
Abstract
Somatic in utero gene therapy aims to treat congenital diseases where pathology develops in perinatal life, thereby preventing permanent damage. The aim of this study was to determine whether delivery of self-complementary (sc) adeno-associated virus (AAV) vector in utero would provide therapeutic long-term transgene expression in a large animal model. We performed ultrasound-guided intraperitoneal injection of scAAV2/8-LP1-human Factor IX (hFIX)co (1 × 1012 vector genomes/kg) in early (n = 4) or late (n = 2) gestation fetal sheep. The highest mean hFIX levels were detected 3 weeks after injection in late gestation (2,055 and 1,687.5 ng/ml, n = 2) and 3 days after injection in early gestation (435 ng/ml, n = 1). Plasma hFIX levels then dropped as fetal liver and lamb weights increased, although low levels were detected 6 months after late gestation injection (75 and 52.5 ng/ml, n = 2). The highest vector levels were detected in the fetal liver and other peritoneal organs; no vector was present in fetal gonads. hFIX mRNA was detectable only in hepatic tissues after early and late gestation injection. Liver function tests and bile acid levels were normal up to a year postnatal; there was no evidence of liver pathology. No functional antibodies to hFIX protein or AAV vector were detectable, although lambs mounted an antibody response after injection of hFIX protein and Freund's adjuvant. In conclusion, hFIX expression is detectable up to 6 months after delivery of scAAV vector to the fetal sheep using a clinically applicable method. This is the first study to show therapeutic long-term hFIX transgene expression after in utero gene transfer in a large animal model.
Introduction
Prenatal lentivirus vector injection has given a phenotypic cure in a range of animal models (Seppen et al., 2003; Waddington et al., 2004; Williams et al., 2006), but undesirable side effects such as germline gene transfer (Lee et al., 2005) and tumor formation (Themis et al., 2005) suggest that the fetus may be sensitive to genetic pertubations with particular integrating vectors. Prenatal adenovirus vector application in large animals such as the fetal sheep produces only short-term transgene expression (Themis et al., 1999; David et al., 2003a) at low levels (Porada et al., 1998).
Adeno-associated virus (AAV) is not toxic and remains mainly in an episomal form, although integration has been associated with the development of hepatocellular carcinoma in mice (Donsante et al., 2007). When delivered prenatally to mice, AAV gives long-term transgene expression in the cochlea (Bedrosian, 2006), skin (Mühle et al., 2006), muscle (Bilbao et al., 2005), and liver (Lipshutz et al., 2001), and tolerization to human factor IX (hFIX) has been observed after fetal delivery (Sabatino et al., 2002, 2007). The only prenatal large animal study of AAV showed low-level long-term transduction of the gut and lung after intra-amniotic AAV2 delivery to non-human primates (Garrett et al., 2003).
Packaging complementary dimers of a mini-hFIX liver-specific expression cassette within a single AAV virion overcomes the need for conversion of the single-stranded genome into transcriptionally active double-stranded forms. The configuration of this self-complementary (sc) AAV vector significantly improves gene transfer (Nathwani et al., 2006).
In this report we determined the stability and level of transgene expression and biodistribution of vector following in utero gene transfer of an scAAV (scAAV-LP1-hFIXco) into a large animal. We studied this in the sheep fetus where the gestational length, physiology, and fetal and immune system development are similar to those in humans (Nathwani et al., 2006; David and Peebles, 2007).
Materials and Methods
Animals and procedures
Animal procedures were in accordance with UK Home Office regulations and the Guidance for the Operation of Animals (Scientific Procedures) Act (1986). Time-mated pregnant Romney Marsh ewes were generated, and gestational age was confirmed as described (David et al., 2006). General anesthesia was induced with thiopentone (1 g intravenously), and after intubation, the ewes were then ventilated on 2% halothane in oxygen. The ewes recovered after extubation.
The scAAV vector (1.5 × 1011–9 × 1012 vector genomes/kg) was administered into the peritoneal cavity in early (n = 4, 60–65 days) or late (n = 2, 97 and 105 days of gestation) (term = 145 days) gestation under ultrasound guidance (Table 1). A 22-gauge spinal needle (Cook, Letchworth, UK) was inserted through the maternal skin and the uterus into the amniotic cavity, avoiding passage through the placentomes if possible. The needle was then passed through the anterior abdominal wall just below the level of the cord insertion to avoid the fetal liver. Vector was injected slowly over a 1-min interval under view. All injection procedures were recorded and reviewed for correct needle and vector placement. Intraperitoneal delivery of the vector was confirmed by observing microbubbles moving throughout the peritoneal cavity during vector injection. The needle was flushed through with 40 ml of saline to clear the dead space.
Fetal plasma was sampled either by ultrasound guidance from the intrahepatic umbilical vein under anesthesia or at postmortem examination. Vector dose was determined using either the fetal weight at postmortem examination when this was within 3 days of vector injection or according to fetal weight charts derived from previous fetal gene therapy experiments (David et al., 2003a; Peebles et al., 2004; Weisz et al., 2005). GA, gestational age; PI, post-vector injection; PN, postnatal; vg, vector genomes.
Following recovery, fetal well-being and growth were monitored daily for 2 days using serial ultrasound measurement of fetal biometry (Kelly et al., 1987) and umbilical artery Doppler examination and then weekly until scheduled postmortem examination or birth.
With the mother under general anesthesia, fetal blood was sampled from the intrahepatic umbilical vein under ultrasound guidance at intervals following vector injection (n = 4) (Themis et al., 1999). Birth of the lambs and sampling of lamb blood were as described (David et al., 2003a).
Vector
We used scAAV vector pseudotyped with serotype 8 capsid proteins (scAAV2/8-LP1-hFIXco). Vector was produced as previously described (Nathwani et al., 2006) and titrated by slot blot (Nathwani et al., 2001). The dose of vector per kg of sheep fetus was determined using historical fetal sheep data (David et al., 2003a,b, 2006; Peebles et al., 2004).
Analysis
Postmortem examination performed during fetal life (n = 2) or after birth (n = 4) (Table 1) involved collection of maternal and fetal tissues, with tissue processing for histological examination and DNA extraction (David et al., 2003a). Enzyme-linked immunosorbent assay was used to measure plasma hFIX levels (Nathwani et al., 2002) and antibodies to hFIX and AAV (Nathwani et al., 2002) in duplicate. Functional assays were used to detect hFIX inhibitor antibodies (Bethesda assay limit of detection, 2 Bethesda units) (Nathwani et al., 2001) and anti-AAV8 antibodies (Nathwani et al., 2002). Liver function tests and bile acids were analyzed according to standard protocols (Royal Veterinary College, Hawkshead, UK) and compared with control sheep results. Interleukin-6 was measured in fetal blood using a kit following the instructions of the manufacturer (R&D Systems, Abingdon, UK). Fetal and maternal tissue spread of vector genomes was detected by hFIX polymerase chain reaction (PCR) (David et al., 2003a). Primers designed to amplify the endogenous sheep sequence associated with the TATA box binding protein were used as a positive control (David et al., 2008). The control liver was from a gestation-matched uninjected sheep fetus. Genome copy number was measured using quantitative PCR (Nathwani et al., 2007) to the codon-optimized FIX sequence (forward primer, GGG CAA GTA TGG CAT CTA CA; reverse primer, AAA GCA TCG AGT CAG GTC AG). DNA (20 ng) was used in the PCR with SYBR Green master mix (Applied Biosystems, Warrington, UK) and detected on the Realplex Mastercycler (Eppendorf AG, Hamburg, Germany). Factor IX levels were determined from a standard curve prepared with linearized plasmid DNA spiked into naive fetal sheep liver DNA. The copy number per diploid cell was based on 6.6 pg of DNA per cell (Gregory, 2000). To normalize DNA content, the housekeeping gene TATA box binding protein was measured for samples against a standard curve of different DNA concentrations from 2.5 to 160 ng per reaction.
RNA preparation and quantitative RT-PCR
Tissues were homogenized in 1 ml of TRIzol® (Invitrogen, Carlsbad, CA) by passing through a 21-gauge needle (Fisher Scientific UK, Loughborough, UK). RNA was extracted from the tissues as per the manufacturer's protocol and redissolved in diethylpyrocarbonate-treated sterile water by incubation at 55°C for 10 min. The concentration of RNA was confirmed by spectroscopy. Two micrograms of RNA from each tissue was treated with DNaseI (Roche Applied Science, Mannheim, Germany) in a 20-μl reaction at 37°C for 1 hr in order to remove contaminating genomic DNA. RNA was then purified with two rounds of phenol–chloroform extraction, precipitated using isopropanol, and resuspended in diethylpyrocarbonate-treated water as before. The purity and concentration of the RNA were again assessed using spectroscopy. The entire volume of RNA was then used for reverse transcription, performed using the RETROscript kit (Applied Biosystems/Ambion, Austin, TX) following the manufacturer's instructions. Quantitative PCR was performed on the cDNA samples thus generated using primers specific to the codon-optimized hFIX sequence (primer sequences given above). Primers to the constitutively expressed gene encoding sheep TATA box binding protein were used as a control (David et al., 2008). PCRs were run on a Realplex Mastercycler® (Eppendorf) using cycling conditions of 95°C for 2 min, followed by 40 cycles of 95°C for 15 sec, 55°C for 15 sec, and 68°C for 20 sec. The fold differences in hFIX RNA levels in different tissues were calculated based on the 2−ΔΔCT values.
Immune tolerance
Lambs were tested for immune tolerance to hFIX protein by antigen challenge together with Freund's adjuvant. Lambs received a subcutaneous injection of 1 ml of aqueous solution of hFIX protein (1 mg, Replenine-VF 1000, Bio Products Laboratory, Elstree, UK) mixed with 50% Freund's complete adjuvant in a 1-ml volume (Sigma-Aldrich Chemie GmbH, Steinheim, Germany) injected subcutaneously at four sites. Lamb serum samples were assayed for hFIX antibodies before and at time intervals after injection.
Results
Effect of gestational and neonatal age on hFIX expression and AAV transgene copy number
The hFIX transgenic protein was used as a secreted marker of transduction that can be assessed by repeated blood sampling in utero and in postnatal life. Therapeutic levels of hFIX were observed in the neonatal period following late gestation intraperitoneal injection of scAAV2/8-LP1-hFIXco into two fetal sheep, with the highest expression being observed 2–3 weeks after transduction (2,055 ng/ml in a male lamb and 1,687.5 ng/ml in a female lamb) (Fig. 1A and Table 1). Thereafter, there was a falloff in hFIX expression, which correlated with increasing body weight of the transduced lambs (Fig. 1B and Table 1), to steady-state levels of 75 and 52.5 ng/ml hFIX levels, respectively, which were maintained until postmortem examination at 6 months of age. Vector genomes were detectable in the lambs' livers at 6 months of age at 2.49 and 0.097 copies per diploid cell, respectively.
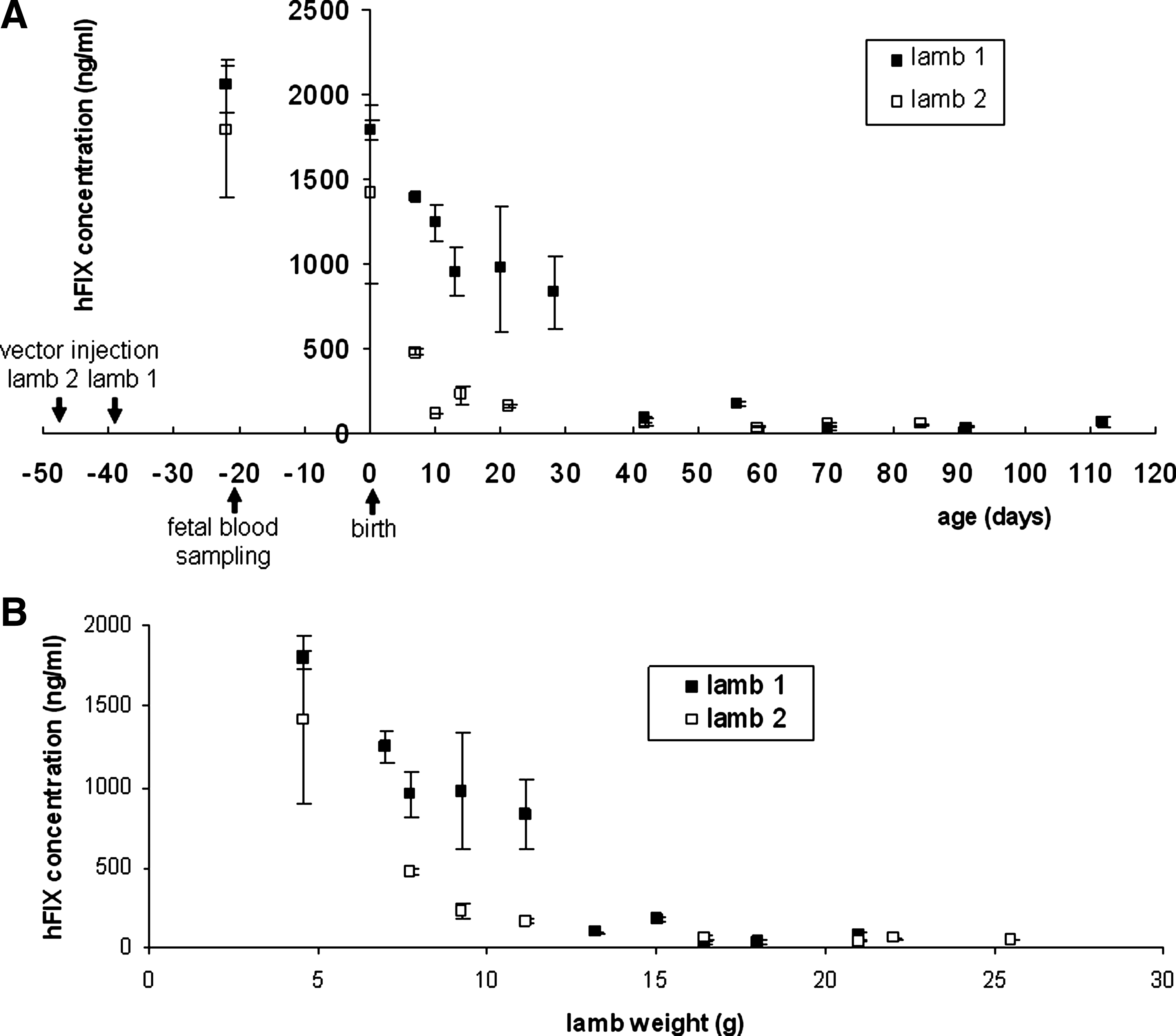
Plasma hFIX levels in lambs born after late gestation intraperitoneal fetal scAAV2/8-LP1-hFIXco injection fall as neonatal age and weight increase. (
In contrast, after early gestation intraperitoneal vector injection, the highest hFIX protein level (mean, 435 ng/ml; n = 1 male lamb) was detected 3 days after fetal injection but measured 85 ng/ml 3 weeks after vector injection in another animal (n = 1 female lamb) (Fig. 2 and Table 1). This reduction in hFIX protein expression mirrored the rapid increase in fetal liver size in early compared with late gestation. Transgenic hFIX protein was still detectable (mean, 37 and 39.5 ng/ml) in the plasma of two male lambs at birth, over 12 weeks after vector injection, but by 1 month after birth transgenic hFIX protein was no longer detectable (Table 1). There were 4.6 and 0.054 copies per diploid cell scAAV transgene detected in the fetal liver at 3 and 21 days, respectively, after scAAV2/8-LP1-hFIXco vector injection. By 1 year after birth, scAAV transgenes in the liver of early gestation-injected lambs were below the level of detection of the assay. There was no plasma hFIX detectable in any ewes at any time after scAAV2/8.LP1-hFIXco injection.
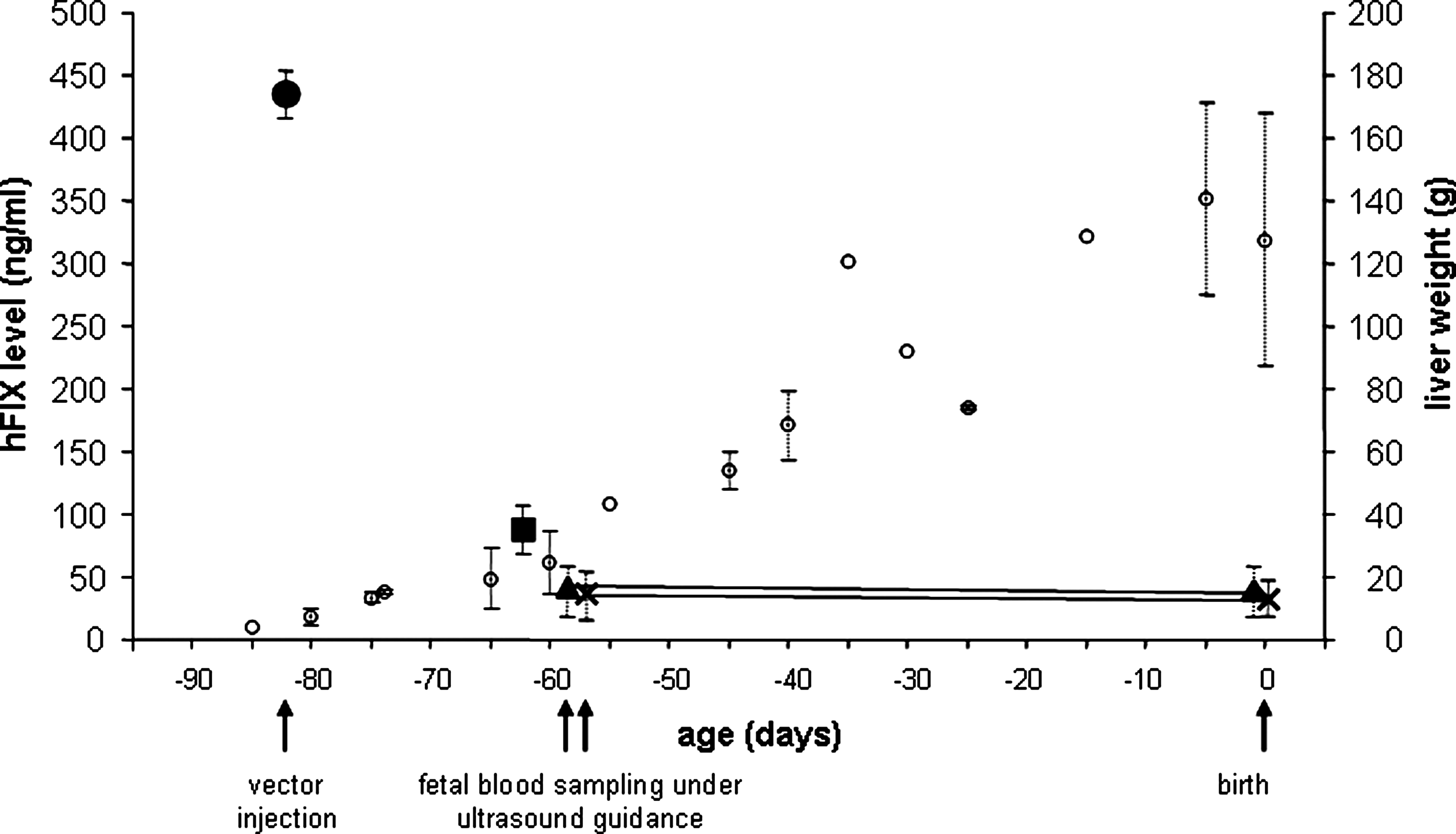
Plasma hFIX level after early gestation intraperitoneal injection of scAAV2/8-LP1-hFIXco is correlated with fetal liver weight. hFIX was measured in the plasma of fetal sheep or lambs taken by ultrasound-guided intrahepatic umbilical vein sampling or at postmortem examination. Lambs 5 and 6 were sampled in utero and at birth. Blood samples were tested twice in duplicate for hFIX, and results are shown as mean ± SEM values (black circle lamb 3; black square, lamb 4; black cross, lamb 5; black triangle, lamb 6). Fetal or lamb liver weights were used from historical controls and are shown as mean ± SEM values (open circles) (David et al., 2003a; Peebles et al., 2004; Weisz et al., 2005).
Outcome
Vector delivery to the fetal sheep by ultrasound-guided intraperitoneal injection was well tolerated, and in particular there were no fetal or maternal deaths. In utero gene transfer of scAAV was not associated with perturbation of inflammatory markers such as interleukin-6 (data not shown). In addition, hematological and biochemical parameters, including liver function and bile acids, in the fetal and lamb blood were normal.
Postmortem examination of fetuses and lambs up to a year after birth showed no peritoneal adhesions, but there were patches of neutrophilic infiltration in the liver of 6-month-old lambs injected in late gestation. A year after early gestation injection, one lamb had hypercellular glomeruli with iron deposition within tubular renal epithelial cells and focal tubular calcification. The other early gestation-injected lamb was normal. There were no histological abnormalities in any other fetal or maternal tissues.
Immune response
Neutralizing antibodies to hFIX were not detected in the plasma of fetal sheep or lambs injected at any stage of gestation, although neutralizing antibodies to the AAV8 capsid protein were detectable after late but not after early gestation injection. Subcutaneous injection of hFIX protein combined with Freund's complete adjuvant gave increasing levels of hFIX antibodies in the plasma of all lambs (n = 4, Fig. 3), showing that immune tolerance if present could be broken.
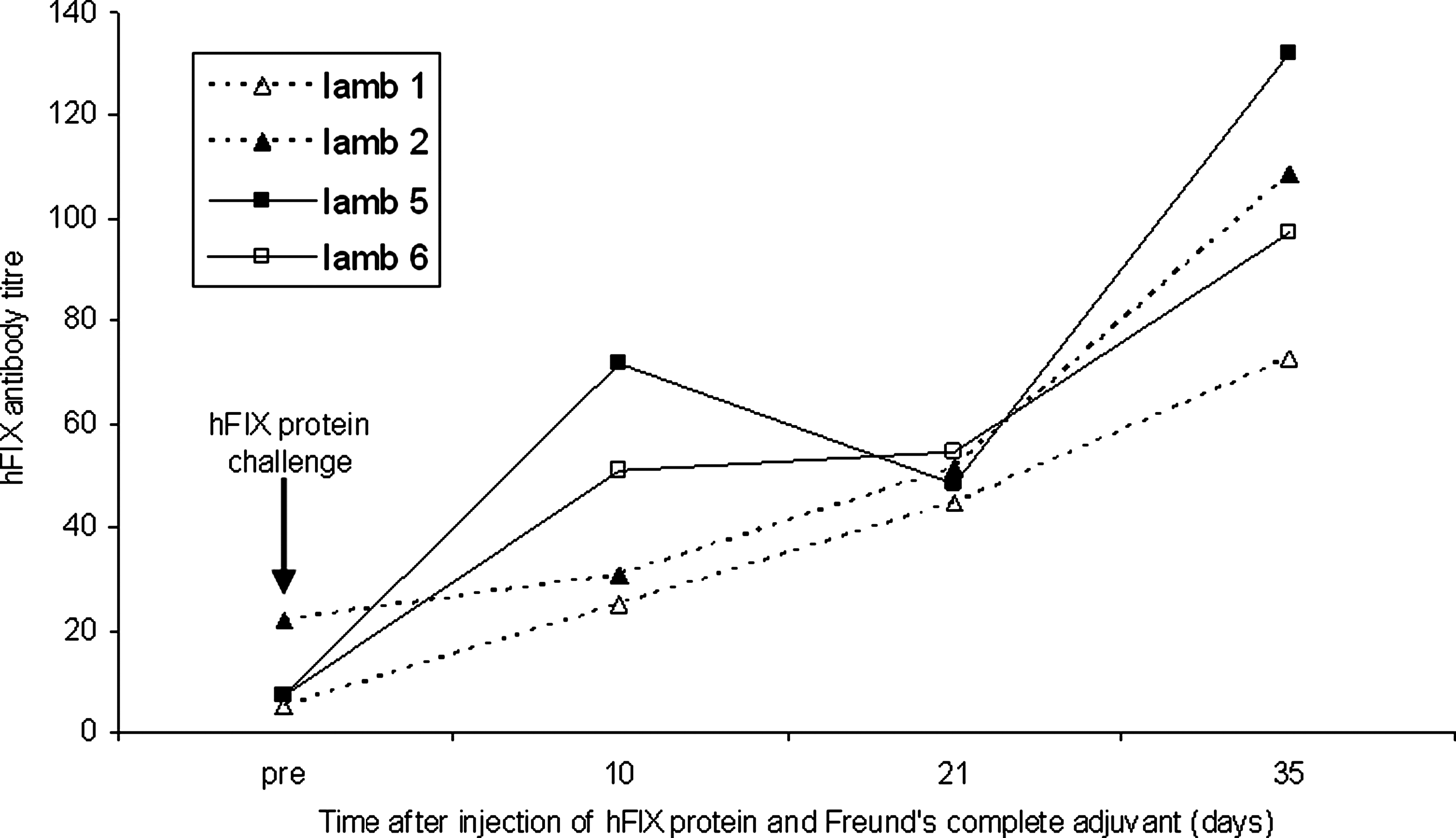
Fetally injected lambs are not tolerant to exogenous hFIX protein in postnatal life after early or late gestation intraperitoneal injection of scAAV8-hFIX. Lambs that had received fetal scAAV vector mounted an immune response to hFIX protein (1 mg) when given subcutaneously with Freund's adjuvant. On the day of hFIX challenge, lamb serum was collected, and baseline hFIX antibody levels (pre-injection) were measured, after which the lamb received a subcutaneous injection of hFIX protein with Freund's adjuvant as indicated (arrow).
Vector biodistribution and expression
Vector biodistribution studies using a sensitive semiquantitative PCR assay showed the presence of the vector genome in all tissues examined, including the brain (Fig. 4). Real-time PCR was then used to calculate genome number. Several organs (liver, spleen, adrenal, kidney, and bladder) had a relatively high copy number (4.4, 0.95, 0.27, 1.04, and 15.3 copies per diploid cell, respectively) that was comparable to levels observed in the liver. Control fetal sheep liver had 0.013 copies per cell.
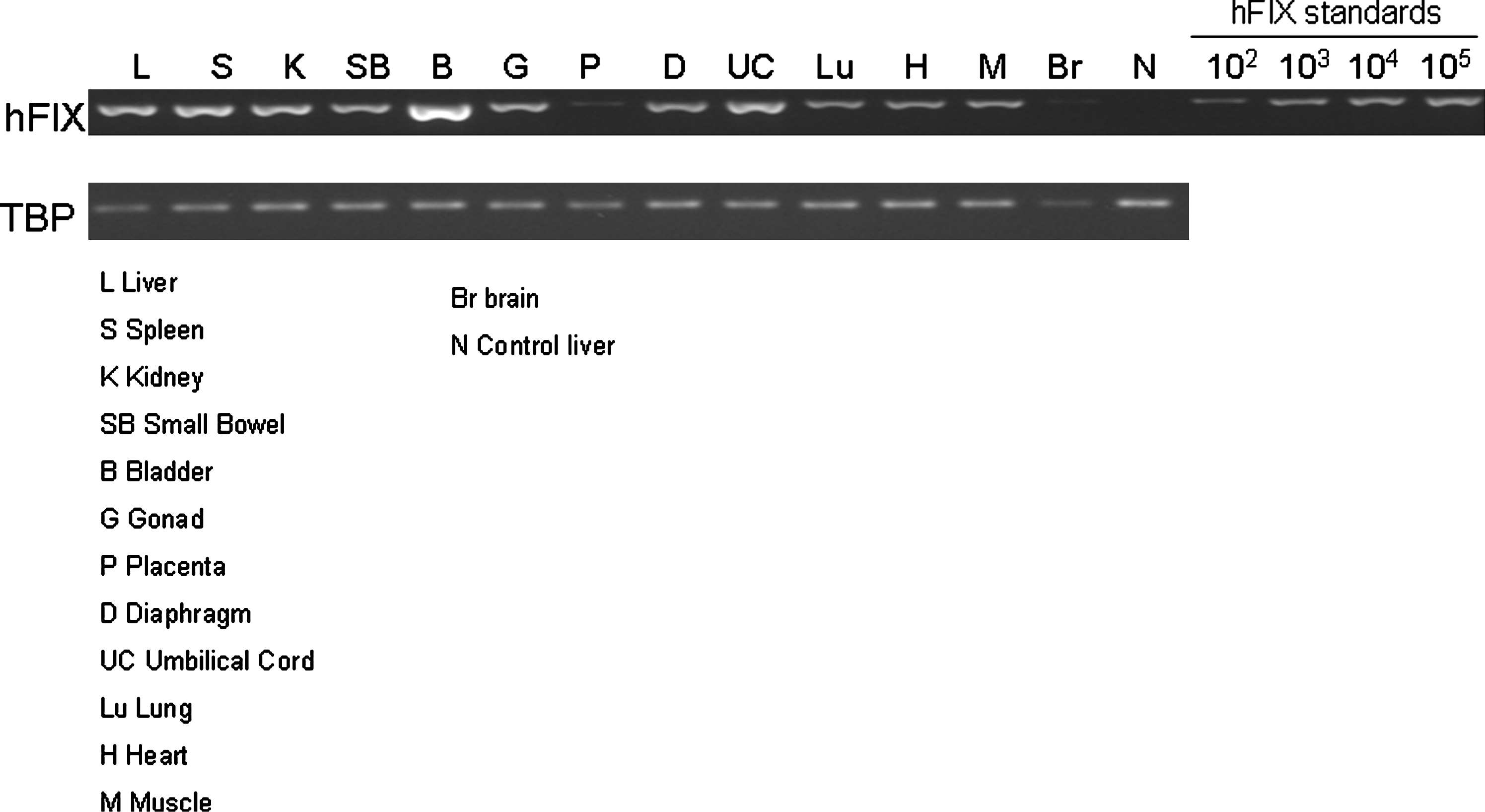
Systemic spread of the hFIX transgene in fetal sheep tissues 3 days after early gestation ultrasound-guided intraperitoneal injection of scAAV8-hFIX. Primers to hFIX and TATA box binding protein (TBP) were amplified from fetal tissue DNA as described (David et al., 2003a). hFIX standards were prepared on a naive fetal sheep liver background.
hFIX mRNA was measured by quantitative RT-PCR in tissues (liver, spleen, stomach, kidneys, adrenal, bladder, diaphragm, lungs, bone marrow, placenta) from fetal sheep injected in early gestation (n = 2) and from lambs injected in early (n = 2) and late (n = 2) gestation (Fig. 5). hFIX mRNA levels were high in the liver samples but were undetectable in the non-hepatic tissues, including those with a relatively high LP1-hFIXco transgene copy number.
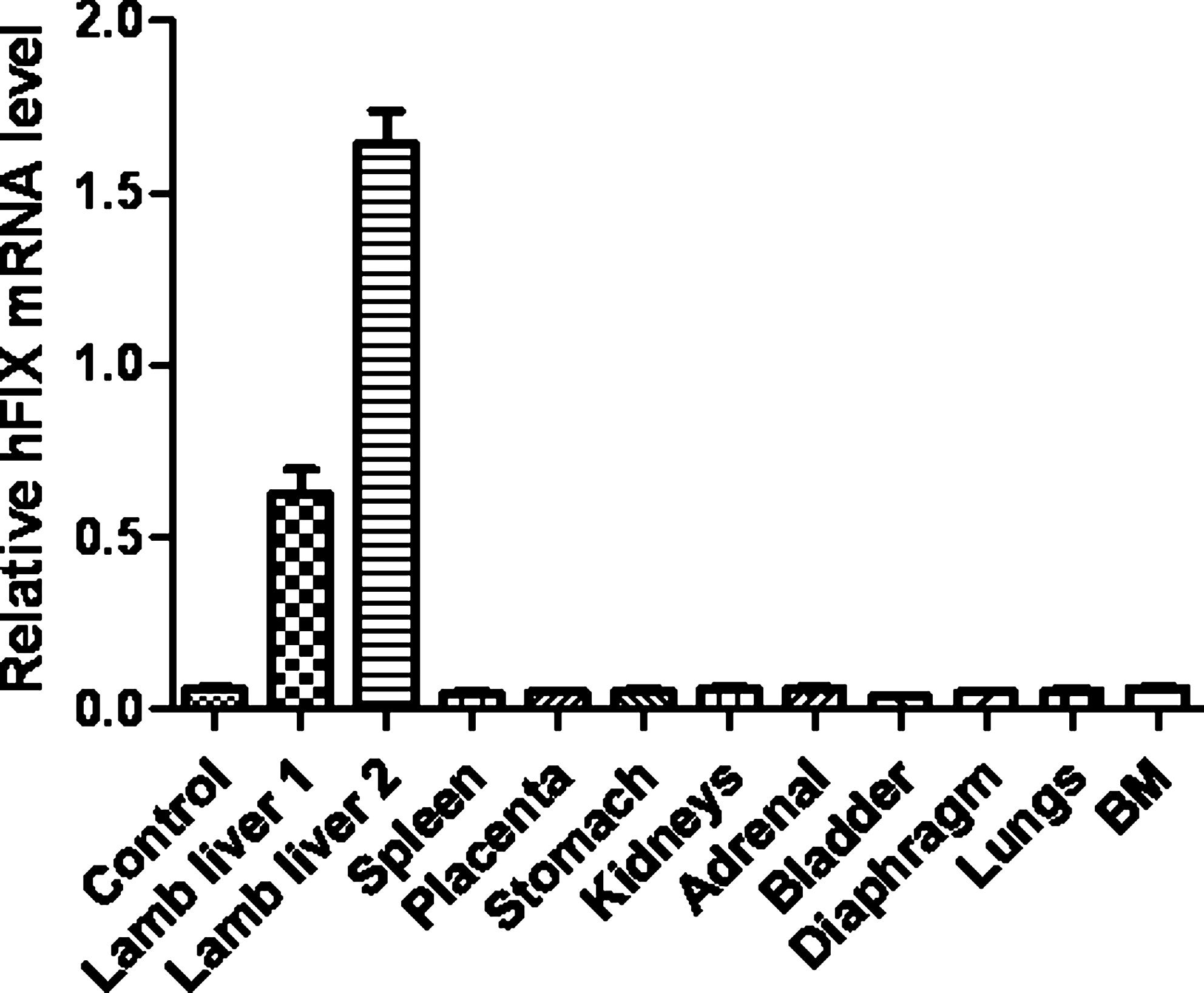
hFIX mRNA measured by quantitative RT-PCR in hepatic and other non-hepatic tissues in lambs after ultrasound-guided intraperitoneal injection of scAAV8-hFIX in late gestation. hFIX mRNA from lamb tissues was measured using quantitative RT-PCR and normalized to TATA box binding protein mRNA levels. hFIX mRNA was only detectable in lamb liver samples. There is no detectable hFIX mRNA in non-hepatic lamb tissues or in the liver from an uninjected lamb of comparable age (control). BM, bone marrow.
Discussion
This is the first study to show perinatal therapeutic transgenic protein expression in a large animal fetus. We used a clinically applicable injection technique (Gallot et al., 2004) to achieve gene transfer that was well tolerated, and in particular there were no fetal or maternal deaths. Transgenic protein expression declined after birth, closely following the increase in lamb weight and liver growth, and apparently stabilized at a low level (∼50 ng/ml or 1% of normal hFIX levels). The exponential fall in hFIX expression and scAAV transgene copy number following in utero gene transfer suggests that the scAAV integration frequency is low, which is an important safety implication.
During fetal and neonatal life the liver and blood volume increase rapidly, while cell division is at its most intense, and all of these factors may be responsible for the fall in hFIX level we observed in fetal and lamb plasma. When assessed approximately 20 days after vector delivery, plasma hFIX levels were more than 10 times lower after early gestation compared with late gestation vector delivery, in spite of the more than 10-fold higher vector dose/kg applied in early gestation. The more rapid liver growth and increase in blood volume in early gestation could be the explanation. Alternatively, the relative immaturity of the fetal liver at the early gestation time point might have been responsible, and this possibility warrants further study. A rapid decline in AAV expression to stable low levels has been reported in association with hepatocellular proliferation after AAV2/8 vector-mediated gene transfer to neonatal ornithine transcarbamylase-deficient Spf ash mice compared with adult animals (Cunningham et al., 2009). In a similar study, clusters of ornithine transcarbamylase-expressing cells in adult mice treated with AAV2/8 as neonates were observed, which the authors concluded were as a result of vector integration (Cunningham et al., 2008). This loss of expression in the developing liver of the fetus and neonate has important consequences for AAV-mediated gene therapy in humans. The rapid growth of the fetal sheep liver is similar to that of the human. Our data suggest that sequential transduction may be required in utero and postnatally to maintain expression within the therapeutic range. Although this has clear logistical implications, the relatively nontoxic features of AAV and the availability of an array of different serotypes with distinct immunological properties make this feasible.
The immune response we observed to the AAV8 capsid after late but not early gestation vector injection is consistent with classic experiments on the development of the fetal immune response in the sheep (Silverstein et al., 1963; Fahey and Morris, 1978), which demonstrated that the fetal response to antigenic stimulus varies according to the type of antigen and with gestational age. In previous experiments using adenovirus gene transfer to fetal sheep we observed an immune response to adenovirus at any gestational age, which is consistent with the higher immunogenicity of this vector compared with AAV. Antibodies were also observed to hFIX protein after late but not early gestation injection (Themis et al., 1999; David et al., 2003a). This could be related to persistence of transgenic protein expression, which was much longer after scAAV compared with adenovirus gene transfer, where hFIX levels declined rapidly within 2 weeks of vector injection (Themis et al., 1999). Nevertheless, the low-level long-term hFIX expression after scAAV administration in utero was not sufficient to maintain immune tolerance. Studies on AAV hFIX or human Factor VIII administration in neonatal mice suggest that the magnitude and persistence of antigen determine the humoral immune responses (Zhang et al., 2007) and compared with low-expression, high-transgenic protein expression avoided antibody formation and reduced the immune response to protein challenge (Xu et al., 2007). Repeating in utero administration or using a higher initial dose of scAAV may maintain sufficiently high transgenic protein expression levels to achieve tolerance.
We observed systemic spread of scAAV in fetal tissues, in keeping with our findings after intraperitoneal delivery of adenovirus into fetal sheep (David et al., 2003a). This contrasts with the relatively liver-restricted biodistribution of scAAV we previously observed in adult mice and non-human primates (Nathwani et al., 2006). This wide biodistribution of scAAV may represent an advantage when targeting disorders such as lysosomal storage disease that affect multiple organs. In contrast, hFIX mRNA was only detectable in hepatic tissues. There was no detectable hFIX mRNA in non-hepatic tissues, including those with a relatively high LP1-hFIXco transgene copy number. This is consistent with our previous experience in adult mice and non-human primates (Nathwani et al., 2006) and confirms that the LP1 promoter mediates liver-specific transgenic protein expression in the fetal sheep.
There were minor histological abnormalities in the liver of two lambs, but there was no evidence of liver function disturbance. In a gene therapy trial using AAV2 hFIX in human adults with severe hemophilia B, a transaminitis was observed in the blood of two participants (Manno et al., 2006). Analysis of T-cell responses to the AAV2 capsid suggested that T-cell-mediated destruction of AAV2-transduced hepatocytes may have contributed to the failure of persistent gene expression that was observed. AAV serotype 8 may have advantages for human gene therapy because of the low prevalence of neutralizing antibodies to its capsid (5% compared with 70% for AAV2) and because it is less likely to evoke a cell-mediated immune response.
The number of animals per cohort is small in our study as these long-term large animal experiments are complex and expensive. Further experiments using scAAV in the fetal macaque are underway to study the biology of transgenic protein expression and the safety profile. Strategies such as repeated in utero vector delivery or postnatal re-injection with other AAV serotypes would be needed at present to achieve long-term therapeutic transgenic protein levels in the growing neonate. The levels of transgene expression achieved, however, would be therapeutically useful during the perinatal period in patients with bleeding disorders such as Factor VII deficiency and might be sufficient in the early neonatal period to prevent pathological damage in other congenital diseases such as metabolic storage disorders.
Footnotes
Acknowledgments
This work was supported by the British Medical Research Council (grant G9901081). S.W. is the recipient of a fellowship from the Katharine Dormandy Haemophilia Trust. This work was undertaken at University College London Hospital/University College London, which received a proportion of funding from the Department of Health's NIHR Biomedical Research Centres funding scheme.
Author Disclosure Statement
No competing financial interests exist for any author.