
Abstract
Effective therapeutic strategies for radiation-induced lung injury (RILI) are lacking. Mesenchymal stem cells (MSCs), as gene therapy delivery vehicles, possess the ability to repair injured lung. In this study, we conducted MSC-based hepatocyte growth factor (HGF) gene therapy for RILI. Mice received single-dose radiation with 20 Gy of γ rays locally to the lung, and then were administered normal sodium, Ad-HGF-modified MSCs, or Ad-Null-modified MSCs. Ad-HGF-modified MSCs (MSCs-HGF) improved histopathological and biochemical markers of lung injury. MSCs-HGF could reduce secretion and expression of proinflammatory cytokines, including tumor necrosis factor-α, interferon-γ, interleukin (IL)-6, and intercellular adhesion molecule-1, and increase the expression of antiinflammatory cytokine IL-10. It could also decrease expression levels of profibrosis factors transforming growth factor-β, Col1a1 (collagen type 1, α1), and Col3a1, and inhibit fibrosis progress. MSCs-HGF could promote proliferation of lung epithelial cells and protect them from apoptosis, and improve the expression of endogenous HGF and its receptor c-Met significantly. We also found that sphingosine-1-phosphate receptor-1 expression was increased in injured lung. These results suggest MSC-based HGF gene therapy not only reduces inflammation but also inhibits lung fibrosis.
Introduction
Studies have demonstrated that bone marrow-derived mesenchymal stem cells (MSCs) engrafted in the injured lung can differentiate into lung epithelial cells in vivo (Kotton et al., 2001; Grove et al., 2002; Yamada et al., 2004). In addition, it has been reported that MSCs or their secreted products can reduce lung inflammation and improve lung structure in experimental rodent models of hyperoxia-induced bronchopulmonary dysplasia (Aslam et al., 2009; van Haaften et al., 2009). Importantly, MSCs can serve as cellular vehicles for gene delivery and are being developed to treat various diseases, including acute lung injury (Grove et al., 2002; Reiser et al., 2005; Mei et al., 2007).
Hepatocyte growth factor (HGF), originally identified and cloned as a potent mitogen for mature hepatocytes (Matsumoto and Nakamura, 1992), has mitogenic, morphogenic, and antiapoptotic activities in a wide variety of cells, including most epithelial and endothelial cells. It is noteworthy that HGF enhances the regeneration of organs such as the liver, kidney, and lung (Matsumoto and Nakamura, 1997). HGF also has an inhibitory effect on fibrosis. Administration of HGF protein or HGF gene therapy has improved renal fibrosis and dysfunction (Mizuno et al., 2008). However, a high level of expression of HGF in the injured areas of radiated lung is unachievable.
In this study, we evaluate the therapeutic effects of MSC-based HGF gene expression on radiation-induced lung injury (RILI), using an in vivo C57BL/6 mouse model.
Materials and Methods
Isolation, culture, and characteristics of bone marrow-derived mesenchymal stem cells
Bone marrow (BM) aspirates were obtained from adult volunteer donors in accordance with the ethical standards of the local ethics committee. BM mononuclear cells (MNCs) were separated by Ficoll density gradient (1.077 g/ml) centrifugation, and then were cultured in minimum essential medium alpha (α-MEM; HyClone, Logan, UT) plus screened 10% fetal bovine serum (FBS; HyClone) for MSC expansion at a density of 1×106 cells/cm2. The cells were cultured at 37°C in a humidified atmosphere containing 5% CO2, and the culture medium was changed every 3 days until the cells reached 70–90% confluency. On isolation, MSCs were expanded for 2–4 passages. Their immunophenotypes were detected by flow cytometry (BD Biosciences, San Jose, CA), and showed positive for HLA-ABC, CD73, CD29, and CD166 and negative for CD31, CD34, CD45, and HLA-DR (Supplementary Fig. S1; supplementary data are available online at
Adenoviral vectors
The adenoviral vectors (AdVs) used in this study were as follows: Ad-HGF, a replication-defective adenovirus expressing human HGF (hHGF); Ad-Luc, a replication-defective adenovirus expressing luciferase; and Ad-Null, a replication-defective adenovirus not carrying exogenous genes. All AdVs were constructed with the AdEasy system (Stratagene, La Jolla, CA). Briefly, the human HGF gene or luciferase gene was amplified by PCR and inserted into the pShuttle-CMV, obtaining the pShuttle plasmid pShuttle-HGF or pShuttle-Luc. pShuttle-HGF, pShuttle-Luc, and pShuttle-CMV were then used to generate Ad-HGF, Ad-Luc, and Ad-Null, respectively, according to the manual accompanying the AdEasy system. All AdVs were purified by double CsCl density gradient ultracentrifugation, dissolved in storage buffer (Hanks' buffer, 10% glycerol), and stored at −80°C. Viral particle (VP) numbers and infectious titers (infectious units per milliliter, IU/ml) were determined as described previously; and the multiplicity of infection (MOI) was calculated from infectious titers (Supplementary Table S1).
MSCs were infected with 150 multiplicities of infection of Ad-HGF (MSCs-HGF) or Ad-Null (MSCs-Null) and cells were collected 48 hr postinfection for use as RILI therapy. HGF expression was detected in vitro by ELISA (Supplementary Fig. S2).
Animals
All animal experiments were conducted according to protocols approved by the Institutional Animal Care and Use Committee of the Beijing Institute of Radiation Medicine (Beijing, China). One hundred and twenty 8-week-old female mice received single-dose radiation with 20 Gy of γ rays locally to the lung and were then randomized into three groups: radiation, MSCs-Null, and MSCs-HGF. Saline (100 μl), MSCs-Null (1×106/100 μl saline), or MSCs-HGF (1×106/100 μl saline), respectively, were injected intravenously via the tail vein 6 hr after radiation. Another 40 mice were used as nonradiation controls. Mice were killed immediately (day 0) after treatment, or on day 1, 3, 7, 14, 28, or 180 after treatment. Bronchoalveolar lavage fluid (BALF) was collected on day 28 for determination of total protein, albumin, and IgM concentration. Serum was collected on days 7, 14, and 28 after treatment for determination of cytokines. The left lungs were removed and used for RNA or genomic DNA isolation and protein extraction; the right lungs were fixed in 4% paraformaldehyde for histopathological analysis.
Lung morphology
Hematoxylin–eosin and Masson staining
Paraffin-embedded lungs were sectioned at a thickness of 5 μm and stained with hematoxylin and eosin (H&E). Lung pathology was evaluated by light microscopy, according to four criteria: alveolar congestion, hemorrhage, infiltration or aggregation of neutrophils in airspaces or vessel walls, and thickness of alveolar wall. Collagen deposition was detected by Masson staining and digital imaging microscopy.
Immunohistochemistry
Paraffin-embedded lung sections were used for immunohistochemistry. Bcl-2, proliferating cell nuclear antigen (PCNA), and mouse HGF (mHGF) receptor c-Met were detected with rabbit anti-Bcl-2 antibody (ZhongShan Golden Bridge Biotechnology, Beijing, China), rabbit anti-PCNA antibody (Abcam, Cambridge, UK), and rabbit anti-c-Met antibody (Boster Biological Technology, Wuhan, China), respectively, and goat anti-rabbit IgG conjugated with horseradish peroxidase, using a 3,3′-diaminobenzidine (DAB) kit (ZhongShan Golden Bridge Biotechnology). For terminal deoxynucleotidyltransferase-mediated nick-end labeling (TUNEL) of the lung, we used the DeadEnd fluorometric TUNEL system (Promega, Madison, WI) according to the manufacturer's instructions.
Mouse cells expressing c-met and bcl-2 show the proteins Buffy within the cytoplasm, and their nuclei turn light blue. For PCNA staining and TUNEL, positive cells show Buffy within the nucleus, whereas the nuclei of negative cells turn light blue. In addition, positive cells in stained tissue sections were quantified on the basis of five random ×400 images per group. The proliferation rate and apoptosis index were determined as the ratio of positive cells to total cells.
Measurement of total protein, albumin and IgM in BALF
The total protein concentration in each sample was determined with a bicinchoninic acid (BCA) kit (Pierce Biotechnology/Thermo Fisher Scientific, Rockford, IL). Albumin and IgM levels in BALF samples were measured with a murine-specific albumin ELISA kit (ALPCO Diagnostics, Salem, NH) and a murine-specific IgM ELISA kit (Bethyl Laboratories, Montgomery, TX).
Measurement of cytokine expression
Levels of cytokines (tumor necrosis factor [TNF]-α, intercellular adhesion molecule [ICAM]-1, and transforming growth factor [TGF]-β) were measured with murine cytokine-specific Quantikine ELISA kits (R&D Systems, Minneapolis, MN) according to the manufacturer's instructions.
Detection and quantitative analysis of engrafted hMSCs in lung
Biological samples underwent DNA extraction and PCR analysis to quantify the numbers of human cells in mouse recipients. Genomic DNA for PCR analysis was prepared from lung tissues, using a QIAamp DNA mini kit (Qiagen, Valencia, CA). The human β-globin gene and the endogenous mouse receptor-associated protein at the synapse (RAPSYN) gene were amplified with Premix Ex Taq (probe qPCR) (Takara Bio, Otsu, Japan). For human β-globin, the forward primer was 5′-GTGCACCTGACTCCTGAGGAGA-3′, the reverse primer was 5′-CCTTGATACCAACCTGCCCAGG-3′, and the probe, labeled with fluorescent reporter and quencher, was 5′-FAM-AAGGTGAACGTGGATGAAGTTGGTGG-TAMRA-3′. For mouse RAPSYN, the forward primer was 5′-CCTTAGCCAATTGGAGAACA-3′, the reverse primer was 5′-TTGGCCAGTTTAAAACCCAT-3′, and the probe was 5′-FAM-TATCTGACCCACCCATCCTGC-TAMRA-3′.
Real-time reverse transcription-PCR
Total RNA was isolated from lungs with TRIzol reagent (Invitrogen/Life Technologies, Carlsbad, CA) and cDNA was synthesized with a RevertAid first-strand cDNA synthesis kit (Thermo Scientific, Wilmington, DE) according to the manufacturer's instructions. mRNA expression of hHGF, mHGF, mouse c-Met (mc-Met), cytokines (TNF-α, ICAM-1, interferon [IFN]-γ, interleukin [IL]-6, IL-10, TGF-β), collagen type 1 α1 (Co1a1) and Col3a1, and sphingosine 1-phosphate (S1P) receptors (S1PR1, S1PR3, and S1PR5) was quantified with the StepOnePlus real-time PCR system (Applied Biosystems/Life Technologies, Foster City, CA) and SYBR Premix Ex Taq II (Perfect Real Time) (Takara Bio). The primers were as follows: hHGF, sense 5′-gccatgaatttgacctct-3′ and antisense 5′-gacatttgatgccactctta-3′; mHGF, sense 5′-acgatgctactggtatcctttcaa-3′ and antisense 5′-cgtccctttatagctgcctcc-3′; mc-Met, sense 5′-agaagttcaccaccaagtca-3′ and antisense 5′-aaggcgtctggacagtattc-3′; mTNF-α, sense 5′-cgtggaactggcagaaga-3′ and antisense 5′-acagaagagcgtggtggc-3′; mICAM-1, sense 5′-gacgcagaggaccttaacag-3′ and antisense 5′-ggcttcacacttcacagttac-3′; mIFN-γ, sense 5′-actaccttcttcagcaacagcaa-3′ and antisense 5′-ctggtggaccactcggatga-3′; mIL-6, sense 5′-gctacctggagtacatgaagaaca-3′ and antisense 5′-ggtccttagccactccttctg-3′; mIL-10, sense 5′-aataagagcaaggcagtggag-3′ and antisense 5′-tgtatgcttctatgcagttgatga-3′; mTGF-β, sense 5′-ggcggtgctcgctttgta-3′ and antisense 5′-ctcatagatggcgttgttgc-3′; mCol1a1, sense 5′-ctcagaggcgaaggcaacag-3′ and antisense 5′-gggaggtcttggtggttttgtat-3′; mCol3a1, sense 5′-atgtttggtttggagaatctat-3′ and antisense 5′-ttgcagtggtatgtaatgtt-3′; S1PR1, sense 5′-cgcaagaacatctccaag-3′ and antisense 5′-gcaggcaatgaagacactc-3′; S1PR3, sense 5′-tgagcgacacctgaccat-3′ and antisense 5′-agccagcacatcccaatc-3′; S1PR5, sense 5′-cttaggacgcctggaaacc-3′ and antisense 5′-gcacgcagaagagcacatag-3′; and mouse β-actin (mβ-actin), sense 5′-tttccagccttccttctt-3′ and antisense 5′-gtctttacggatgtcaacg-3′. Expression levels were normalized to mβ-actin.
Statistical analysis
Values are presented as means±SEM or as means±SD. One-way analysis of variance was used to compare the means of two or more experimental groups, followed by the Dunnett post hoc test. Differences between groups were considered to be significant at p<0.05.
Results
MSC-based HGF gene therapy attenuates radiation-induced histopathological changes
In vitro, Ad-HGF transduction increased the expression of hHGF in MSCs, but no obvious change was detected in their phenotype (Supplementary Fig. S1). Histological changes in lung sections obtained 28 and 180 days after treatment were assessed (Fig. 1A). Radiation induced marked inflammatory infiltrates, alveolar hemorrhage, alveolar wall rupture, and fibroblast proliferation on day 28, and interalveolar septal thickening was detected on day 180. Compared with the MSC-Null group, treatment with MSCs-HGF reduced airspace inflammation and alveolar hemorrhage. The thickness of interalveolar septa was modestly increased in the radiation group, with a slight reduction in the MSC-Null mice and a significant reduction in the MSC-HGF group.

Effect of mesenchymal stem cells (MSCs)-Null and MSCs-hepatocyte growth factor (HGF) treatment on radiation-induced lung injury.
Masson staining on day 28 showed alveolar, bronchial, and vascular collagen deposition, which was increased on day 180 (Fig. 1B). MSC-HGF treatment reduced collagen deposition compared with the MSC-Null group and the radiation group.
Effect of MSCs-Null and MSCs-HGF on radiation-induced lung permeability
Concentrations of total protein, albumin, and IgM were assayed in BALF to evaluate the integrity of the alveolar–capillary membrane barrier and to assess pulmonary vascular leakage. Total protein was markedly increased (4-fold) on day 28 after radiation (p<0.01 compared with the nonradiation group). Compared with the radiation group, total protein, albumin, and IgM were altered slightly after treatment with MSCs-Null, but the difference was not significant. However, treatment with MSCs-HGF reduced these lung injury indicators, and the total protein concentration was 1.6-fold greater than that in the nonradiation group (Fig. 1C).
Distribution and engraftment time of MSCs in radiated mouse model
To evaluate the homing efficiency of implanted MSCs for injured lung tissue, Ad-Luc infected MSCs were prepared and injected into radiated and nonradiated mice via the tail vein and their distribution was determined by the IVIS 50 imaging system (Xenogen, Alameda, CA). MSCs preferentially homed to radiation-injured lung tissue and their survival time was longer than in nonradiated lung (Fig. 2A).
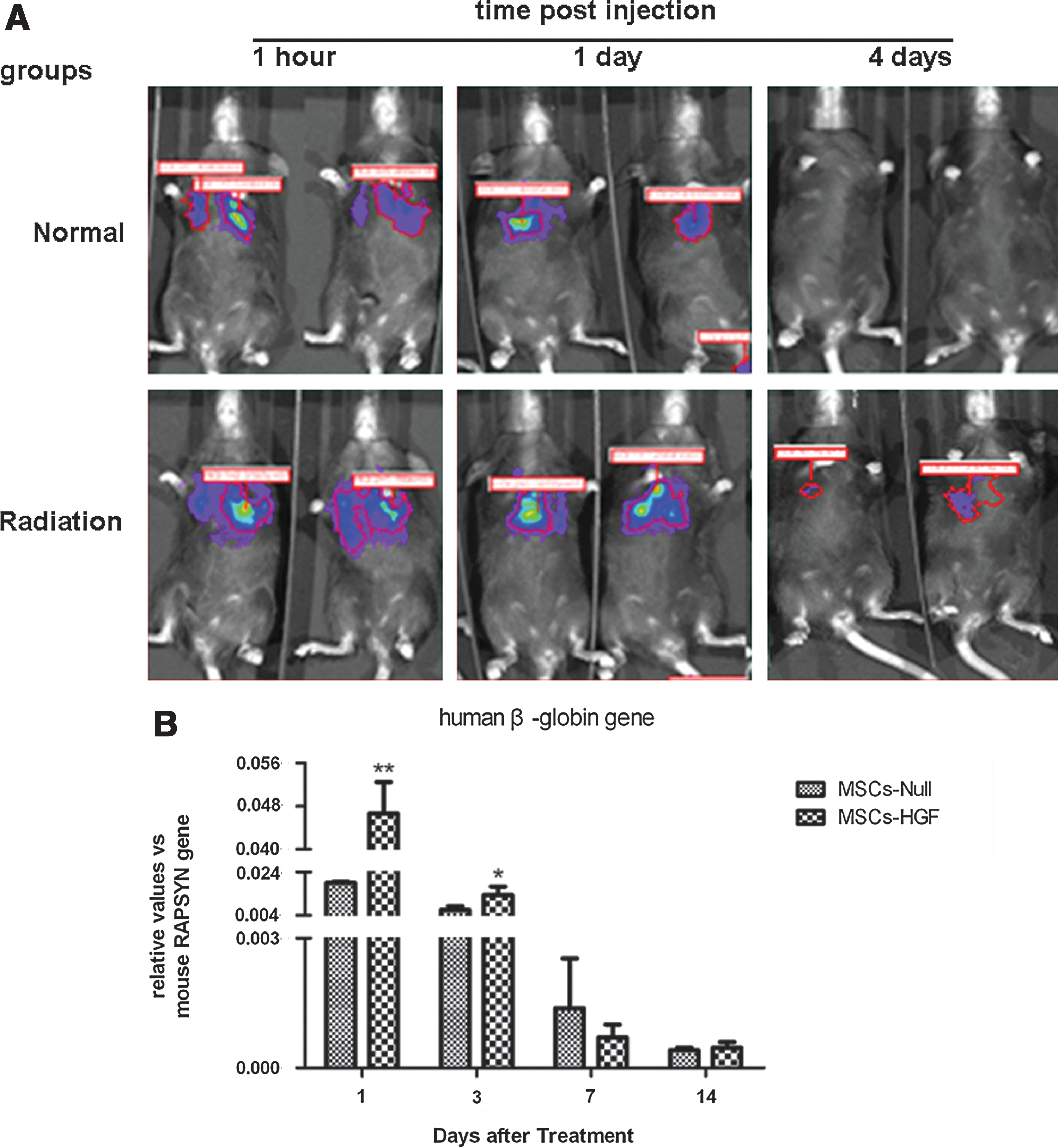
Retention of injected human mesenchymal stem cells (MSCs) in mice with and without radiation-induced lung injury (RILI). The distribution and engraftment time of human MSCs in an RILI mouse model were determined by the Xenogen in vivo optical imaging technique
We evaluated expression of the human β-globin gene in radiation-injured lung by quantitative PCR (qPCR) at various time points after MSC-based gene therapy. Levels of human β-globin gene expression were higher in the MSC-HGF group than in the MSC-Null group, and the difference was significantly different (p<0.05) (Fig. 2B).
MSC-HGF therapy mediates HGF expression in mouse lung after radiation
Our previous experiments showed that MSCs preferentially homed to radiation-injured lung tissue. Bone marrow-derived MSCs were then isolated, expanded, and infected with Ad-HGF or Ad-Null before being transfused into the radiated mouse model. We evaluated hHGF gene expression in the radiation-injured lung by qPCR at various time points after the MSC-based gene therapy. Gene expression was normalized to mβ-actin. hHGF expression was increased significantly in the MSC-HGF group compared with the MSC-Null and radiation groups, and the differences were significantly different (p<0.05) (Fig. 3A).

Mesenchymal stem cell (MSC)-hepatocyte growth factor (HGF) therapy mediated expression of human HGF (hHGF), mouse HGF (mHGF), and mouse c-Met receptor in lung. Total RNA was extracted from lung tissue on days 1, 3, 7, 14, and 24 after treatment. hHGF
MSC-HGF therapy induces expression of mHGF and its receptor c-Met
MSCs-HGF can induce the secretion not only of hHGF, but also other cytokines by radiation-injured lung tissue in vivo. Using qPCR, we found mHGF expression to be increased significantly after MSC-HGF treatment (Fig. 3B).
Biological responses to HGF are initiated by the interaction with its receptor c-Met. It has been reported that radiation can induce c-Met protein production. In the present study, c-Met protein was detected in the plasma membrane and cytoplasm of alveolar epithelial cells. Compared with the radiation group and the MSC-Null group, MSCs-HGF produced a significantly greater increase in the expression of c-Met (Fig. 3C and D).
MSC-HGF therapy attenuates secretion and expression of proinflammatory cytokines and improves the expression of antiinflammatory cytokines
To evaluate the antiinflammatory activity of MSC-HGF, TNF-α and soluble ICAM-1 production was measured by ELISA in peripheral blood on days 7, 14, and 28 after radiation (Fig. 4A and B). TNF-α levels increased steadily up to day 28. Treatment with MSCs-Null variably decreased TNF-α and soluble ICAM-1, whereas treatment with MSCs-HGF markedly reduced their secretion.
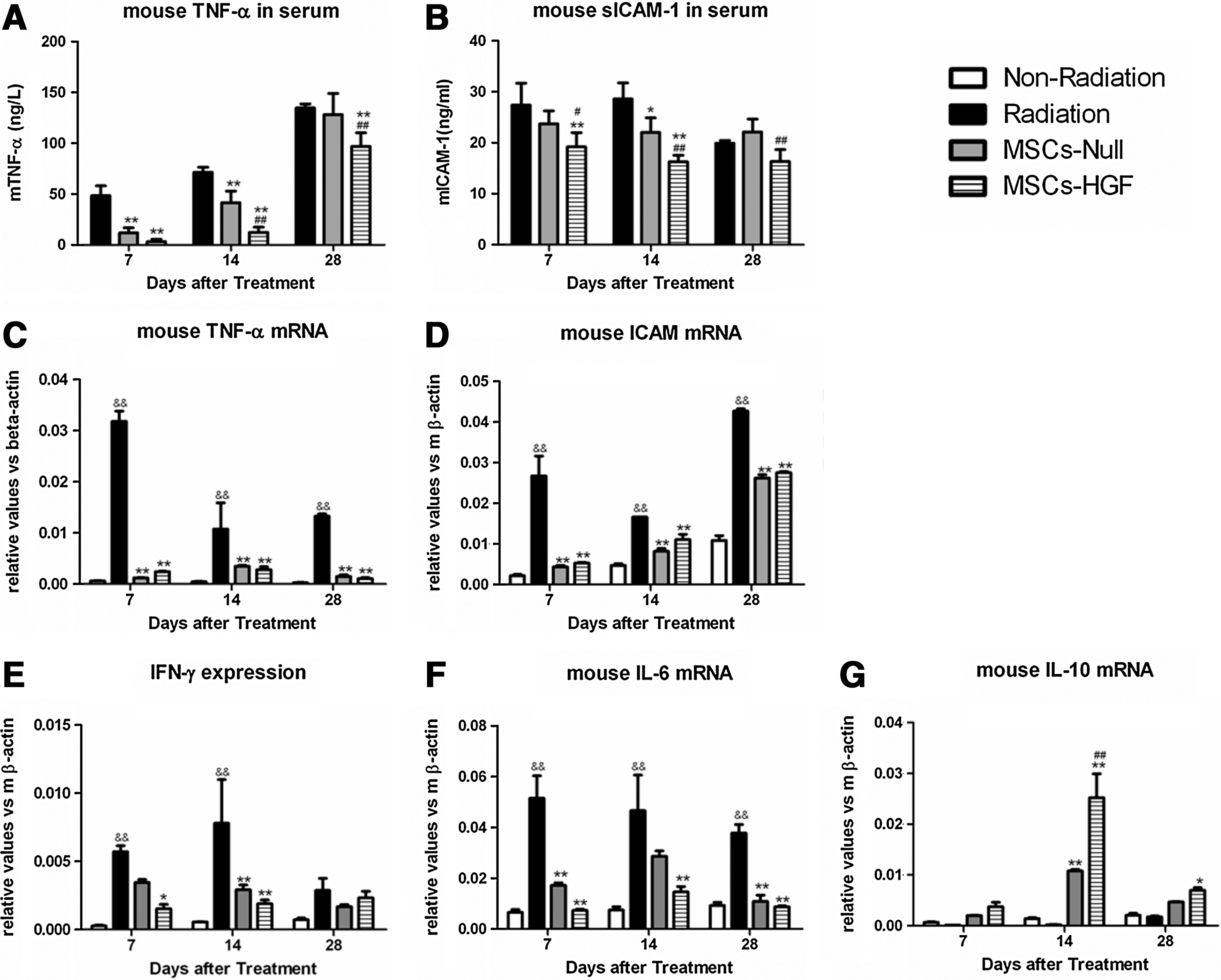
Expression levels of proinflammatory and antiinflammatory cytokines in peripheral blood and lung tissue. Peripheral blood was collected from the tail vein on days 7, 14, and 28 after mesenchymal stem cell (MSC)-based HGF gene therapy and expression levels of the proinflammatory cytokines tumor necrosis factor (TNF)-α
Real-time RT-PCR was used to detect the local inflammatory reaction of lung tissue on days 7, 14, and 28 after radiation (Fig. 4C and D). Compared with the radiation group, MSCs-Null and MSCs-HGF reduced the mRNA expression levels of TNF-α, ICAM-1, IL-6, and IFN-γ, and increased the expression of IL-10, and the difference was significantly different (p<0.01)
MSC-HGF therapy reduces expression of profibrotic factors TGF-β, Col1a1, and Col3a1
TGF-β plays an important role in the development of fibrosis after radiation of C57BL/6J mice. Its protein concentration in peripheral blood was measured by ELISA (Fig. 5A). On days 14 and 28 after treatment, the MSC-HGF group had reduced serum TGF-β levels compared with the radiation and MSC-Null groups. On days 7, 14, and 28 after treatment, MSCs-HGF had significantly reduced the mRNA expression levels of TGF-β, Col1a1, and Col3a1 in mouse lung tissue compared with the radiation and MSC-Null groups (Fig. 5B–D).
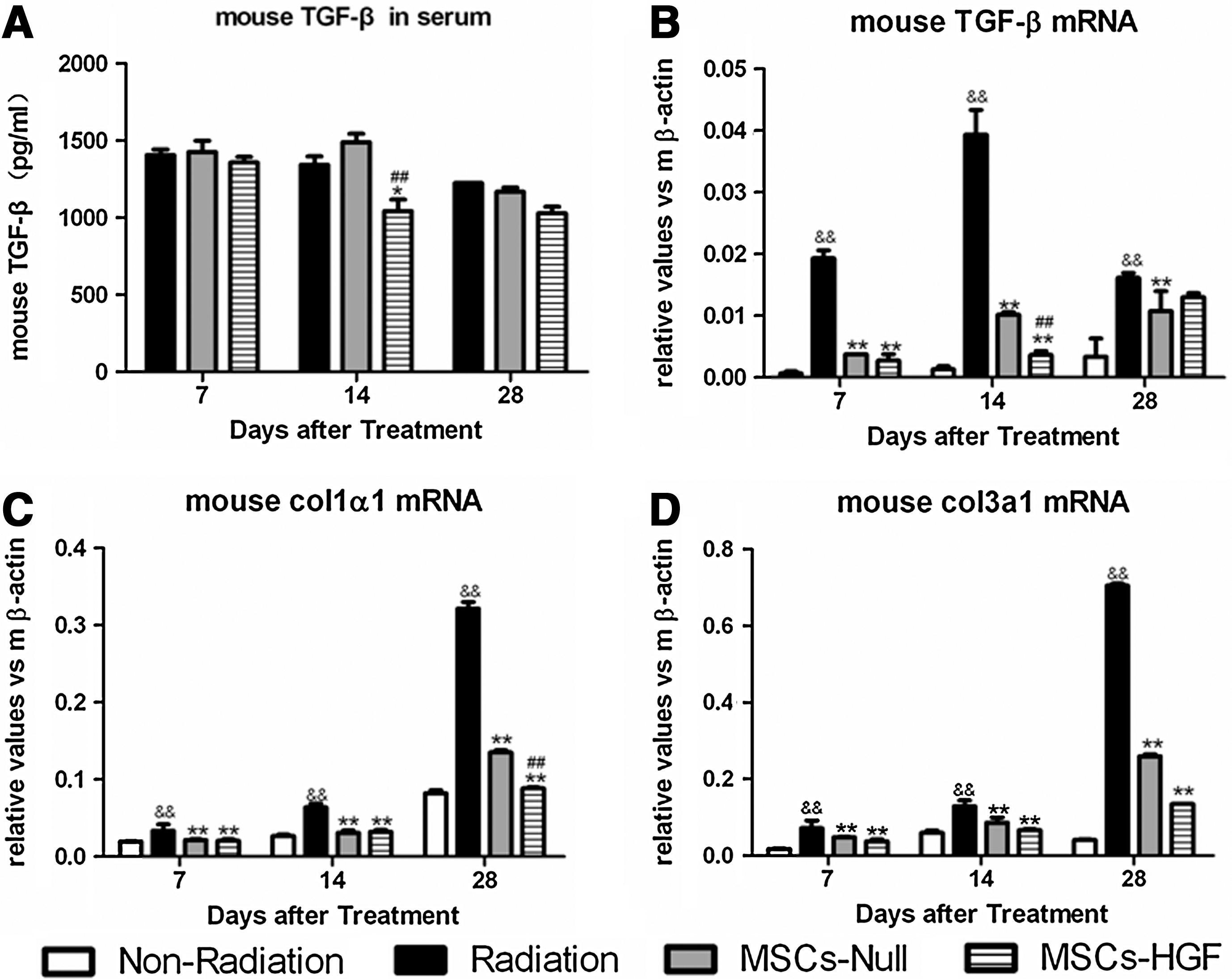
Expression levels of transforming growth factor (TGF)-β, Col1a1, and Col3a1 in serum and lung tissue after mesenchymal stem cell (MSC)-based hepatocyte growth factor (HGF) gene therapy. Peripheral blood was collected from the tail vein on days 7, 14, and 28 after MSC-based HGF gene therapy and levels of TGF-β were measured by ELISA
MSCs-HGF promote proliferation of lung epithelial cells and protect them from apoptosis
PCNA staining showed that a single tail vein injection of MSCs-HGF or MSCs-Null induced a significant increase in alveolar epithelial cell proliferation compared with the nonradiation and radiation groups (Fig. 6A and D). The maximal increase in PCNA labeling was observed on day 7 after MSCs-HGF injection; a slight decline was then detected.

Proliferation and apoptosis of lung cells after mesenchymal stem cell (MSC)-hepatocyte growth factor (HGF) therapy. Immunohistochemical staining of proliferating cell nuclear antigen
Because both cell proliferation and cell death can modulate alveolar repair, we also studied the percentage of apoptotic cells in radiation-injured lungs after treatment. Apoptotic cells were common within the thin alveolar septal walls and at intraalveolar sites after radiation. The rate of apoptosis reached a peak 7 days after radiation, but was significantly decreased in MSC-HGF and MSC-Null mice from day 1 to day 28. For example, on day 28 after radiation, apoptosis rates were 20.14±3.21% in the radiation group, 9.72±2.04% in the MSC-HGF group, and 13.39±3.04% in the MSC-Null group (Fig. 6B and E). Bcl-2 is one of the antiapoptotic members of the Bcl-2 family, and plays a key role in regulating apoptosis. The results showed that MSCs-Null or MSCs-HGF could improve the expression of Bcl-2 in lung on day 14 after treatment (Fig. 6C).
MSCs-HGF alter mRNA expression of S1PR1, S1PR3, and S1PR5
It has been demonstrated that HGF can stimulate the release of sphingosine kinase (SPK), leading to the production of S1P, which regulates diverse physiological and immunological processes by activating the G protein-coupled receptors S1PR1–5 to engage downstream pathways. To clarify the mechanism of MSC-based gene therapy for RILI, local mRNA expression of S1PR1, S1PR3, and S1PR5 was determined by qPCR (Fig. 7). Compared with the MSC-Null and radiation groups, MSCs-HGF increased S1PR1 mRNA expression on days 28 and 180 after radiation, and the difference was significant (p<0.01). Expression of S1PR3 and S1PR5 was not increased on either day 28 or day 180.
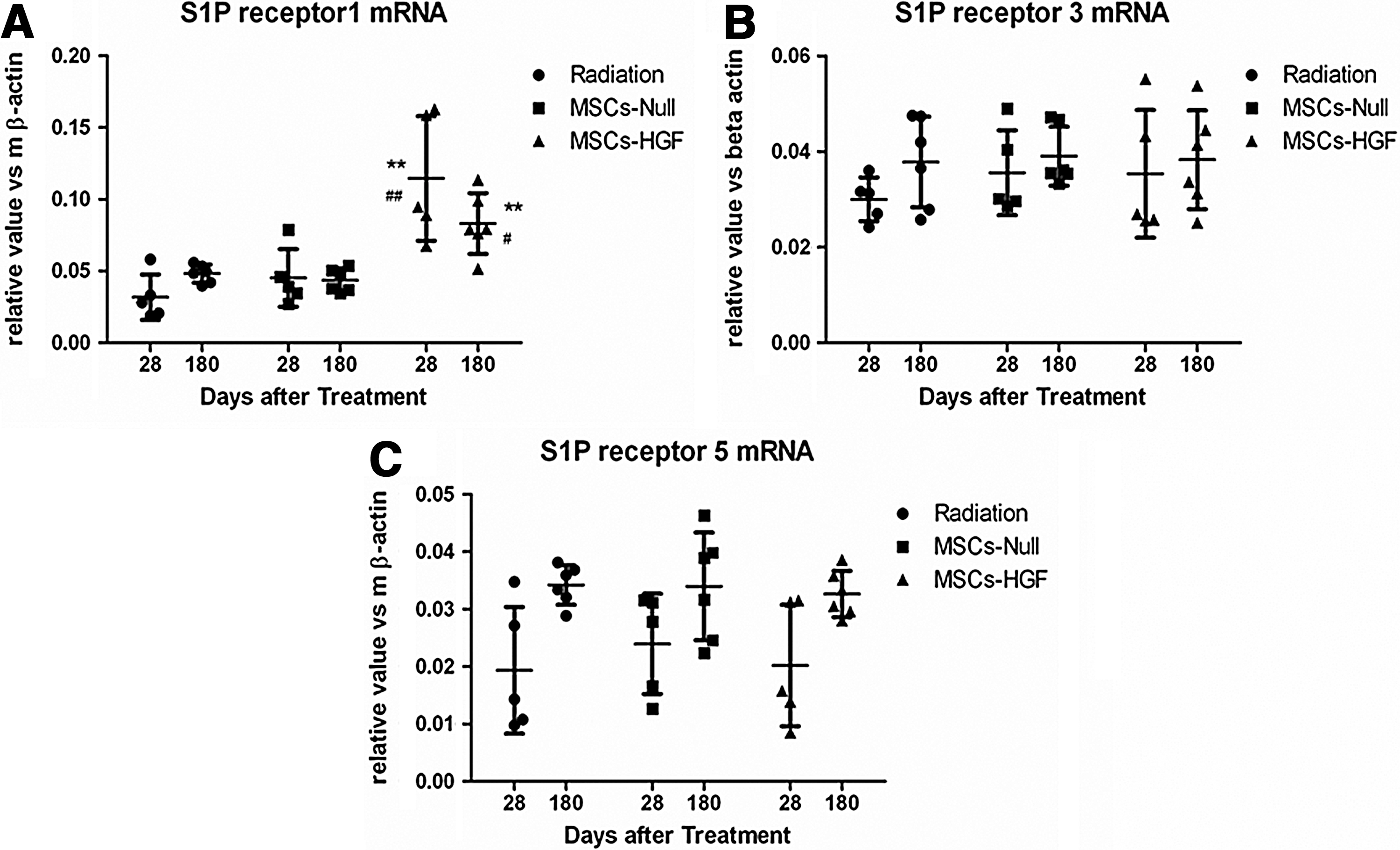
mRNA expression of sphingosine 1-phosphate receptor (S1PR) in lung tissue on days 28 and 180 after mesenchymal stem cell (MSC)-based hepatocyte growth factor (HGF) gene therapy. Expression of
Discussion
C57BL/6J (B6) mice respond to high-dose whole-thorax radiation by developing alveolitis and atelectatic regions of fibrosis at approximately 6 months after treatment (Sharplin et al., 1989; Haston et al., 2007). In this study, we established an RILI model in B6 mice and demonstrated that MSC-based HGF gene therapy attenuated both alveolar inflammation and permeability, and inhibited lung fibrosis.
MSCs are recognized as efficient gene delivery vehicles because of their low immunogenicity and their ability to accumulate at the site of tissue/organ damage, inflammation, or cancer (Mangi et al., 2003; Reiser et al., 2005). Studies have demonstrated that bone marrow-derived MSCs can engraft in injured lung and even differentiate into lung epithelial cells in vivo (Ortiz et al., 2003; Yamada et al., 2004; Denburg and van Eeden, 2006). In the present study, bone marrow-derived MSCs were transduced with adenovirus carrying the luciferase gene and these marked cells were injected intravenously into radiated B6 mice with injured lungs. The distribution of MSCs in normal and injured tissues was determined by the Xenogen in vivo optical imaging technique, which showed that MSCs preferentially homed to radiation-injured lung tissue (Fig. 2A). MSC-based gene therapy may thus have beneficial effects in the therapy of RILI. We found that HGF mRNA was expressed efficiently in lung tissue 28 days posttreatment, indicating that MSCs can deliver exogenous genes to local sites of injury.
MSCs are emerging as a therapeutic modality in various inflammatory diseases because of their antiinflammatory and immunomodulatory properties. It has been reported that MSCs can reduce the inflammatory response in the lung in both lipopolysaccharide- and bleomycin-induced acute lung injury models (Ortiz et al., 2003, 2007; Mei et al., 2007). We found that MSCs-Null decreased the expression of TNF-α and soluble ICAM-1 in peripheral blood on days 7 and 14 after radiation. However, bone marrow-derived stem cells may participate in the generation of fibrosis, correlating with the early inflammatory stages of the pathogenesis. In the bleomycin model, it has been demonstrated that MSCs must be delivered early to attenuate the resulting fibrosis (Ortiz et al., 2003). In the present study, MSCs were injected intravenously immediately after radiation to reduce early inflammation and to inhibit late fibrosis. However, no difference was found in TGF-β expression, which plays an important role in fibrosis, after MSC-Null treatment; the mechanism of this is not understood.
HGF is a multifunctional factor with actions including the stimulation of angiogenesis, the inhibition of fibrosis, and the reduction of apoptosis (Matsumoto and Nakamura, 1992, 1997). HGF is expressed locally in response to injury in numerous tissues, including the lung, kidney, and liver (Yamazaki et al., 1996). Although HGF expression is increased after lung injury, an inverse correlation has been identified for HGF expression during the development and/or progression of fibrosis. Here, we successfully developed an MSC-based HGF gene therapy using an adenoviral vector for the treatment of RILI. Expression of HGF was detected in lung tissue 28 days after treatment. We also found that the combination of MSCs and HGF gene therapy improved the histopathology, reduced inflammation, and inhibited fibrosis.
c-Met, a tyrosine kinase receptor of HGF, is expressed by epithelial and endothelial cells. In response to HGF binding, phosphorylation sites at critical tyrosine residues on c-Met associate with multiple adaptor proteins and signaling molecules (Ponzetto et al., 1994), and signal transduction leads to various biological responses including migration, proliferation, and morphogenesis (Rubin et al., 1993). It is generally accepted that endogenous HGF/c-Met signaling plays important roles in tissue repair. And their expression was upregulated by exogenous stimuli such as MSCs and gene-modified MSCs (Kearns-Jonker et al., 2012). Our data showed that MSC-based HGF gene therapy increases expression of endogenous HGF/c-Met in a mouse RILI model. MSC-based HGF gene therapy increased c-Met expression in both the plasma membrane and the cytoplasm of alveolar epithelial cells on days 28 and 180 after radiation. These results indicate that signaling pathways of the HGF/c-Met system might play an important role in the therapy of RILI.
It has been reported that HGF stimulates SPK release via the phosphoinositide 3-kinase and ERK/MAPK pathways, leading to S1P production. S1P (Duan et al., 2004), a potent bioactive lipid produced by SPK, activates S1P receptors and downstream signaling by autocrine and paracrine mechanisms (Takabe et al., 2008). In the present study, we found that MSCs-HGF increased expression of S1PR1 in the lung, whereas expression of S1PR3 and S1PR5 showed no significant change after treatment with MSCs-Null or MSCs-HGF. It has been demonstrated that S1P1 plays a key role in angiogenesis and vascular maturation, as well as in the regulation of immune cell trafficking, endothelial barrier function, and vascular tone (Matloubian et al., 2004; Sanna et al., 2006). Thus, S1P/S1PR1 may play an important role in HGF-mediated RILI therapy.
In conclusion, we have demonstrated that MSC-based HGF gene therapy after RILI could mediate HGF expression in the injured lung, improve lung function and histopathology, reduce inflammatory cytokines and profibrotic factors, and inhibit fibrosis. S1P/S1PR1 may participate in HGF/c-Met-mediated antiinflammatory and antifibrotic responses.
Footnotes
Acknowledgments
This work was supported by grants from the National High Technology Research and Development Program of China (863 Program, no. 2012AA020807), the National Basic Research and Development of China (973 Program, no. 2012CB518205), the National Natural Science Foundation of China (no. 81072240), and the International Atomic Energy Agency Program (no. 15309).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.