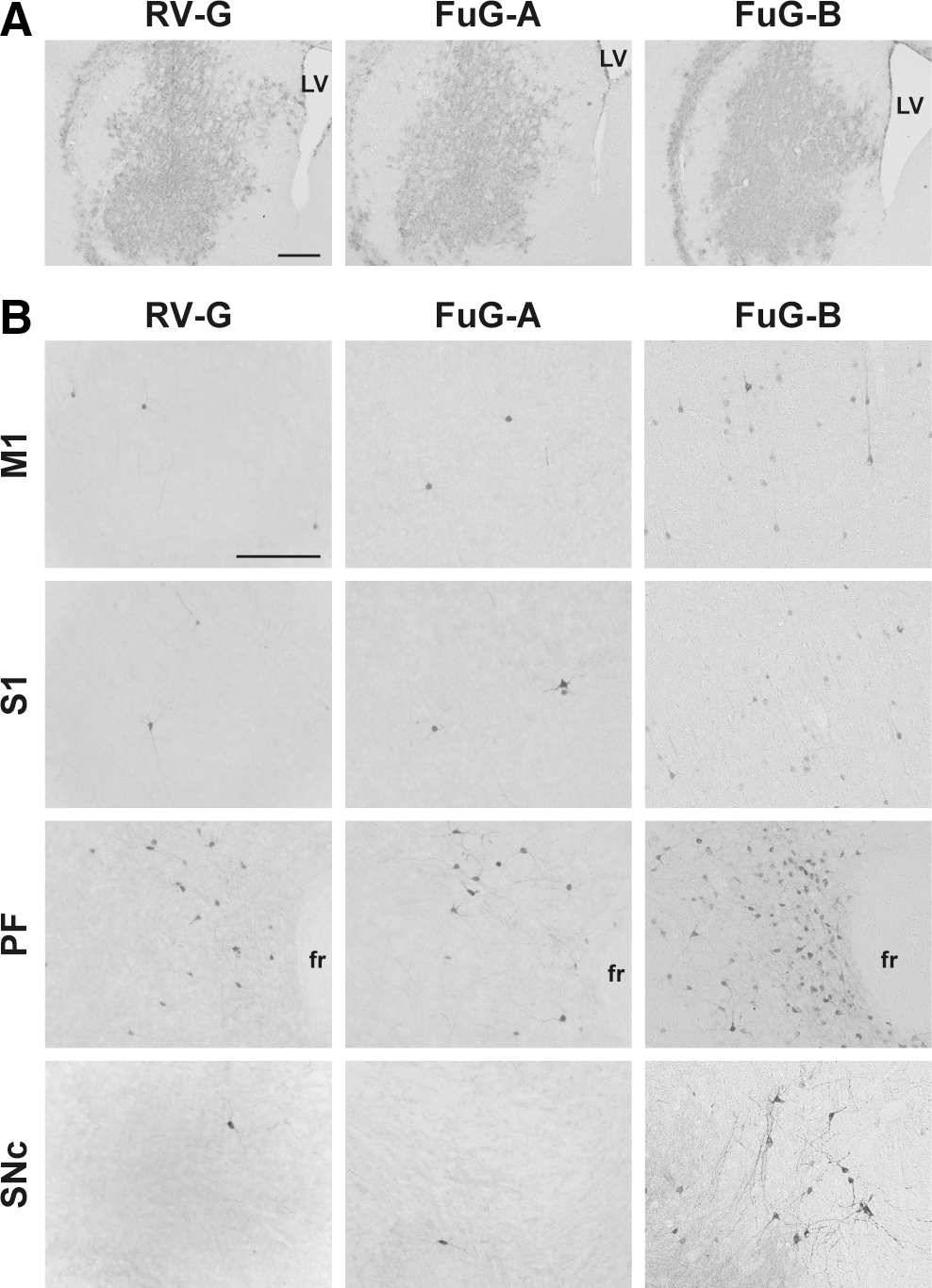
Expression patterns of transgene in brain regions after injection of pseudotyped HIV-1-based vectors into the dorsal striatum in mice. Lentiviral vectors pseudotyped with RV-G, FuG-A, or FuG-B with equivalent copy numbers of viral RNA (1.2 × 1010 copies/ml, 1.0 μl × two sites) were injected into the dorsal striatum, and 4 weeks later a series of sections were used for immunohistochemistry with anti-GFP antibody. (
Lentiviral vectors pseudotyped with RV-G, FuG-A, or FuG-B with equivalent copy numbers of viral RNA (1.2 × 1010 copies/ml, 1.0 μl × 2 sites) were injected into the dorsal striatum of mice (n = 4), and a series of sections through the M1, S1, PF, and SNc were immunostained with anti-GFP antibody. The number of GFP-positive cells per section is given.
p < 0.001, b p < 0.01, significant differences from RV-G or FuG-A pseudotype (ANOVA, Tukeys HSD test).
To further characterize the enhanced retrograde gene transfer of the FuG-B pseudotype, we injected different titers of viral vectors pseudotyped with RV-G or FuG-B (6.0 × 109–1.0 × 1011 copies/ml) into mouse striatum. Sections through the M1, S1, PF, and SNc were stained with anti-GFP antibody, and GFP-positive cells were counted (Fig. 3). Representative microscopic images of the immunohistochemistry are shown in Supplementary Fig. S1 (Supplementary Data are available online at
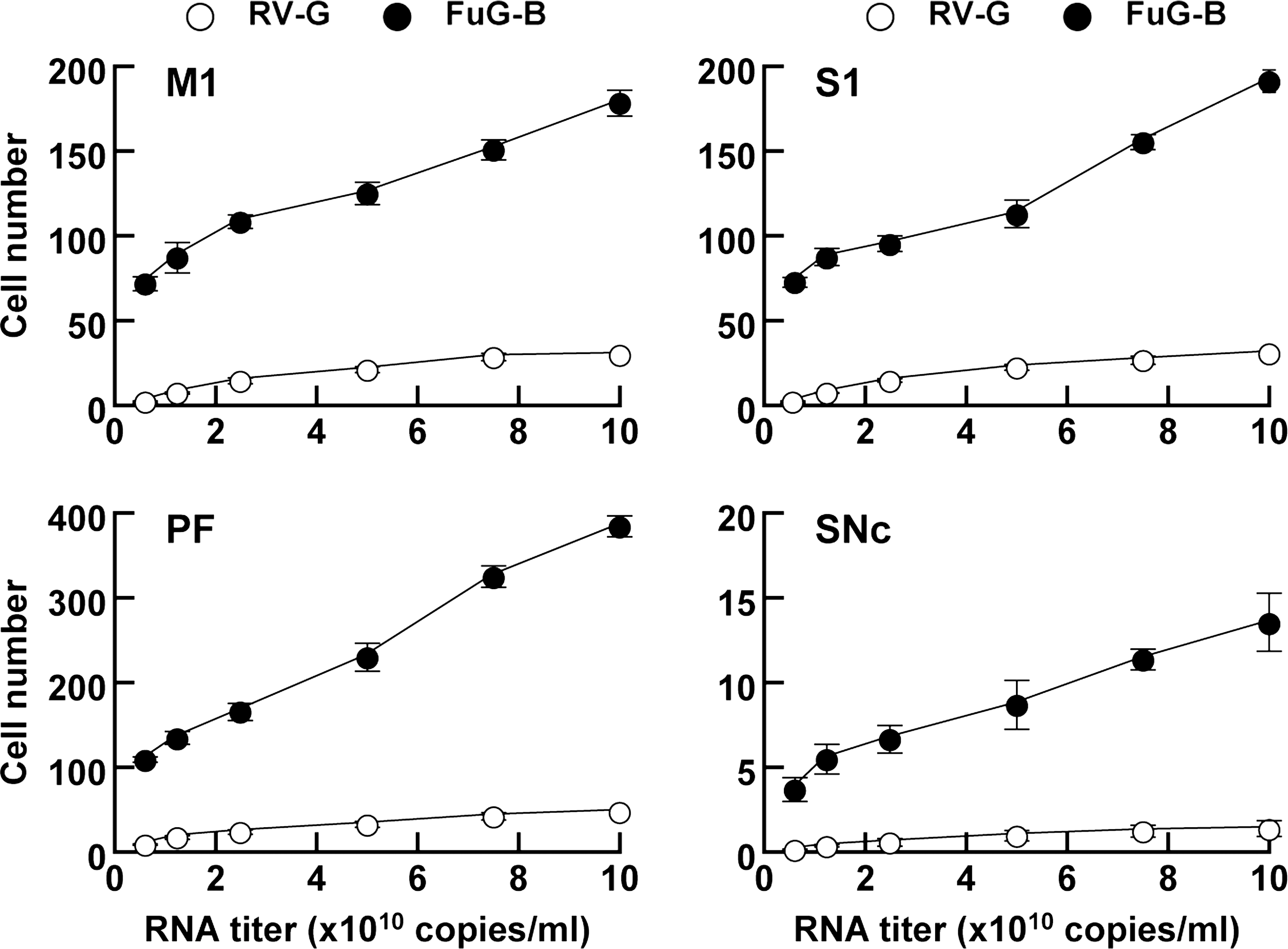
Efficiency of gene transfer through retrograde axonal transport of pseudotyped lentiviral vectors. RV-G- or FuG-B-pseudotyped vector of different RNA titers was injected into the dorsal striatum of mice (n = 4), and a series of sections through the M1, S1, PF, and SNc were used for GFP immunostaining. The number of GFP-positive cells per section is plotted. Data obtained from injection of the vectors with 1.2 × 1010 copies/ml were taken from the experiment indicated in Table 1. Representative microscopic images of the staining obtained from the vector injection are shown in Supplementary Fig. S1.
Property of gene transduction around injection sites of FuG-B pseudotype
To compare the efficiency of gene transduction around injection sites between the RV-G- and FuG-B-pseudotyped vectors, we injected different RNA titers of lentiviral vectors (7.5 × 108–1.2 × 1010 copies/ml) into the dorsal striatum and then stained striatal sections using double immunofluorescence histochemistry for GFP and the neuronal marker NeuN or for GFP and the glial marker GFAP. Representative microscopic images of the double immunohistochemistry are shown in Supplementary Fig. S2. To validate the gene transfer efficiency into neuronal and glial cells in the striatum, we normalized the numbers of GFP+/NeuN+ and GFP+/GFAP+ cells by the total numbers of NeuN+ and GFAP+ cells, respectively (Fig. 4). For both the RV-G and FuG-B vectors, the ratio of GFP+/NeuN+ relative to total NeuN+ number rose along with the increasing titer and reached a plateau at the titer of 3.1 × 109 copies/ml. Two-way ANOVA indicated no significant effect between the two kinds of vectors. Likewise, the titer-dependent change in the ratio of GFP+/GFAP+ relative to total GFAP+ number was indistinguishable between the two vectors (two-way ANOVA). This evidence indicates that the efficiency of gene transduction surrounding the injection sites was comparable between the RV-G and FuG-B pseudotypes.
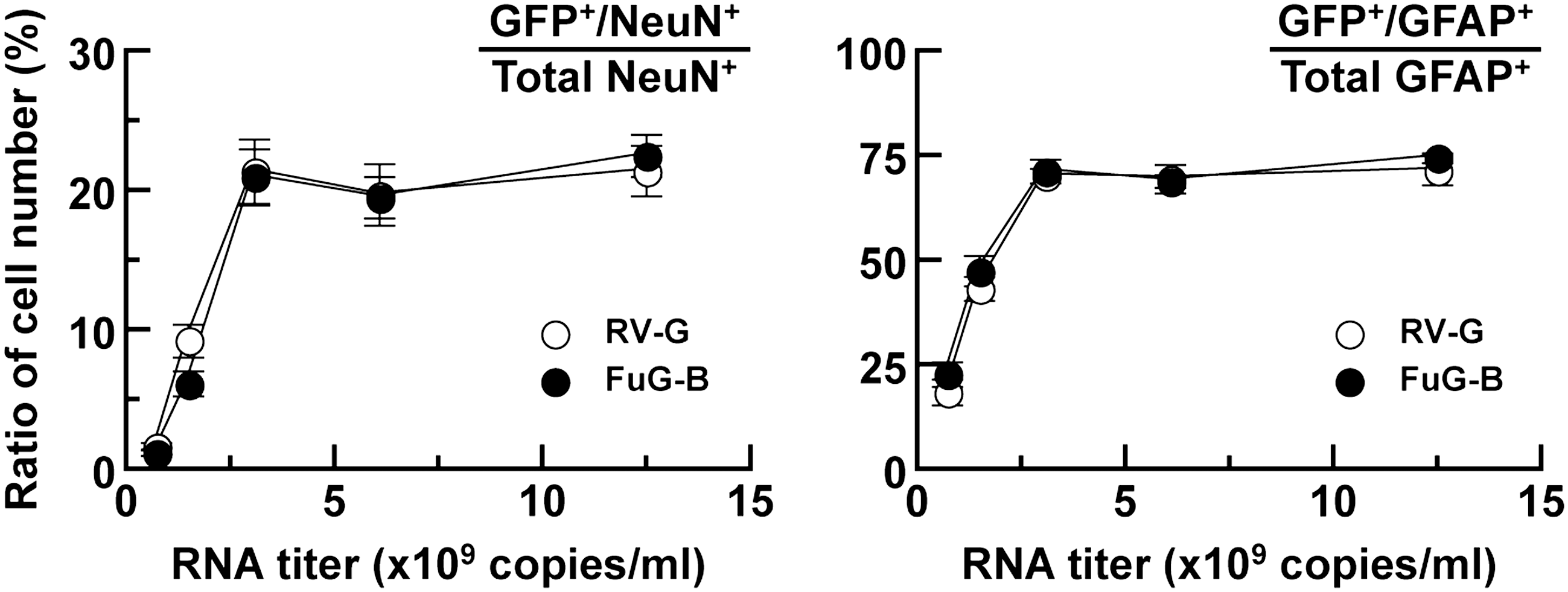
Efficiency of gene transfer around injection sites of pseudotyped vectors. RV-G- or FuG-B-pseudotyped lentiviral vector of different RNA titers was injected into the dorsal striatum of mice (n = 4), and sections through the striatum were stained by double immunofluorescence histochemistry for GFP/NeuN or for GFP/GFAP. The ratios of GFP+/NeuN+ relative to total NeuN+ cell number and GFP+/GFAP+ relative to total GFAP+ cell number are plotted. Representative microscopic images of the double immunostaining are shown in Supplementary Fig. S2.
The in vivo study demonstrated that the FuG-B-pseudotyped vector, compared with the RV-G pseudotype, does not change the efficiency of gene transduction into striatal cells in the injection areas, whereas this vector allows greater gene transfer via retrograde transport into neuronal populations. Our data suggest that the use of FuG-B for pseudotyping influences the transducing property of the lentiviral vector, thereby enhancing the retrograde transport through the nerve terminals.
Retrograde gene transfer of FuG-B pseudotype into other brain regions
To explore gene transfer via retrograde transport of the FuG-B-pseudotyped vector into other brain regions in mice, we injected the lentiviral vector (1.0 × 1011 copies/ml) into the ventral striatum (nucleus accumbens) (Fig. 5A). Injection of the FuG-B pseudotype caused GFP expression in the area including the shell and core of the nucleus accumbens around the injection site (Fig. 5B). The vector injection also led to transgene expression in many brain regions that project to the nucleus accumbens, such as the piriform cortex, subiculum, basolateral nucleus of the amygdala, and lateral hypothalamus (Fig. 5C). In addition, we injected the vector into the mPFC containing the prelimbic and infralimbic regions (Fig. 5D). The vector injection labeled the mPFC area around the injection site (Fig. 5E) and transduced retrogradely the brain regions innervating the mPFC, such as the cingulate cortex, hippocampal structure (CA1), and mediodorsal and ventromedial thalamic nuclei (Fig. 5F). These results verify that the FuG-B-pseudotyped vector enables efficient gene delivery mediated by retrograde transport into various brain regions in rodents.
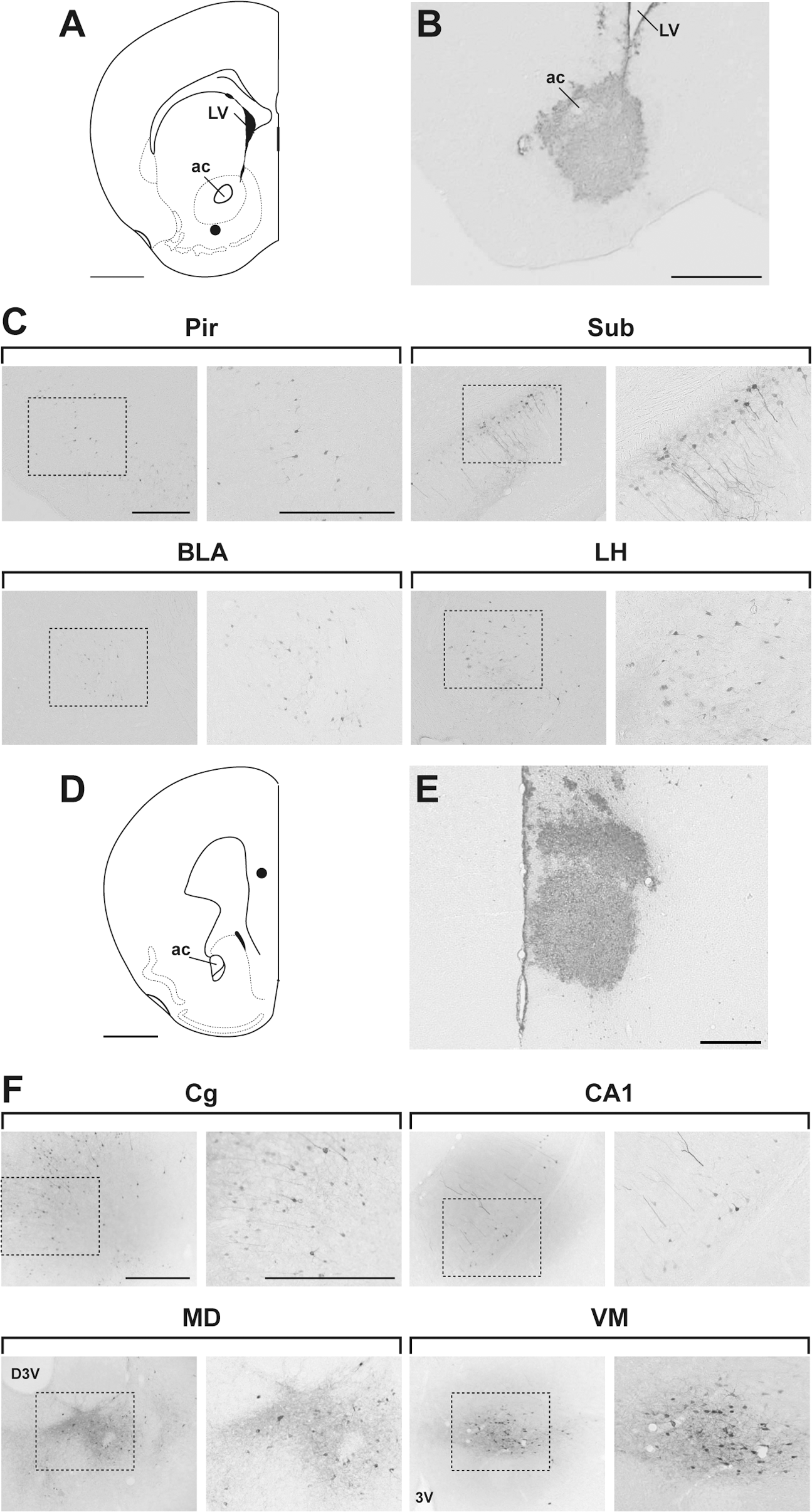
Transgene expression in brain regions after injection of the FuG-B pseudotype into the nucleus accumbens or mPFC. (
Retrograde gene delivery of FuG-B pseudotype in monkey brain
The nigrostriatal dopamine system is a major target for gene therapy of Parkinson's disease. To verify whether the lentiviral vector pseudotyped with FuG-B would permit efficient gene transfer through retrograde transport into nigrostriatal dopamine neurons in primates, we injected the vector pseudotyped with RV-G or FuG-B into the striatum (caudate nucleus and putamen) of crab-eating monkeys (8.5 × 109 copies/ml, 7.5 μl × 14 sites) and stained sections with GFP immunohistohemistry. Injection of the vector pseudotypes caused transduction in a large area in the monkey striatum, and the range and extent of transduction were similar for the two pseudotypes (Fig. 6A). To investigate the capability of retrograde transport of the vector into the nigrostriatal dopamine neurons, a series of SNc sections were analyzed. Injection of the FuG-B pseudotype transduced a larger number of neurons in the SNc compared with the RV-G pseudotype (Fig. 6B). The number of GFP-positive cells in the RV-G and FuG-B vector-injected monkeys was 64.3 and 653.6 cells per section (n = 2), respectively. SNc sections prepared from the FuG-B-injected animals were stained by double immunofluorescence histochemistry for GFP and the dopamine neuron marker TH. A majority of TH-immunoreactive neurons (74.8%, n = 2) co-expressed the GFP transgene (Fig. 6C). In addition, in the RV-G and FuG-B vector-injected animals there were GFP-positive signals like axon terminals in the substantia nigra pars reticulata, which appeared to be derived from vector transduction of striatonigral neurons (Fig. 6B). However, we found no GFP-positive signals in the cell bodies of substantia nigra pars reticulata neurons. These data indicate that the FuG-B-pseudotyped vector achieves highly efficient gene transfer into the nigrostriatal dopamine system through retrograde transport in the monkey brain. This finding suggests the potential use of the FuG-B vector for model experiments aiming at gene therapy of Parkinson's disease.
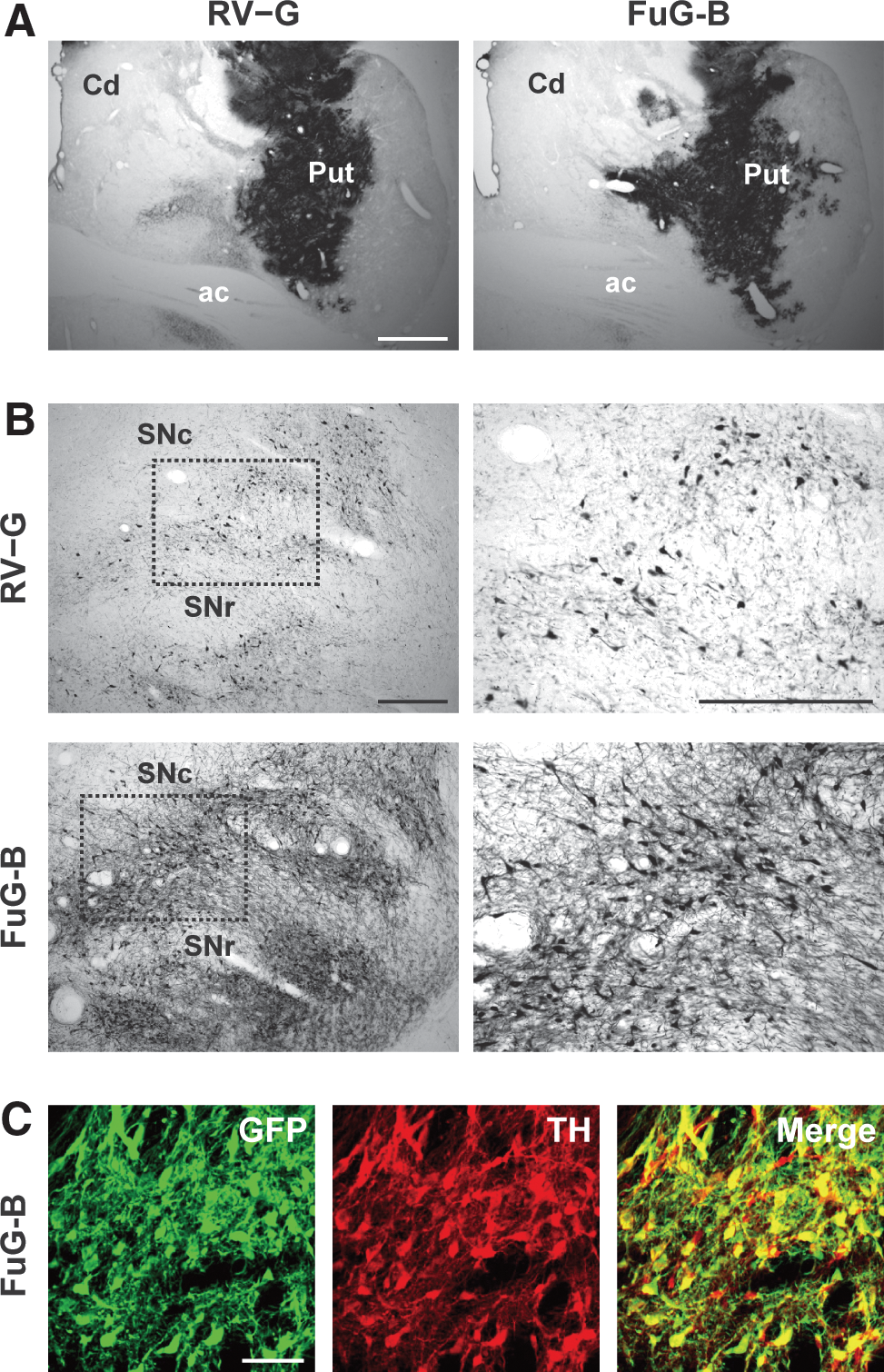
Transgene expression in nigrostriatal dopamine neurons after intrastriatal injections of pseudotyped vectors into monkey striatum. RV-G- or FuG-B-pseudotyped vector (8.5 × 109 copies/ml, 7.5 μl × 14 sites) was stereotaxically injected into the caudate (Cd) and putamen (Put), and 4 weeks later a series of sections were used for histological analyses. (
Discussion
In the present study, we developed a novel system for highly efficient gene transfer via retrograde axonal transport by pseudotyping the HIV-1-based lentiviral vector with a chimeric envelope glycoprotein. We tested two kinds of fusion glycoproteins composed of different combinations of the N-terminal and C-terminal segments derived from RV-G and VSV-G. The HIV-1 pseudotypes with these fusion glycoproteins modified the vector titer in different cell lines and in vivo neuronal gene transfer through retrograde axonal transport. The use of FuG-B, which contains the extracellular and transmembrane domains of RV-G fused to the cytoplasmic domain of VSV-G, resulted in a marked increase in vector transduction into the cell lines. The FuG-B-pseudotyped vector also enhanced the efficiency of retrograde gene transfer into the brain regions and sustained the transduction efficiency into the cells around injection sites to a similar extent as the RV-G-pseudotype. In addition, the FuG-B pseudotype achieved a high level of gene delivery into the nigrostriatal dopamine neurons through retrograde transport in the nonhuman primate brain. Thus, our results demonstrate that FuG-B pseudotyping of the HIV-1 vector greatly improves the efficiency of retrograde gene delivery into various central nervous system regions in both rodents and primates.
Substitution of the cytoplasmic domain of RV-G with the corresponding part of VSV-G in the pseudotyped HIV-1 vector was the most efficacious means to enhance the transduction of cell lines and in vivo retrograde gene transfer. The cytoplasmic domain differs in length between RV-G (44 amino acids) and VSV-G (29 amino acids), and their amino acid sequences do not show any particular homology (Rose et al., 1982). The cytoplasmic domains of viral glycoproteins are involved in specific interactions with viral nucleocapsid or matrix proteins to drive efficient viral budding (Simons and Garoff, 1980; Stephens and Compans, 1988; Cosson, 1996). Previous studies show that the VSV-G cytoplasmic domain is required for efficient incorporation of the glycoprotein into budding VSV virions (Witt et al., 1989) and that its cytoplasmic domain fused to a foreign envelope glycoprotein promotes the assembly of recombinant glycoprotein into viral particles (Owens and Rose, 1993; Johnson et al., 1997). A subsequent study using detailed mutagenesis reveals the importance of a short part of the VSV-G cytoplasmic domain, consisting of at least nine amino acids, in viral budding (Schnell et al., 1998). In contrast, the cytoplasmic domain of VSV-G, together with a part of the extracellular domain and the transmembrane domain, mediates the process of membrane fusion of the viral envelope (Robison and Whitt, 2000; Jeetendra et al., 2002). Based on our titration experiments of the pseudotyped vectors, the extent of increase in the functional titer of the FuG-B pseudotype relative to that of the RV-G pseudotype varied among the cell lines. In contrast, the viral RNA titration displayed no remarkable change in the yield of physical particles between the pseudotyped vectors. These findings suggest that replacing the cytoplasmic domain of a viral glycoprotein shifts the property of gene transduction of the pseudotyped vectors into the host cells, although the efficacy of formation or budding of the particles appears to be unaffected.
The host range of lentiviral vectors is altered by pseudotyping with different envelope glycoproteins (for a review, see Cronin et al., 2005). RV-G interacts with certain neuronal receptors, such as the nicotinic acetylcholine receptor α-subunit, low-affinity nerve growth factor receptor, and neural cell adhesion molecule (Hanham et al., 1993; Gastka et al., 1996; Thoulouze et al., 1998; Tuffereau et al., 1998). The RV-G-pseudotyped vector transduces not only neuronal cells through retrograde transport but also neuronal and astroglial cells around injection sites (Kato et al., 2007). Our FuG-B-pseudotyped vector showed enhancement in the efficiency of neuronal gene transfer via retrograde transport compared with the RV-G pseudotype, whereas its transduction efficiency into cells surrounding the injection sites resembled that of the RV-G pseudotype. These results suggest that complex mechanisms underlie the in vivo gene transfer of vectors pseudotyped with RV-G or FuG-B. These findings also suggest that substitution of the cytoplasmic domain of a viral glycoprotein may influence the mechanism involved in vector entry into synaptic terminals, which is required for enhanced retrograde transport. In addition, the FuG-B pseudotype exhibited various extents of increase of the functional titer in different cell lines compared with the RV-G vector, whereas there was no significant difference between the two pseudotypes in the in vivo gene transfer efficiency into neuronal and glial cells around injection sites. The efficiency of gene delivery of the pseudotyped vectors appears to differ between cell types or experimental conditions. Replacement of part of an envelope glycoprotein may affect incorporation of the glycoprotein into vector particles. Alternatively, this partial replacement may cause conformational changes of the glycoprotein structure, which are involved in binding to receptor molecules or membrane fusion of the vector. These changes may influence the interaction with host cells or the transduction level of the pseudotyped vector. The detailed mechanism underlying shifted transduction properties of the FuG-B vector remains to be resolved. For this purpose, extensive studies are needed as to identification of receptor molecules and characterization of biochemical processes of vector transduction of the FuG-B pseudotype.
The efficiency of retrograde gene transfer of the FuG-B-pseudotyped vector varied among brain regions. After intrastriatal injection in mice, we found marked gene transfer into the cortical areas and thalamic nucleus, but transfer into the SNc region was less efficient. After intra-accumbens injection, we observed efficient gene transfer into the piriform cortex, subiculum, amygdala, thalamus, and hypothalamus, whereas there was a low level of transfer into some cortical areas including the prelimbic and perirhinal cortices (Supplementary Fig. S3), which are known to be anatomically connected to the nucleus accumbens (Brog et al., 1993; McIntyre et al., 1996; Vertes, 2004). In addition, the efficiency of retrograde gene transfer into the SNc was much higher in the monkey than in the mouse. These variations in the transfer efficiency may reflect the distinct properties of viral transduction among neuronal types or species. The distribution of receptor molecules localized in synaptic terminals in various brain regions may influence the transfer efficiency of a pseudotyped vector. The reasons for the varied levels of retrograde gene transfer among different neuronal types or species remain to be clarified.
Retrograde axonal transport of viral vectors provides great advantages in model experiments of gene therapy for neurological and neurodegenerative diseases. Injection of adenoviral vectors into the striatum enables the delivery of genes into nigrostriatal dopamine neurons in chemically induced models of Parkinson's disease (Zheng et al., 2005; Barkats et al., 2006). Injection of these vectors into muscles permits gene delivery into the spinal cord motor neurons in an axotomy-induced injury model (Baumgartner and Shine, 1998; Perrelet et al., 2000). More recently, the intramuscular injection of the RV-G pseudotype of equine infectious anemia viral vector also results in gene transfer into motor neurons in a genetic mouse model of motor neuron disease (Azzouz et al., 2004b). Here, we successfully developed a highly efficient gene transfer system via retrograde axonal transport using the FuG-B pseudotype of the HIV-1-based vector. The FuG-B-pseudotyped vector provided efficient retrograde gene delivery into the nigrostriatal dopamine system in the nonhuman primate brain. By promoting retrograde transport of the vector, our gene transfer system will serve as a powerful tool for gene therapy trials of neurological and neurodegenerative diseases by delivering genes required for neuronal survival and protection.
Footnotes
Acknowledgments
This work was supported by a Grant-in-Aid from Core Research for Evolutional Science and Technology of Japan Science and Technology Corporation. Part of this work was supported by “Highly Creative Animal Model Development for Brain Sciences” carried out under the Strategic Research Program for Brain Sciences by the Ministry of Education, Culture, Sports and Technology of Japan. We thank St. Jude Children's Research Hospital (Dr. A. Nienhuis) and George Washington University for providing the HIV-1-based vector system. We are grateful to M. Kikuchi, N. Sato, and T. Kobayashi for technical support in animal experiments and A. Endo for conducting the immunohistochemical staining.
Author Disclosure Statement
K.K. holds a provisional patent for the application of fusion glycoproteins for the lentiviral vector system, and the other co-authors have no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.