
Abstract
In the spinal cord, protein kinase C isoform γ (PKCγ) plays an important role in the development of central pain sensitization. However, there are currently no specific PKCγ inhibitors available. Therefore, the aim of the present study was to assess the role of PKCγ in the modulation of pain using a more selective experimental tool. Although small interfering RNAs have been used to silence genes in neurons, in vivo delivery of RNA interference (RNAi) remains a major challenge, thus limiting its applications. Here we developed a highly efficient method of lentiviral-mediated delivery of short-hairpin RNAs targeting PKCγ for in vivo gene silencing in the spinal cord of rats. This method decreased the expression of PKCγ mRNA and protein, and additionally attenuated chronic constriction injury–induced mechanical allodynia and thermal hyperalgesia for more than 6 weeks. Our study suggests that PKCγ is a potential RNAi target for neuropathic pain. Furthermore, the lentiviral vector delivery strategy holds great promise as a novel approach for the treatment of neuropathic pain and study of PKCγ gene function.
Introduction
Protein kinase C isoform γ (PKCγ) belongs to the protein kinase C (PKC) superfamily and is found mostly in the brain and spinal cord. It is thought to be an important second messenger in the intracellular signal transduction of various cellular processes (Li et al., 2001). Accumulating evidence indicates that the activation of PKCγ in the spinal cord is involved in spinal hyperexcitability and/or persistent pain states (Malmberg et al., 1997; Martin et al., 2001; Celerier et al., 2004), suggesting that PKCγ plays an important role in the development of central sensitization, especially in neuropathic pain after nerve injury. From a clinical perspective, the very restricted spinal cord location of the PKCγ-containing interneurons is advantageous (Malmberg et al., 1997). If selective inhibitors of PKCγ can be developed, it may be possible to alleviate nerve injury–induced neuropathic pain states without the profound side effects that are inevitable with nonselective inhibitors of PKC. However, there are no specific PKCγ inhibitors available currently. The aim of the present study was to investigate the roles of PKCγ in pain modulation by using small interfering RNAs (siRNAs) to silence PKCγ gene expression selectively.
In recent years, RNA interference (RNAi) has emerged as a powerful tool to induce loss-of-function phenotypes by posttranscriptional silencing of gene expression (Fire et al., 1998; Dorn et al., 2004). Compared with gene knockout techniques, RNAi-based gene silencing is more rapid and cost-effective (Nemunaitis et al., 2010). Several methods for delivery of DOR, NR2B, TrkB, TRPV1, and NR1 as molecular targets have been successfully established using RNAi-mediated gene silencing (Luo et al., 2005; Christoph et al., 2006; Garraway et al., 2009; Suzuki et al., 2009). These various delivery methods for the expression of short hairpin RNA (shRNA) include direct application of naked siRNAs and the use of lipid-based delivery vehicles (Luo et al., 2005). However, these methods are limited due to low transduction efficiencies, weak control of gene expression, and the short duration of the effects, especially when it is necessary to generate long-term gene silencing in vivo (Hassani et al., 2005).
Viral delivery of shRNA expression cassettes allows efficient transduction in tissues such as the central nervous system in vivo. Recently, long-term knockdown of gene expression has been achieved using lentiviral vector constructs that express hairpin RNAs within vector-infected cells, including nondividing cells (Naldini et al., 1996; Rubinson et al., 2003; Xia et al., 2004; Manjunath et al., 2009). A lentiviral delivery of an shRNA expression system has been developed as a new therapeutic strategy for the treatment of amyotrophic lateral sclerosis (Raoul et al., 2005). This system provided useful tools for elucidating gene function by analysis of loss-of-function phenotypes and for exploring the application of RNAi in gene therapy (Matta et al., 2003). Targeted delivery of shRNA against PKCγ to the spinal cord can be achieved using lentiviral vectors and may be exploited as a therapeutic strategy in the treatment of neuropathic pain; this represents an interesting approach to refine the understanding of mechanisms through which PKCγ participates in pain processing in the spinal cord.
Intrathecal (IT) application of siRNAs is a suitable approach to analyze the function of receptors or other proteins hypothesized to play an important role in pain signaling (Finegold et al., 1999; Luo et al., 2005; Rohl and Kurreck, 2006; Doré-Savard et al., 2008; Towne et al., 2009; Zhang et al., 2009). Therefore, based on the effective target sequence of RNAi targeting PKCγ, we introduced lentiviral vectors encoding a specially designed shRNA against the PKCγ gene (LV-shPKCγ) into neurons and spinal cord of rats, and observed the inhibitory effect of the shRNA in vitro and in vivo. We evaluated the antinociceptive effect of IT administration of LV-shPKCγ on chronic constriction injury (CCI)–induced neuropathic pain in rats and probed the potential of intrathecally injected shRNA as a tool for functional investigation of PKCγ in neuropathic pain.
Materials and Methods
Lentiviral vectors
The design of siRNA followed previously published guidelines (Ui-Tei et al., 2004) and was performed using the Ambion application. To minimize off-target effects (Scherer et al., 2004), a BLAST homology search (based on sense and antisense sequences) was systematically performed to ensure that a single mRNA sequence was targeted (
Sense siRNA sequences targeting PKCγ (GenBank accession no. 012628) or control sequences were as follows: shPKCγ1 (1,119–1,137), AGTTTGAGGCCTGTAATTA; shPKCγ2 (1,343–1,361), CTCTATGCCATCAAGATAC; nonsilencing siRNA, TTCTCCGAACGTGTCACGT. The oligonucleotides were designed according to the structure of the siRNA sense strand-loop-siRNA antisense strand. The oligonucleotides (sense, 5'-CCGGGAAGTTTGAGGCCTGTAATTA
To generate the lentivirus, the recombinant work vector and package plasmids were cotransduced into 293T cells using Lipofectamine 2000 (Invitrogen, Carlsbad, CA). Lentivirus was generated as previously described (Coleman et al., 2003). The culture medium was collected over 48 hr, concentrated by ultracentrifugation, aliquoted, and stored at −80°C until used. Virus titer is the number of cells expressing green fluorescent protein (GFP) multiplied by the corresponding dilution, and the titer of lentivirus was determined by hole-by-dilution titer assay (Deglon et al., 2000). The final titer of recombinant virus was 1 × 109 transducing units (TU)/ml.
Cell culture, transduction, and infection
The 293T packaging cell line (Academy of Life Science, China) was maintained in Dulbecco's modified Eagle's medium (Invitrogen), supplemented with 10% fetal bovine serum (GIBCO) and antibiotics (100 mg/ml streptomycin and 100 U/ml penicillin) at 37°C in a humidified incubator containing 5% CO2. Cells were seeded in 24-well plates at 50–70% confluence 24 hr prior to transduction. To analyze transduction efficiency, 293T cells were gated to determine the percentage of GFP-positive cells. Cells with > 85% viability were cultured for additional experiments.
Primary neuronal cultures were obtained from the cerebral cortex of embryonic day 16 adult Sprague-Dawley rats, as described previously with minor modifications (Meberg and Miller, 2003). Transient transduction was performed using Lipofectamine Plus (Invitrogen) according to the manufacturer's instructions in six- or 12-well plates with cells at 70–90% confluence. All procedures were performed according to the local guidelines for animal research approved by the Administrative Committee of Experimental Animal Care and Use of Central South University.
Transduction of primary cultured neurons and GFP fluorescence observation
Neurons were plated in eight-well chamber slide dishes (Nalge Nunc International, Naperville, IL) coated with poly-D-lysine and laminin (Sigma–Aldrich, St. Louis, MO), and cultured in Neurobasal media (Invitrogen) with supplements. Dissociated neurons were infected with lentivirus vectors 1 hr after plating at a multiplicity of infection (MOI) of 10 TU/cell, yielding about 95% infection of neurons with no apparent toxicity. Forty-eight hours after infection, the cells were assayed for the expression of the transgenes. The GFP fluorescence in the cells was monitored with a fluorescent microscope (Leica, Solms, Germany) at 24, 48, and 72 hr post transduction. We collected duplicates of each condition for protein analysis 5 days after infection.
Real-time quantitative PCR analysis
Total RNA was extracted using an RNA isolation kit from Invitrogen following the manufacturer's instructions. The reactions were run on a real-time PCR system (iQ5 real-time PCR; Bio-Rad, Hercules, CA) with the following cycle conditions: 95°C for 15 sec, 45 cycles at 95°C for 5 sec and at 60°C for 30 sec. A standard curve for PKCγ was generated using serially diluted total RNA from neuronal cells and spinal cord and was used to quantify relative PKCγ mRNA levels. The sequences of the PKCγ gene primers were as follows: forward primer, 5’-TTCACAACCAGGGCATCATC-3’; reverse primer, 5’-AACATCTCATACAGCAGGACTC-3’; the product was 235 bp. β-Actin served as a control for normalization. The primers were as follows: forward primer, 5’-CGGCATTGTCACCAACTG-3’; reverse primer, 5’-CGCTCG GTCAGGATCTTC-3’; the product was 369 bp. The relative expression of mRNA was calculated by the 2–ΔΔCt method (Livak and Schmittgen, 2001).
Western blot analysis
Cells or dissected spinal cords were washed with cold phosphate-buffered saline (PBS) containing 2 mM EDTA and lysed with denaturing SDS-PAGE sample buffer using standard methods. The final concentration of protein in each sample was adjusted to 2 μg/μl. Protein lysates were separated by 12% SDS-PAGE and transferred onto polyvinylidene fluoride membrane (Millipore, Bedford, MA). The membranes were blocked with 5% skim milk for 1 hr at 37°C and then incubated with rabbit polyclonal anti-PKCγ antibody (dilution at 1:300; Santa Cruz Biotechnology, Santa Cruz, CA) at 4°C overnight. After the membranes were washed three times with Tris-buffered saline, they were incubated with horseradish peroxidase (HRP)–conjugated goat anti-rabbit immunoglobulin G (IgG) antibody (dilution at 1:5,000; Santa Cruz) at room temperature for 2 hr. Signal detection was carried out with an ECL system (Amersham Biosciences, GE Healthcare, Waukesha, WI). The band density was measured with Quantity One analysis software (Bio-Rad, Hercules, CA), and the quantification of PKCγ to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) levels was done by densitometry analysis.
Animals
Male Sprague-Dawley rats (250–300 g) were purchased from the Experimental Animal Center of the Central South University. All animals were housed at a controlled temperature of 20 ± 0.5°C in cages and maintained on a 12-hr light/dark cycle, with free access to food and water. Rats were allowed to acclimate for 1 week after arrival. In animal experiments, all efforts were made to minimize the suffering and number of animals used. All animal experimental procedures conformed to the guidelines established by the Council of the China Physiologic Society and were approved by the Administrative Committee of Experimental Animal Care and Use of Central South University. The study adhered to the Ethical Guidelines of the International Association for the Study of Pain (Zimmermann, 1983).
Chronic constriction injury
The surgery to generate the CCI model was first described by Bennett and Xie (1988). In brief, each rat was anesthetized by chloral hydrate (300–350 mg/kg, i.p.), and the left common sciatic nerve was exposed at mid-thigh level by blunt dissection through the biceps femoris. Proximal to the sciatic trifurcation, four ligatures (4-0 chromic gut) with about 1-mm spacing were loosely tied around the nerve. At the time of tying, the ligatures just barely reduced the nerve diameter. A sham surgery was performed with the sciatic nerve exposed but not ligated. The incision was closed in layers. Upon recovery from anesthesia, the animals were housed postoperatively in individual clear plastic cages with solid floors covered with 3–6 cm of soft bedding (sawdust), with free access to food and water. Animals that had undergone CCI surgery and demonstrated a vigorous mechanical and thermal hypersensitivity effect of the nerve injury were used for further experiments.
Intrathecal catheter implantation
IT catheters were implanted as described by Yaksh and Rudy (1976). In brief, under anesthesia with chloral hydrate (300–350 mg/kg, i.p.), the occipital muscles were bluntly separated, and then the cisternal membrane was exposed. Microspinal catheters (inner diameter 0.12 mm, outer diameter 0.35 mm) were inserted into the subarachnoid space via an incision in the cisterna magna and advanced 8.0–8.5 cm caudally to the level of the lumbar enlargement region. Subcutaneously, 30,000 units of natrium penicillin (Huabei Pharmaceuticals, Shijiazhuang, China) was given to prevent infection. Correct IT placement was confirmed by the dragging or paralysis of bilateral hind limbs after injection of 10 μl of 2% lidocaine. The incision site was closed in layers, and the catheter was fixed firmly under the skin and sealed effectively. The rats were allowed to recover for 3 days before the CCI test and monitored daily after surgery for signs of motor deficiency or infection.
Experimental animal grouping
Sprague-Dawley rats weighting 260–320 g were randomly divided into six groups: group sham (Sham), group CCI +normal saline (NS), group CCI + pGCSIL-GFP (LV-NC), group CCI + pGCSIL-GFP-ShPKCγ1 (LV-shPKCγ1, 10 μl or 5 μl), and group CCI + pGCSIL-GFP-ShPKCγ2 (LV-shPKCγ2, 10 μl or 5 μl). The animals were anesthetized with intraperitoneal chloral hydrate (300–350 mg/kg). After the vector (10 μl or 5 μl) was administered, the catheter was flushed with 5 μl or 10 μl of PBS. Rats in each group were intrathecally treated with NS, LV-NC, or LV-shPKCγ plus PBS in a total volume of 15 μl. One week after IT administration, animals in the LV-NC and LV-shPKCγ groups were sacrificed, and the L4–5 lumbar segment of the spinal cord was removed for determination of GFP expression (Garraway et al., 2009). The expression of PKCγ mRNA and PKCγ protein in the spinal cord was determined by real-time RT-PCR and Western blot analysis 7 days after IT delivery (six animals in each group). Eight animals in the Sham, NS control, LV-NC, LV-shPKCγ1, and LV-shPKCγ2 groups were tested to measure pain thresholds.
Immunohistochemistry
Seven days after IT delivery, rats treated with vehicle or LV-shPKCγ were deeply anesthetized with chloral hydrate (500 mg/kg, i.p.) and perfused transcardially with 100 ml of PBS (composition in mM: NaCl 137, KCl 2.7, KH2PO4 1.5, NaH2PO4 8.1; pH 7.4), followed by 250 ml of ice-cold 4% paraformaldehyde. The lumbar (L4–5) segment of the spinal cord sections was removed and postfixed at 4°C for 5 hr and then transferred to 30% sucrose/PBS for 24 hr. Transverse L4–5 spinal cord sections (20 μm) were incubated for 2 hr at room temperature in a blocking solution (3% normal goat serum) and then incubated for 48 hr at 4°C with rabbit polyclonal anti-PKCγ antibody (dilution at 1:500; Santa Cruz). Following incubation, tissue sections were washed and incubated for 3 hr at room temperature in the secondary antibody solution HRP-conjugated goat anti-rabbit IgG antibody (dilution at 1:2,000; Santa Cruz).The spinal cord sections were analyzed using an LSM confocal imaging system (Carl Zeiss Japan, Tokyo, Japan).
Measurement of pain thresholds
Pain threshold testing was performed by experimenters blinded to the drugs being administered. Paw mechanical withdrawal thresholds (PMWT) were determined in response to pressure from an electronic von Frey anesthesiometer (2390 series; IITC Instruments, Woodland Hills, CA) as described previously (Chaplan et al., 1994). The amount of pressure (in grams) needed to produce a paw withdrawal response was measured three times on each paw, separated by 3-min intervals. The three tests were averaged for each paw for a given day.
The thermal nociceptive threshold [paw withdrawal thermal latency (PWTL)] was detected by a Hargreaves apparatus (Plantar test, 7370; Ugo Basile, Comerio, Italy) according to a previous method (Patwardhan et al., 2006). Rats were placed in clear plastic cages on an elevated glass plate and allowed to acclimate to their surroundings for 30 min before testing. After acclimation, a constant-intensity, radiant heat source was focused from underneath the glass and aimed at the mid-plantar area. A digital timer automatically read the duration between the start of stimuli and paw withdrawal. The PWTL was measured to the nearest 0.1 sec. The baseline PWTL of rats was controlled at about 11 sec by adjusting the light intensity of the pain threshold detector. A cutoff time of 15 sec of irradiation was used to avoid any tissue damage. Five minutes was allowed between stimulations. PMWT and PWTL were measured 1 day before CCI (baseline) and 0, 3, 7, 14, 28, 35, and 42 days after IT administration.
Statistical analysis
Data are presented as means ± SD. One-way analysis of variance (ANOVA) with Student-Newman-Keuls post hoc analysis was used to detect differences of expression of PKCγ mRNA and protein among groups. Two-way repeated-measures ANOVA was used to detect differences of pain thresholds among groups. Differences are deemed significant at p < 0.05.
Results
PCR identification of constructed shRNA expression plasmids and titer determination and packaging of the lentiviral vector
The pGCSIL-GFP PKCγ shRNA expression plasmids were identified using a PCR protocol, rather than restriction endonuclease digestion, because the length of the inserted fragment was only 37 bp. The lengths of the PCR products of the successfully constructed shRNA expression plasmids and the empty pGCSIL-GFP vector were 343 and 306 bp, respectively (Fig. 1). Results of DNA sequencing were as expected, and the recombination vector of lentivirus encoding the specially designed shRNA against the PKCγ gene was named pGCSIL-shPKCγ-GFP (LV-shPKCγ).
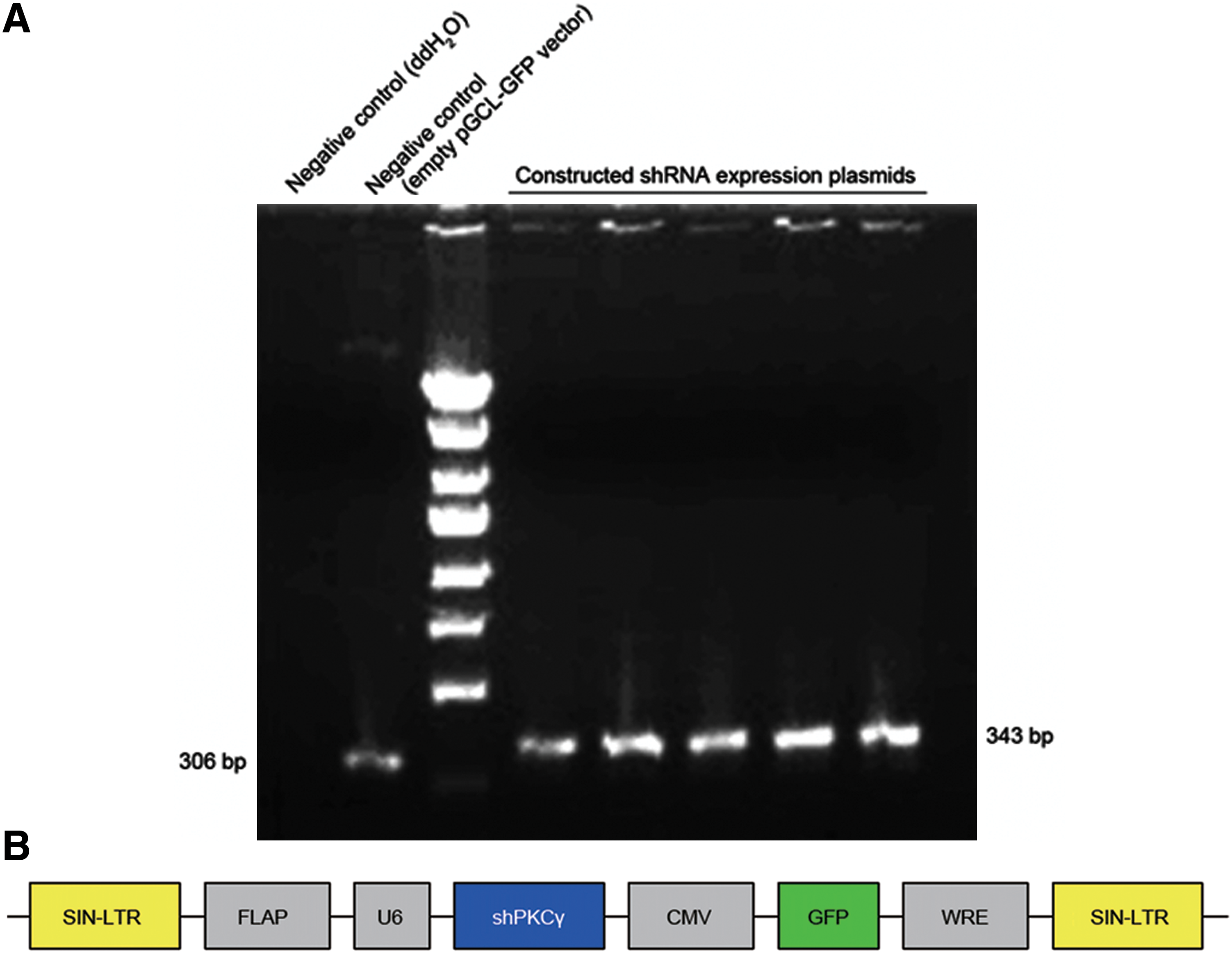
PCR identification of constructed shRNA expression plasmids and schematic representation of the LV-shPKCγ-CMV-GFP expression vector. (
Forty-eight hours after cotransfection of the three-plasmid lentiviral vector into 293T cells, strong green fluorescence was observed using an inverted fluorescence microscope. After a single exposure of 293T cells to the lentivirus, a high percentage (>90%) of transfectants expressed GFP at 48 hr after the transduction, indicating a high and stable transduction of the lentiviral vector system.
Lentivirus containing a PKCγ shRNA and GFP were transduced in vitro and in vivo
The lentiviral vector was transduced into neuronal cells in vitro for 72 hr, and no significant changes in cell morphology and no cell death were observed, indicating that the lentiviral vector has no deleterious effect on neurons. There was no difference between the LV-shPKCγ, the lentiviral vectors encoding PKCγ shRNA negative control, and pGCSIL-shPKCγNC-GFP (LV-NC) groups. Under the fluorescent microscope, the strong GFP was visible in neuronal cells 72 hr after transduction with LV-shPKCγ or LV-NC, indicating that recombinant lentivirus led to a successful transduction of neurons (Fig. 2A and B).
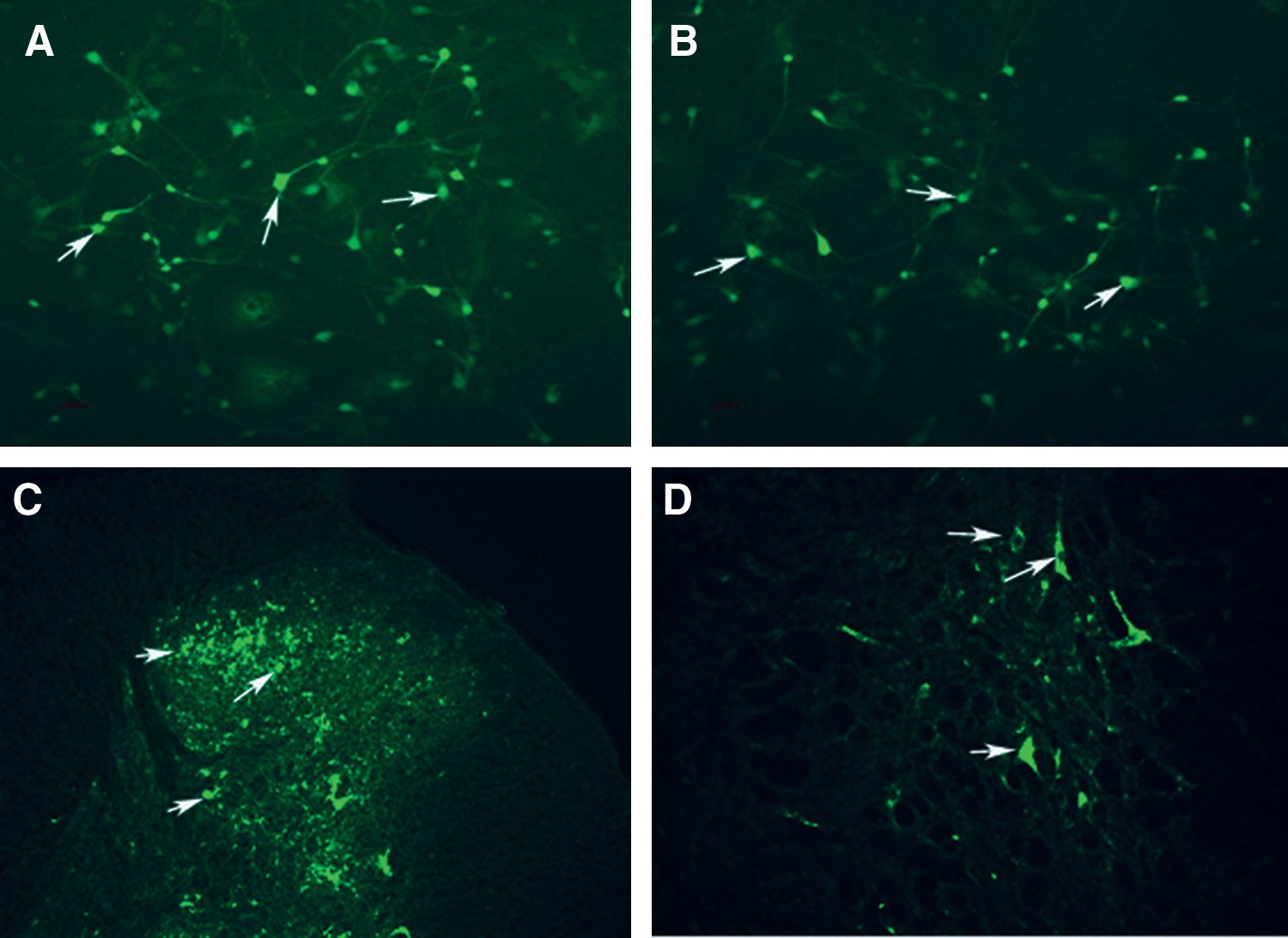
(
The neurons were transduced with the indicated lentiviral vectors at an MOI of 10. After 3 days of transduction, transduction efficiency was measured by the frequency of GFP-positive cells. As demonstrated by the representative experiment shown in Fig. 2, >95% of neurons were successfully transduced.
To confirm the delivery of LV-shPKCγ into neuronal cells in the spinal cord, the uptake of labeled GFP in spinal cord cells was evaluated by fluorescence microscopy. Many cell bodies were highly fluorescent after IT injection of LV-shPKCγ or LV-NC. Fluorescence was observed primarily in the cytoplasm (Fig. 2C and D). The fluorescence intensity was variable among the cell bodies irrespective of cell size.
Lentivirus-mediated delivery of shRNA inhibits PKCγ expression in neuronal cells in vitro
To evaluate the inhibition of PKCγ mRNA expression, real-time RT-PCR was performed 72 hr after transduction. Figure 3A shows that LV-shPKCγ1 and LV-shPKCγ2 transduction resulted in about 95.2% and 96.4% reduction, respectively, of PKCγ mRNA expression compared with the negative controls including LV-NC and medium controls (p < 0.05), indicating that the mRNA sequences corresponding to the PKCγ shRNA were specific RNAi targets. Western blot analysis was performed 72 hr after transduction and showed that LV-shPKCγ1 and LV-shPKCγ2 induced an 85.3% and 86.5% down-regulation, respectively, of the PKCγ protein level compared with LV-NC (p < 0.05) (Fig. 3B and C).
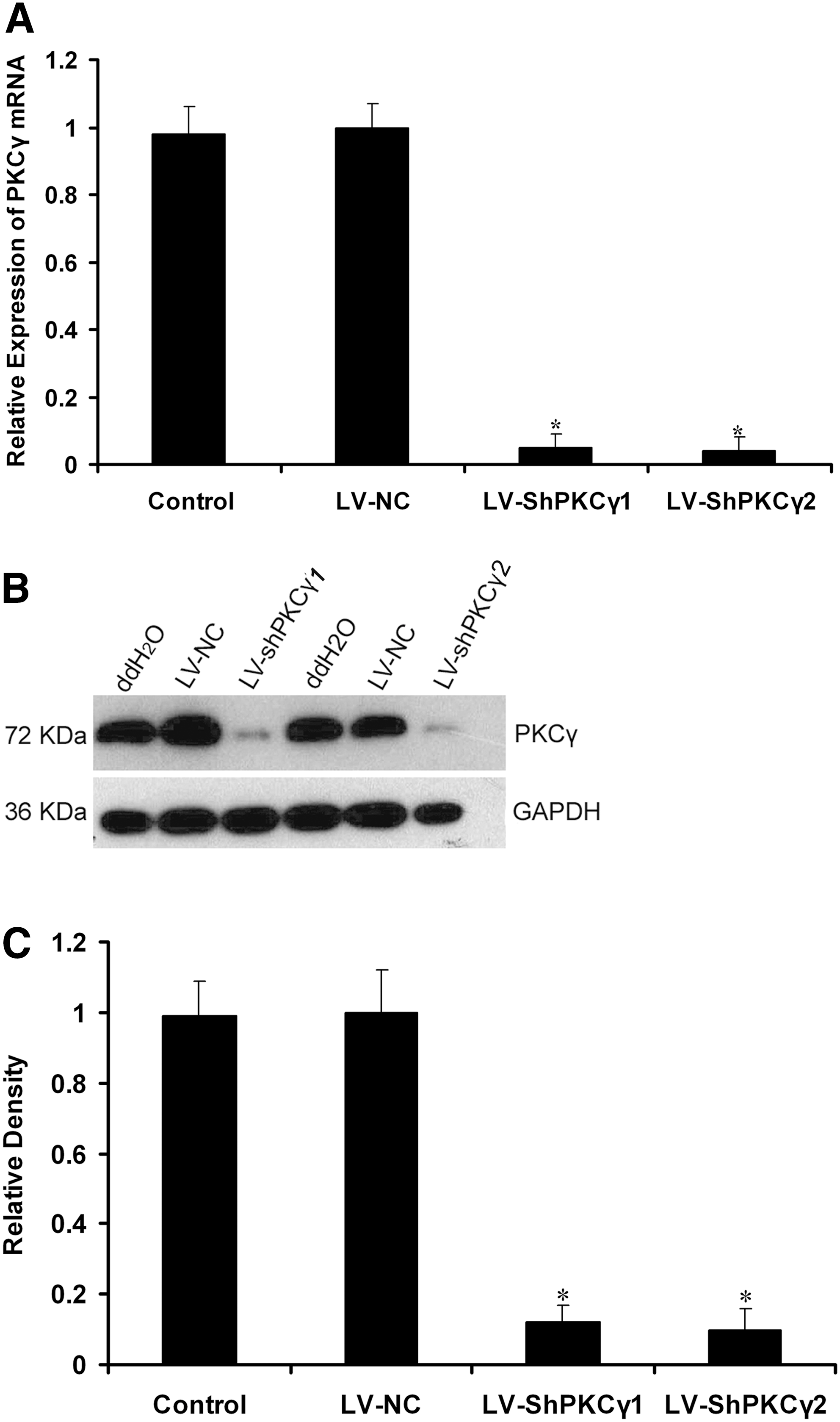
Real-time PCR analysis of the level of PKCγ mRNA in neurons. ddH2O was used as a negative control. (
Lentivirus-mediated delivery of shRNA inhibits PKCγ expression in the dorsal spinal cord in vivo in a dose-dependent manner
To evaluate the inhibition of PKCγ mRNA expression in the dorsal spinal cord, real-time RT-PCR was performed 7 days after IT delivery. LV-shPKCγ1 and LV-shPKCγ2 (10 μl) transduction resulted in 75.5% and 76.2% reduction, respectively, of PKCγ mRNA compared with the LV-NC and blank control groups (p < 0.05) (Fig. 4A). Western blot analysis was performed 7 days after IT delivery. PKCγ protein expression was significantly down-regulated in LV-shPKCγ1 and LV-shPKCγ2 groups (interference efficiency was up to 76.6% and 77.3%, respectively) (Fig. 4B and C) compared with the LV-NC and blank control groups. Similarly, PKCγ mRNA and protein expression was also significantly down-regulated in LV-shPKCγ1 and LV-shPKCγ2 (5 μl) (mRNA interference efficiency was up to 65.9% and 66.7%, and protein interference efficiency was up to 67.1% and 66.2%, respectively) compared with the LV-NC and blank control groups (p < 0.05) (Fig. 4A–C), whereas no obvious inhibition of PKCγ protein was observed in the NS control group and LV-NC group (p < 0.05).
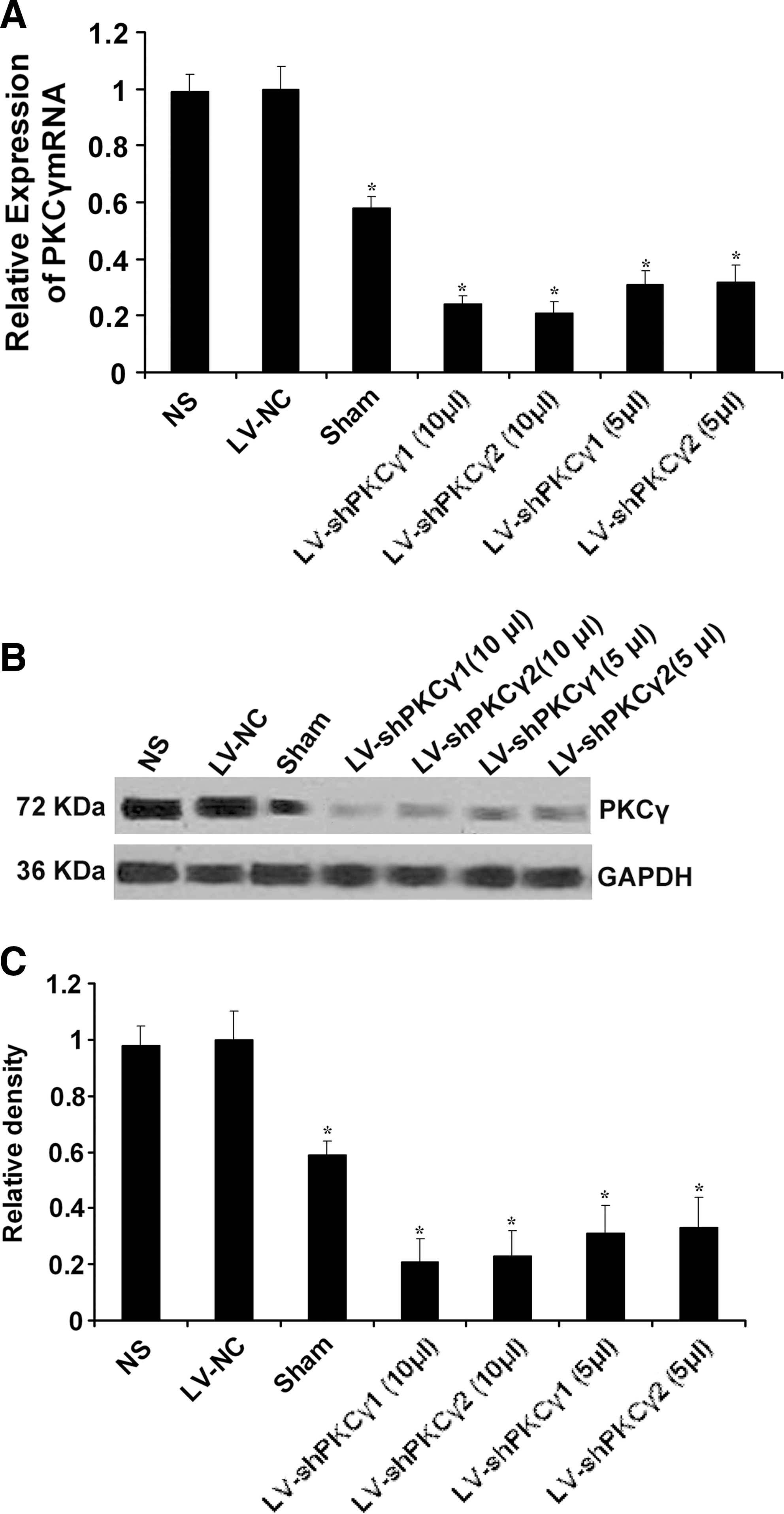
PKCγ mRNA expression in the spinal cord after IT delivery of NS, LV-NC, LV-shPKCγ1, and LV-shPKCγ2. (
PKCγ immunoreactivity in the dorsal horn of the rat spinal cord after LV-NC, LV-shPKCγ (10 μl), or LV-shPKCγ (5 μl) treatment
The immunoreactivity of PKCγ was found predominantly in the superficial laminae of the dorsal horn of the lumbar spinal cord. The immunostaining was significantly lower in tissues from LV-shPKCγ–treated rats than from LV-NC–treated animals (p < 0.05), and IT injection of LV-shPKCγ resulted in a significant reduction of immunoreactivity of PKCγ in a dose-dependent manner.
The expression of PKCγ decreased dramatically in the dorsal horn of the lumbar spinal cord in LV-shPKCγ–treated rats as compared with that in LV-NC–treated rats (p < 0.05; Figs. 4 and 5). Furthermore, IT injection of LV-shPKCγ inhibited PKCγ expression in the dorsal spinal cord in vivo in a dose-dependent manner.
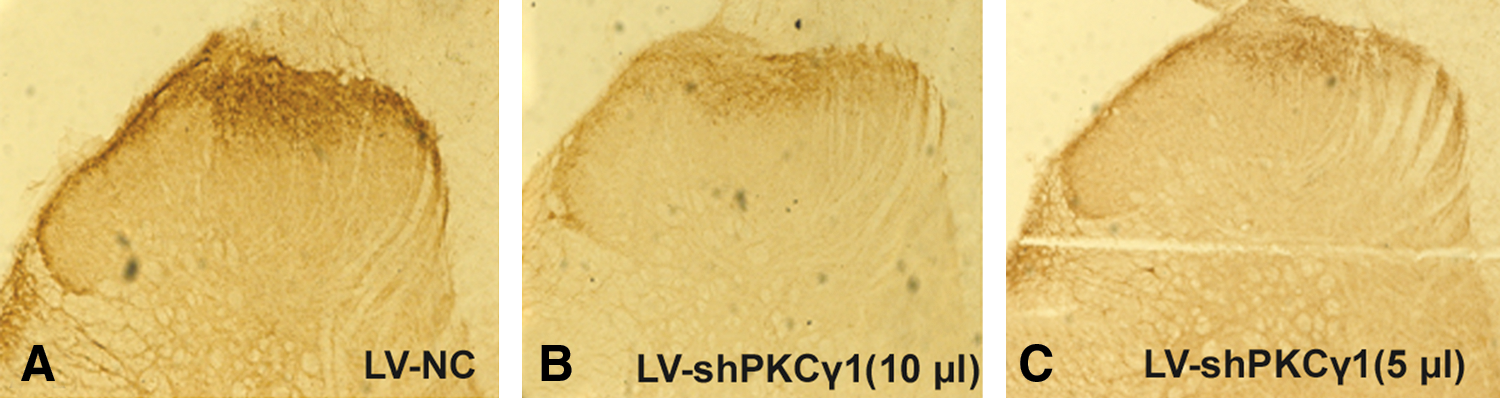
PKCγ immunoreactivity in the dorsal horn of the rat spinal cord after LV-NC, LV-shPKCγ (10 μl), or LV-shPKCγ (5 μl) treatment. The immunoreactivity of PKCγ was found predominantly in superficial laminae of the dorsal horn. PKCγ immunoreactivity was decreased significantly in LV-shPKCγ–treated animals (
Lentivirus-mediated delivery of shRNA reduced tactile allodynia and thermal hyperalgesia in a neuropathic pain model in rats
Following the injection of vectors, we observed the behavior of the rats for 6 weeks. The rats had a normal appearance and level of activity and fed regularly. LV-shPKCγ1 and LV-shPKCγ2 significantly reduced mechanical allodynia and thermal hyperalgesia in CCI animal models after administration of a 10-μl and 5-μl dose of IT LV-shPKCγ1 or LV-shPKCγ2 vectors, compared with the response after administration of LV-NC (Fig. 6A and B). Groups treated with LV-shPKCγ1 and LV-shPKCγ2 (10 μl) demonstrated a significant increase in pain thresholds as compared with groups treated with LV-shPKCγ1 and LV-shPKCγ2 (5 μl). This result correlated with the diminished PKCγ protein level revealed by Western blot analysis (Fig. 4B and C). Compared with the responses of the untreated group, the administration of LV-NC alone or NS did not alter the latency of paw withdrawal from a painful thermal stimulus.
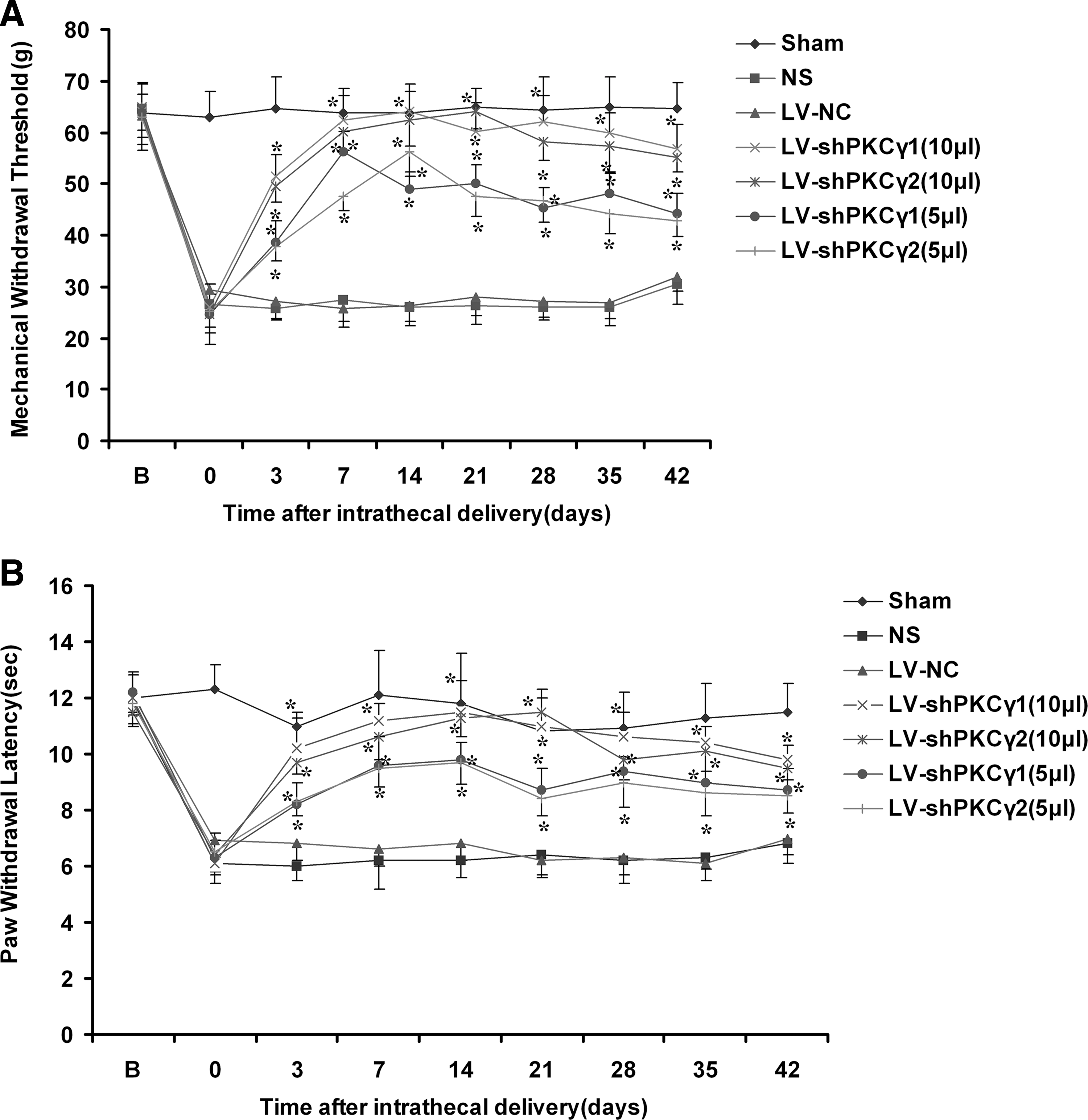
Tactile allodynia (
Discussion
The PKC family is classified into three subgroups, based on structural differences in their regulatory domains and mode of action. Conventional or classic-type PKCs (cPKC-α, βI, βII, and γ) are activated by Ca2+, diacylglycerol (DAG), and phorbol esters. Novel-type PKCs (nPKC-ɛ, δ, η, and θ) are activated by DAG and phorbol esters, but not Ca2+. Atypical-type PKCs (aPKC-λ and ξ) are unaffected by Ca2+, DAG, or phorbol esters, but are activated by phospholipids such as phosphatidylserine or phosphatidylinositol 3,4,5-triphosphate (Way et al., 2000; Velazquez et al., 2007). It is well recognized that the up-regulation of PKCγ activity in the spinal cord is a key factor for the induction of central sensitization, especially in neuropathic pain following nerve ligation.
The study demonstrates that IT delivery of lentiviral vectors is capable of silencing PKCγ and provides a considerable antinociceptive therapeutic effect. In this study, the top priority was how to select the correct target for therapy. We chose PKCγ, which is considered a central molecular integrator of nociceptive signaling (Malmberg et al., 1997; Martin et al., 2001; Celerier et al., 2004). Many studies have indicated that some enhanced processes of reactivities, such as hyperalgesia, may be related to central sensitization. During the formation of central sensitization, PKCγ plays a key role and is an important molecular target for gene therapy on chronic pain (Wu et al., 2001). In the study, we observed that CCI-induced neuropathic pain rats showed significant increases in both PKCγ mRNA and protein levels; this is because peripheral nerve injury not only produces a neuropathic pain syndrome, but also significantly up-regulates PKCγ activity in the spinal cord dorsal horn, which is consistent with previous studies (Miletic et al., 2000; Mao et al., 1995).
RNAi-based knockdown can be generated by the expression of a vector-mediated RNAi anywhere in the genome (Zhai et al., 2009). Less than a decade after its discovery, RNAi-mediated gene silencing is already being tested as a potential therapy in clinical trials for a number of diseases (Shrey et al., 2009). Previous studies applied shRNAs locally into the spinal cord to investigate the functional role of pain-related receptors. It was demonstrated that shRNAs were either continuously infused via a minipump (Dorn et al., 2004) or were mixed with delivery agents like the transduction reagent i-Fect (Luo et al., 2005) or a polyethylenimine-based gene delivery system (Suzuki et al., 2009). On the other hand, viral delivery of shRNA expression cassettes allows efficient transduction in tissues such as the brain and liver. Recombinant adeno-associated virus vectors expressing shRNA were used to knock down the mutant gene (Franich et al., 2008). Anesti et al. (2008) demonstrated an efficient delivery of RNAi to peripheral neurons in vivo using herpes simplex virus. Therefore, viral-based vectors have been developed as an alternative strategy.
The second concern was how to choose the optimal vector. We selected lentiviral vectors as a gene delivery tool as they are able to induce limited inflammation, provide a means to express shRNA to induce stable and long-term gene silencing in both dividing and nondividing cells, and have a relatively large cloning capacity. Thus, lentiviral vectors are probably the best choice currently available for delivering and stably expressing shRNAs or siRNAs in target cells. The use of lentiviral vectors allows for a rapid and convenient way to stably express shRNAs in order to analyze the effects caused by knockdown of a specific gene (Xia et al., 2004; Vandekerckhove et al., 2006). Lentiviral vectors are an effective tool for the treatment of neuropathic pain, which can mediate different molecular target genes for chronic pain gene therapy (Pezet et al., 2006; Meunier et al., 2007 , 2008; Meunier and Pohl, 2009).
To construct LV-shPKCγ, we used a virus packaging system including plasmids pGCSIL-GFP, pHelper 1.0, and pHelper 2.0, of which pGCSIL-GFP contains elements to constitutively express small RNAs and GFP. The plasmid pHelper 1.0 contains the gag gene from the HIV virus (a main structural protein), the pol gene (a specific enzyme for the encoding virus), and the rev gene (a factor to regulate expression of the gag and pol genes). The plasmid pHelper 2.0 contains the vesicular stomatitis virus glycoprotein (VSVG) gene originating from the herpes simplex virus, which provides the capsid protein for virus packaging (Kafri et al., 1997). As a packaging cell, 293T generates high-titer virus particles. In this study, using the third-generation lentivirus of virus-replicated self-inactivation as a vector, we constructed LV-shPKCγ, whose titer was 1 × 109 TU/ml. These vectors contain GFP, which can be used as a reporter to verify the expression of the vector. Then primary cultured neurons were transduced by lentiviral vectors in vitro. After 72 hr, we observed high levels of GFP in a large number of neurons, suggesting that the recombinant lentiviral vector was successfully transduced into neuronal cells. Our results show that LV-shPKCγ induced an 85% down-regulation of PKCγ protein levels compared with LV-NC.
The third concern was to determine the site for therapeutic intervention. Clinically, IT administration of analgesics is commonly used for advanced cancer, surgery and postoperative analgesia, labor analgesia, and other noncancer chronic pain treatment. Therefore, IT administration to reduce pain at the level of the spinal cord is a commonly used method of gene therapy for chronic pain (Yao et al., 2003; Beutler et al., 2005; Luo et al., 2005; Ledeboer et al., 2007).
LV-shPKCγ reduced mechanical allodynia and thermal hyperalgesia in a neuropathic pain model. These were significantly inhibited in the neuropathic paw (partially ligated sciatic nerve) in the LV-shPKCγ– versus LV-NC– or NS-treated rats. In comparison with previous reports, our results are novel in three important ways. First, an endogenous disease-related neuronal gene, PKCγ, in the nervous system was targeted. Second, LV-shPKCγ was delivered intrathecally to the spinal fluid, a delivery method commonly used for the clinical treatment of severe pain. Third, these data have immediate implications in the study of genes for in vivo target validation in drug discovery.
In this study, after spinal cord neurons were transduced using LV-shPKCγ and LV-NC in vivo, spinal dorsal horn cells demonstrated strong GFP expression (Chalfie, 1995), suggesting that recombinant lentiviral vectors carrying GFP can be successfully transduced into target cells, resulting in interference effects. The results indicate that the use of lentiviral-mediated delivery of shRNA targeting PKCγ decreased the expression of PKCγ mRNA and its associated protein, as demonstrated by real-time PCR and Western blotting, respectively. In addition, PKCγ targeting also attenuates CCI-induced mechanical allodynia and thermal hyperalgesia for at least 6 weeks in rats. Furthermore, IT injection of LV-shPKCγ inhibits PKCγ expression in the dorsal spinal cord and attenuates pain thresholds in a dose-dependent manner.
PKCγ has been found to potentiate N-methyl-D-aspartate (NMDA)–induced currents in the spinal cord via alleviation of its voltage-dependent Mg2+ block, and thus provides an ideal mechanism for PKC-dependent central sensitization. This evidence supports a role for PKC in directly mediating neuronal excitability within the spinal cord. LV-shPKCγ provides long-term inhibition of PKCγ. The constitutive suppression of PKCγ may inhibit phosphorylation of the NMDA receptor, which leads to weakening of the NMDA-mediated multiple excitatory neurotransmitter and neuromodulatory peptides released at the primary afferent-spinal cord synapse. Finally, long-term potentiation of synaptic transmission was achieved, resulting in suppression of CCI-induced mechanical allodynia and thermal hyperalgesia (Malmberg et al., 1997; Martin et al., 2001; Labombarda et al., 2008).
In the present study, we found that IT administration of LV-shPKCγ significantly attenuated CCI-induced neuropathic pain in rats but failed to have obvious effects on the general behavior. We compared the weight, food intake, drinking water, and reactive ability of the rats after LV-shPKCγ injection and found that there was no difference among the LV-shPKCγ–, LV-NC–, and NS-treated rats. However, some studies reported that mice with the deletion (knockout) of PKCγ have a slight ataxia, modest learning and memory impairments, and some motor incoordination, which may be related to the elimination of multiple climbing fiber innervation of Purkinje cells (Chen et al., 1995; Kano et al., 1995). In addition, some scholars reported that PKCα, but not PKCβ or PKCγ, regulates contractility and heart failure susceptibility, indicating that PKCγ knockout did not affect cardiovascular functions (Liu et al., 2009).
Considering its effectiveness in vivo, our data indicate that shRNA could be used extensively to test the involvement of potential novel targets in disease progression. This approach is particularly attractive when drugs or small-molecule compounds directed at these targets are not available. On a more speculative level, there is the possibility that shRNA can be used as a therapy for certain disease states where delivery and target protein knockdown can be optimized. Therefore, these results suggest that IT delivery of LV-shPKCγ can alleviate allodynia and hyperalgesia through a PKCγ-related pathway without profound side effects. The PKCγ pathway may play a pivotal role in chronic neuropathic pain. These results support the potential use of shRNA expression vectors as a gene therapy approach to neuropathic pain.
In summary, IT delivery of lentiviral-mediated RNAi targeting PKCγ effectively attenuates CCI-induced neuropathic pain in rats. The present study suggests that PKCγ is a feasible RNAi target for neuropathic pain. Furthermore, lentiviral vector delivery strategies are a promising novel approach for the treatment of neuropathic pain and for future studies on gene functions of PKCγ.
Footnotes
Acknowledgments
This study was supported by a grant from the National Science Council of China (30801074, 30772080).
Author Disclosure Statement
No competing financial interests exist.