
Abstract
Electrotransfer of genes is one of the preferred strategies used to deliver plasmid DNA into skeletal muscle. In our experience, the combination of hyaluronidase (HYA) with electrotransfer (ET) of DNA vaccine enhances transfection of muscular fibers and increases expression of the encoded antigen. However, the contribution of HYA to the inflammatory reaction induced by ET, and its role in supporting ET adjuvancy, has never been investigated. We analyzed the events occurring in the first 2 weeks after electrotransfer to mouse muscle in the presence of HYA, to verify whether HYA contributes to the local inflammatory response induced by ET. Our results demonstrate that HYA amplifies the ET effect in terms of inflammatory cell recruitment enhancing the early release of interleukin (IL)-1β, tumor necrosis factor-α, and IL-6 cytokines. In contrast, HYA does not induce helper T cell type 1 and 2 cytokine production, confirming that the DNA vaccine is indispensable to induce mediators of antigen-specific immune responses. We observed inflammatory cell migration in the muscle treated with HYA plus ET in a time window between days 4 and 7 after cytokine induction. These observations are important in the choice of prime–boost intervals for optimizing ET-based DNA vaccination protocols. Because HYA contributes to vaccine spread and enhances the proinflammatory effect of ET in muscle we strongly support the use of HYA to potentiate DNA vaccine efficacy.
Introduction
Remarkable success in intramuscular DNA vaccination is obtained when vaccines are applied in the presence of an electrical field after plasmid DNA injection. This technology is widely known as DNA or gene electrotransfer (Mir et al., 1999; Khan et al., 2003; Prud'homme et al., 2006; Reed and Li, 2009; Chiarella et al., 2010; Sardesai and Weiner, 2011). Gene electrotransfer is a safe tool in DNA vaccination. In particular, proteomic studies have shown minor alterations in the gene expression profile as well as in the protein expression pattern (Hojman et al., 2007; Mlakar et al., 2009) with the exception of the stress proteins, the induction of which is a consequence of inflammatory events (Rubenstrunk et al., 2004). As we demonstrated previously, the application of electrotransfer (ET) on the skeletal muscle of a mouse is able to induce local production of inflammatory cytokines and recruitment of inflammatory cells to the treated site (Chiarella et al., 2008b). The mild stress condition induced by ET is responsible for the production of danger signals activating the innate immune reaction, indispensable for supporting the adaptive immunity response to the vaccine. Lymphocytes and inflammatory cell migration as well, were shown to be sensitive to the applied electrical fields according to a phenomenon called electrotaxis (Lin and Pullar, 2011). As a result, ET emerged not only as a delivery tool but also as an adjuvant for naked DNA vaccination (Gronevik et al., 2005b; Ahlen et al., 2007; Chiarella et al., 2008b).
Diffusion of plasmid DNA vaccines into skin and skeletal muscle was further improved by preinjection of hyaluronidase (HYA) (Molnar et al., 2004), an enzyme responsible for degradation of the extracellular matrix surrounding muscle fibers. The treatment of skeletal muscle with HYA and ET was a successful combination enhancing the number of transfected muscle fibers without increasing the muscle damage associated with ET (McMahon et al., 2001). The final effect was improved delivery of the foreign gene and a general enhancement of DNA vaccine efficacy (Signori et al., 2010). HYA enzyme is successfully used in applications other than vaccination. In mice it increases the efficiency of transfection by gene ET in solid subcutaneous tumors, and it has been shown to be effective for the spreading of chemotherapeutic agents in various cancer types (Spruss et al., 1995; Klocker et al., 1998; Cemazar et al., 2012).
The use of HYA in vaccination has an earlier origin. In the years 1940–1950 HYA was used as a “spreading factor” to allow better diffusion of traditional vaccines in the site of inoculation. In 1950 Bergqvist reported that the effect of bacillus Calmette-Guèrin (BCG) vaccine in guinea pigs was enhanced when HYA was added to the vaccine (Bergqvist, 1950) and Tozuka and colleagues published clinical results on the enhancement of the effect of BCG vaccination in the presence of HYA (Tozuka et al., 1954). Salvioli and colleagues as well demonstrated improved intradermal penetration of heat-killed microorganisms of the antituberculosis vaccine in infants (Salvioli et al., 1952). Although HYA was proven to increase the efficacy of vaccines when used with both traditional (Bergqvist, 1950; Salvioli et al., 1952; Tozuka et al., 1954) and plasmid DNA vaccines (McMahon et al., 2001; Vandermeulen et al., 2009; Potter and Heller, 2011), no one studied its role when used in combination with ET. As a consequence, we asked whether exogenous HYA is able to affect skeletal muscle and may influence the muscle inflammatory reaction when used alone or with ET. To fulfill this task we investigated the biological events occurring in muscle in the first 2 weeks after HYA treatment in the presence and absence of ET. In particular we analyzed (1) the immunogenic properties of exogenous HYA in muscle, (2) skeletal muscle morphology and the inflammatory cell phenotype observed after HYA inoculation with and without ET in a defined time interval, and (3) the cytokines of innate and adaptive immunity produced at the local level.
Our results indicate that HYA enhances ET-induced inflammation and that its use should be recommended in DNA vaccination protocols.
Materials and Methods
Animal treatment and reagents
Six-week-old male BALB/c mice were housed according to the guidelines for animal care of Università Cattolica del Sacro Cuore of Rome. Animals were checked for their weight in order to have homogeneity and then distributed into five groups according to the treatment they underwent. Each group consisted of at least 12 animals and the specific treatment was applied on both posterior hind limbs. Groups included the following: (1) negative control, (2) ET alone, (3) HYA plus ET, (4) HYA alone, and (5) lipopolysaccharide (LPS) (Sigma-Aldrich, St. Louis, MO). For each treatment group three animals were killed at the following time points: 3 hr, 4 days, 7 days, and 14 days. A total of six skeletal muscles and three spleens for each time point were harvested for morphological analysis, cytokine detection, and immunohistochemistry. Before electrotransfer mice were anesthetized with a mixture of a KETAMINE–medetomidine (Domitor) in the muscle of the anterior limb. The following conditions were used for ET treatment: an electric field of 175 V/cm was applied in 10 pulses of 20-msec square-wave pulses at 1 Hz, using an ECM 830 pulse generator provided of Tweezertrodes (Harvard Apparatus, BTX Instruments Division, Holliston, MA). Mice were shaved after anesthesia and electric fields were applied to the gastrocnemius muscle of both posterior limbs through forceps with terminal circular plates after conductive gel application. As positive control for endotoxin-induced inflammation mice were injected intramuscularly with 25 μg of LPS. Ten units of bovine HYA (H-4272; Sigma-Aldrich, Milan, Italy) were resuspended in 30 μl of 0.15 M sodium phosphate buffer and were intramuscularly injected into the muscles 2 hr before ET treatment.
Histology examination
Skeletal muscles and spleens were removed from mice belonging to each experimental group and were either frozen in dry ice and stored at −80°C or fixed with 4% buffered formalin for 24 hr. Some of the frozen organs were embedded in cryostat embedding medium (Bio-Optica, Milan, Italy) for cryosectioning, and the remaining spleens were used for protein extraction. Organs fixed with 4% formalin were dehydrated by an automated machine (Histo-Lab, Gothenburg, Sweden) and embedded in paraffin for histology analysis.
For the tissue morphology examination, paraffin sections (thickness, 12 μm) were cut with a microtome (Microm HM325; Thermo Fisher Scientific, Waltham, MA) and collected on SuperFrost Plus slides (Bio-Optica), and hydrated in a descending sequence of ethanol before staining with hematoxylin–eosin (Bio-Optica). Slides were dehydrated in an ascending ethanol series and mounted with Eukitt (O Kindler, Freiburg, Germany). Slides were air-dried overnight before observation under an optical microscope (Leitz DMR; Leica, Wetzlar, Germany). Images were acquired with NIS-Free software, using a Nikon digital sight DS-U1 camera (Nikon, Tokyo, Japan).
Immunohistochemistry
Tissue cryosections (20 μm) were cut with a cryostat (Leica), fixed for 10 min with chilled acetone, dried, and rinsed in phosphate-buffered saline (PBS) for 10 min. Endogenous peroxidase activity was quenched by incubating the slides for 15 min in 0.5% H2O2–PBS. After one wash with PBS the nonspecific binding sites of the tissues were saturated by incubating slides with 3% bovine serum albumin (BSA) in PBS (blocking buffer) for 30 min. Sections were incubated in a humid chamber for 1 hr at room temperature with the following primary antibodies used at 1:50 dilution in blocking buffer: anti-mouse F480 clone BM8, anti-mouse MHC class II clone M5/114.15.2, and anti-mouse CD11c, clone N418 (eBioscience, San Diego, CA). After three washes with PBS, sections were incubated with biotinylated anti-rat IgG (diluted 1:1000) and anti-hamster IgG (diluted 1:1000) (eBioscience) for 1 hr at room temperature. Slides were washed again as described previously and incubated with streptavidin–horseradish peroxidase (HRP) (BD Biosciences, San Jose, CA) for 30 min at room temperature. Slides were washed and developed for 3 min with diaminobenzidine (DAB) according to the instructions of the manufacturer (Sigma-Aldrich). The reaction was stopped with H2O and slides were counterstained with hematoxylin (Bio-Optica), washed again with H2O, dehydrated in an ascending ethanol scale, and mounted with Eukitt. Slides were observed with a Leica microscope. Images were acquired with a Nikon digital camera equipped with NIS-Free software.
Protein extraction
For cytokine extraction, mouse tissues were weighed and lysed in extraction buffer consisting of 0.1% Igepal (Sigma-Aldrich), PBS, and protease inhibitor cocktail (Sigma-Aldrich) diluted according to the manufacturer's instructions. A volume of 50 μl of buffer was used to lyse 10 mg of tissue. Organs were homogenized with an Ultra-Turrax T25 (Janke & Kunkel, IKA Labortechnik, Staufen, Germany) and kept 20 min on ice before centrifugation at 20,000×g for 10 min at 4°C. Supernatants were collected, aliquoted, and stored at −20°C. Protein concentration was determined by Bradford assay, and the absorbance of samples after addition of dye reagent was measured at 595 nm in a spectrophotometer (Eppendorf, Hamburg, Germany).
Sandwich ELISA for cytokine detection
Sandwich ELISAs for detection of interleukin (IL)-1β, tumor necrosis factor (TNF)-α, IL-6, interferon (IFN)-γ, IL-2, IL-4, and IL-10 cytokines were carried out according to the instructions of the kit manufacturers (Invitrogen/Life Technologies [Carlsbad, CA] and eBioscience). Briefly, plates were coated overnight at 4°C with capture antibody and blocked with blocking buffer (3% BSA in PBS or diluent buffer) for 1 hr at room temperature. After blocking, plates were incubated for 1 hr at room temperature or overnight at 4°C with 75 μg of protein extract per well diluted in blocking buffer. Duplicates of each sample were made in each assay. Plates were washed with wash buffer (PBS–0.05% Tween) (Sigma-Aldrich), incubated with biotinylated detection antibody for 1 hr at room temperature, and washed again with wash buffer before incubation for 30 min at room temperature with streptavidin conjugated to HRP. After a last wash with PBS, plates were developed with 3,3′,5,5′-tetramethylbenzidine (TMB; Unimed Scientific, Rome, Italy) substrate for 20–30 min. The reaction was stopped by adding 1 N H2SO4 to each well and the optical density was measured at 450 nm, using 620 nm as reference wavelength. In each assay recombinant cytokines were used as positive controls.
Measurement of mouse antibody titer versus HYA
Serum samples were collected from the tail vein of untreated mice, mice treated with HYA alone, mice treated with ET alone, and mice treated with HYA plus ET. Serum samples were harvested before immunization (preimmune serum) and 7 days after treatment and were used to perform the ELISA. Nunc MaxiSorp 96-well plates (Nunc, Roskilde, Denmark) were coated with bovine HYA (8 μg/ml) in 0.1 M NaHCO3 buffer, pH 9.6. Serum ELISA was performed as previously described (De Masi et al., 2005). Briefly, plates were rinsed in PBS and blocked with 3% BSA in PBS (blocking buffer) for 1 hr at room temperature. Mouse sera were incubated for 1 hr at room temperature at 1:100 and 1:500 dilutions in blocking buffer. After four washes of 5 min each with 0.05% Tween 20 in PBS (wash buffer) plates were incubated for 1 hr at room temperature with anti-mouse IgG antibody conjugated to alkaline phosphatase (Jackson ImmunoResearch, West Grove, PA) at 1:2500 dilution. Plates were then washed as described previously and developed for 50 min in the presence of p-nitrophenyl phosphate (PNPP) substrate prepared according to the instructions of the manufacturer (Sigma-Aldrich) After addition of 2 N NaOH to block the enzymatic reaction, the absorbance of the plate was read at 450 nm with an ELISA reader (Tecan, Grödig, Austria).
Statistical analysis
Analysis of variance (ANOVA) was done to compare differences in protein expression determined by ELISA in the four study groups. The differences that were likely to exist between groups were analyzed by unpaired t test. Statistical analysis was done with GraphPad software (GraphPad Software, San Diego, CA), where a two-tailed p value ≤0.05 was considered significant.
Results
Experimental design
For each experiment mice were distributed into five treatment groups, with each group consisting of 12 animals. Animals undergoing a specific treatment were killed at various time points (3 hr, 4 days, 7 days, and 14 days after treatment). To detect differences between early, intermediate, and late time points, antigen-presenting cell recruitment was evaluated at all the time points (3 hr, 4 days, 7 days, and 14 days). Analysis of skeletal muscle morphology was carried out 3 hr, 4 days, and 7 days after treatment. Proinflammatory and helper T cell type 1 (Th1)/Th2 cytokine release were measured at 3 hr, 4 days, and 14 days.
Comparative analysis of skeletal muscle morphology in the early phase after HYA pretreatment
Application of electrical field pulses to mouse skeletal muscle results in slight tissue damage and recruitment of infiltrating monocytes and granulocytes, which create a proinflammatory context useful in local antigen presentation if a DNA vaccine is coinjected. As the initial step we examined skeletal muscle morphology microscopically in the absence of plasmid DNA vaccine and looked for the presence of infiltrating leukocytes in the first 7 days after the following treatments: electrotransfer (ET), HYA injection (HYA), and electrotransfer combined with HYA injection (ET plus HYA). These observations were compared with those of skeletal muscles taken at the same times (3 hr, 4 days, and 7 days) after LPS injection. As shown in some representative images (Fig. 1), 3 hr after treatment none of the applied chemical or electrical stimuli (LPS, HYA, or ET) seemed to induce strong alteration of muscle morphology. Four days after ET, infiltrating inflammatory cells appeared numerous in the muscle and their localization in the interstitial space was enhanced when HYA was injected before ET. A substantial population of these cells was represented by monocytes/macrophages, which were attracted by the injury induced by electrical field pulses, as evident in muscle sections taken on day 7 after ET, and would probably take part in muscle regeneration (Arnold et al., 2007) (Fig. 1). After observing numerous skeletal muscle sections, we confirmed that ET alone is a potent inflammatory stimulus for skeletal muscle and that local cell infiltration was not an immediate event but occurred 4 days after ET application. Although HYA pretreatment before ET favored the spreading of infiltrating leukocytes into the interstitial areas of muscle, it did not affect significantly the amount of recruited cells in comparison with ET alone. The infiltrating cells were enrolled mainly by the inflammation process induced by ET and were associated with skeletal muscle necrosis of the injured fibers. As visible in Fig. 1, centrally nucleated fibers appeared on day 7, indicating myogenic cells proliferation and differentiation. We point out that although the morphology of treated skeletal muscle on day 14 is not shown in Fig. 1, it can be inferred in Fig. 2A–C where, in addition to the immunolocalization of antigen-presenting cell (APC) surface markers, the morphology of tissue is also clearly shown at all the time points considered.
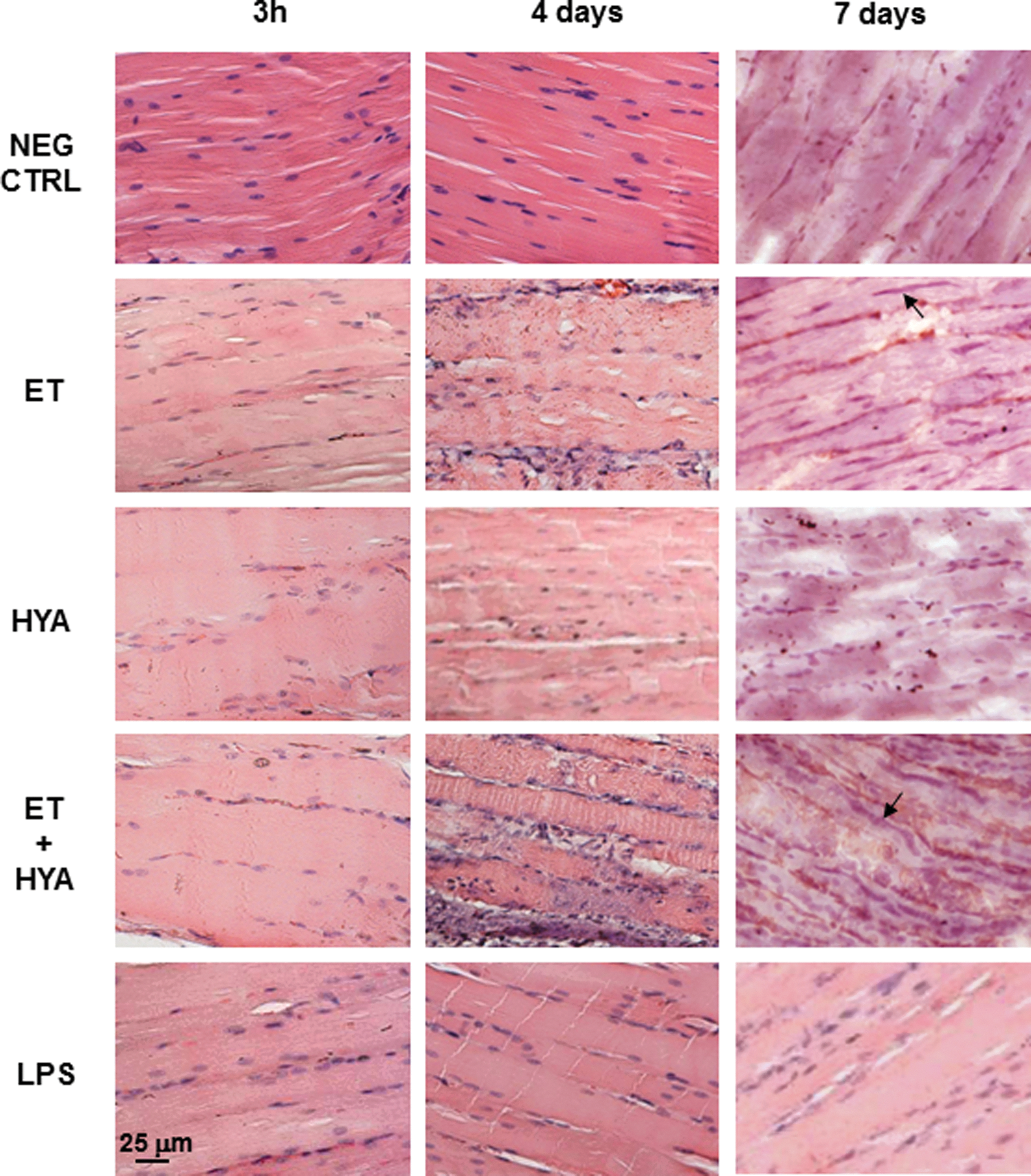
Histological analysis of skeletal muscle morphology and regeneration. Paraffin sections of skeletal muscles taken 3 hr, 4 days, and 7 days after various treatments as shown. Arrows indicate nuclei of regenerating muscle fibers. All images were taken at an original magnification of ×20 by optical microscopy. NEG CTRL, negative control; ET, electrotransfer; HYA, hyaluronidase injection; ET+HYA, electrotransfer after hyaluronidase injection; LPS, lipopolysaccharide injection. Color images available online at

Immunohistochemistry of muscles treated with HYA in the presence and absence of ET. Skeletal muscle cryosections were stained at various times after treatment (3 hr, 4 days, 7 days, and 14 days) with monoclonal antibodies specific for macrophages (


Analysis of local inflammatory cell recruitment
Having determined that infiltrating cells appeared 4 days after ET, we characterized the cell phenotypes and monitored their migration to the site of treatment in a time interval between 3 hr and 14 days. The cells were analyzed for the following surface markers: macrophages (F4/80-positive cells) and APCs, that is, MHC-II- and CD11c-positive cells, which include mature macrophages and dendritic cells. In Fig. 2 we show how ET treatment was able to recruit a conspicuous population of macrophages (Fig. 2A) and specialized APCs (MHC-II- and CD11c-positive cells) between days 4 and 7 (Fig. 2B and C). Intramuscular injection of HYA alone, in the absence of ET, did not recruit significant numbers of cells positive for the three surface markers. When the two treatments were combined (ET plus HYA), a peak in the number of F4/80-, MHC-II-, and CD11c-positive cells appeared on about day 4, similar to the LPS treatment (Fig. 2A–C). Therefore macrophage and APC migration to the inflammation site was not an early event but it took 4 days to occur. HYA preinjection amplified the effect of ET by recruiting a higher number of F4/80- and MHC-II-positive cells in comparison with ET alone at both day 4 and day 7 (Fig. 2A and B). No specific staining for the three surface markers was observed on day 14. Skeletal muscles treated with ET, alone or in the presence of HYA, clearly showed fiber regeneration as evident by the nuclei located centrally within the fibers (day 14) (Fig. 2A–C, right).
Antibody production after hyaluronidase injection
To understand whether bovine HYA per se had some antigenic properties when injected intramuscularly, we measured antibody production in the serum of mice 7 days after treatment with the enzyme.
Day 7 was preferred to day 14 to measure the antibody level against HYA because, due to the high number of APCs observed in the treated muscles in the time interval between days 4 and 7, we assumed an increase in presentation of HYA by the migrated APCs to the resident lymphocytes.
We point out that, in order to see differences in the colorimetric assay, the ELISA plate was developed for a longer time than usual. Mice injected with HYA alone responded with the production of specific IgG antibodies. This immune response was statistically significant with respect to the negative control (absence of HYA) (Fig. 3). In electrotransferred HYA-treated mice, the antibody titer did not change significantly, although it was slightly lower than in mice injected solely with HYA. Because ET was usually applied to skeletal muscle 1–2 hr after HYA injection we conclude that the enzyme was not modified by the electrical stimulation, maintaining unaltered its function. We conclude ET does not increase the physiological antibody response to HYA, 7 days after injection of the enzyme.
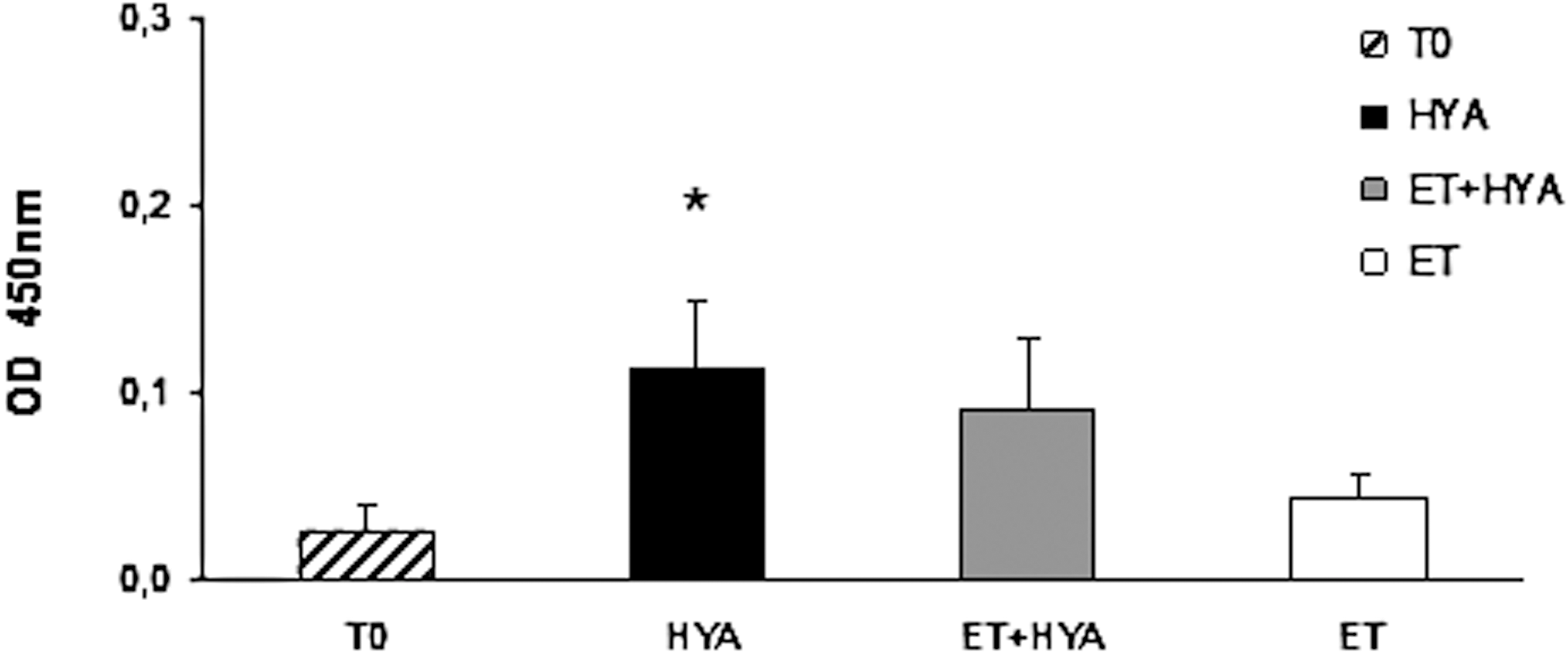
Evaluation of antibody response against HYA. ELISA was performed with mouse sera (1:100 dilution) harvested 7 days after HYA injection in the presence or absence of ET. HYA versus T0, p=0.0161 (*); HYA versus ET, p=0.0315; HYA versus HYA plus ET, p=0.5017. T0, negative control; ET, electrotransfer; HYA, hyaluronidase injection; ET+HYA, electrotransfer after hyaluronidase injection.
Evaluation of inflammatory and Th1/Th2 cytokine release induced by ET and HYA
We have already assessed that ET per se causes early production of proinflammatory cytokines such as TNF-α and IL-1β (Chiarella et al., 2008b). However, up to now we have never analyzed cytokine production in a late phase after ET application, nor did we know whether the combination of HYA with ET could have any specific effect on the process. Because hyaluronic acid (HA) is a product of degrading HYA activity and was shown to play a role in inflammation, tissue injury, and repair (Slevin et al., 2002), we wondered whether this combined treatment on skeletal muscle could influence the profile of cytokines involved in this phenomenon. We performed the analysis on two types of cytokines: TNF-α, IL-1β, and IL-6, generally characteristic of acute inflammations; and IL-2, IFN-γ, IL-4, and IL-10 which, respectively, define Th1- and Th2-mediated immune responses. The measurements of cytokine levels were carried out 3 hr, 4 days, and 14 days after treatment. In particular, 3 hr was chosen as an early time point because we were aware that production of cytokines related to an inflammatory event is usually an early phenomenon occurring within the first 8 hr after application of the inflammatory stimulus (Frost et al., 2002; Zhou et al., 2003; Chiarella et al., 2008b). Because inflammatory cytokines affected the migration of APCs and a peak in cell migration was observed mainly on day 4 and maintained until day 7, we assumed day 4 was the best intermediate time point for detecting cytokine release. To exclude any chance of a late release we selected 14 days as the latest time point to perform measurements.
Despite TNF-α levels appearing generally low, cytokine release in skeletal muscle was confirmed as an early phenomenon. Maximal cytokine release was observed 3 hr after ET, and the absence or presence of HYA did not seem to have an effect on their production (Fig. 4A). No significant difference in cytokine production was observed in the spleen, indicating that this organ is not affected by any treatment (Fig. 4A, bottom).
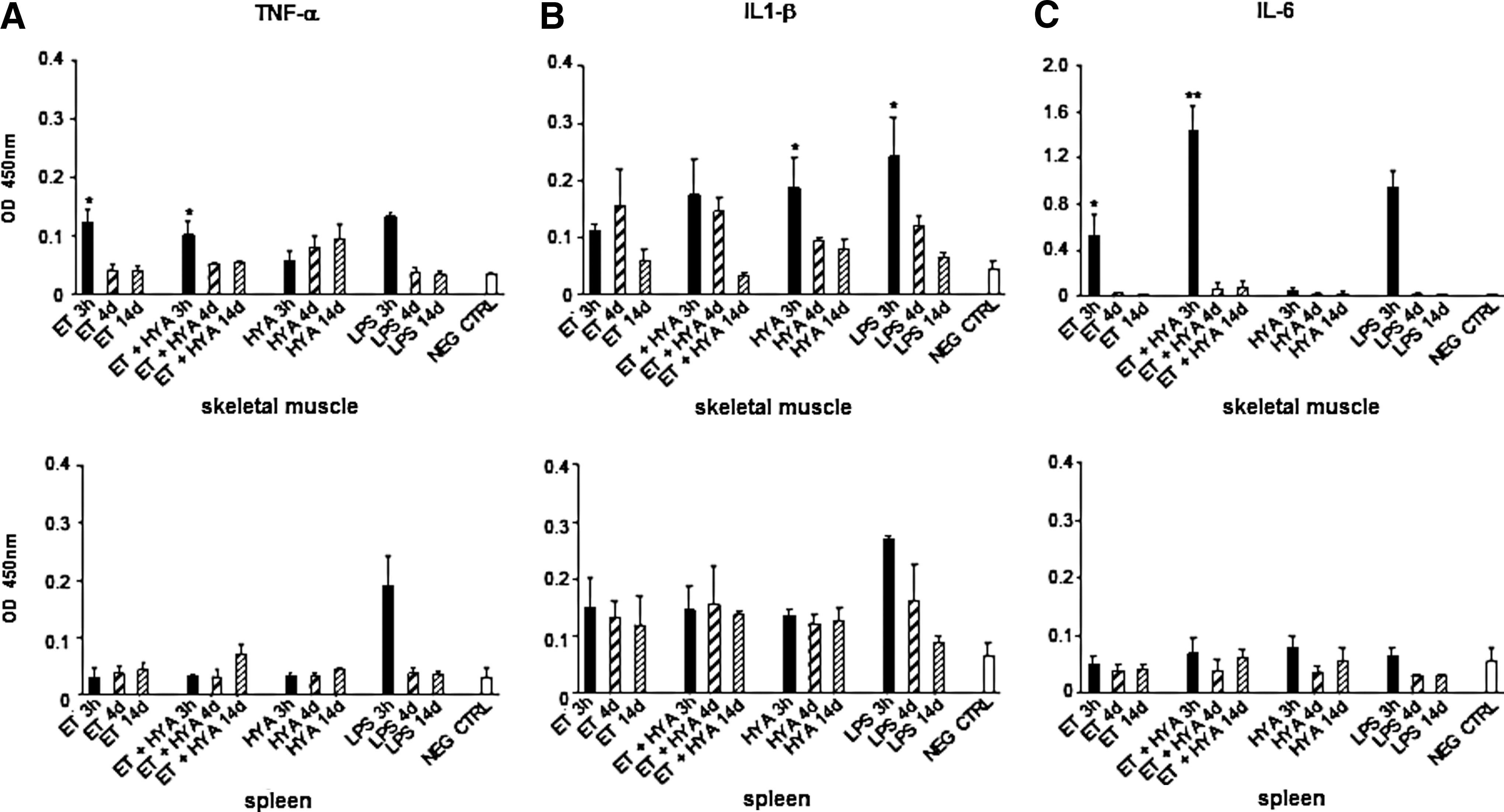
Measurement of TNF-α, IL-1β, and IL-6 release in skeletal muscle and spleen of treated mice. Differences in cytokine levels were estimated by comparing the optical density values (450 nm) measured by sandwich ELISA.
Similarly, IL-1β release was immediate, and the injection of HYA amplified the ET effect as shown in the muscles undergoing the combined treatment. Surprisingly, HYA alone induced an increase that was statistically significant in comparison with the negative control (untreated mouse) (Fig. 4B). We point out that cytokine production was not exhausted in the first hours; secretion was maintained until day 4. In particular, the combinatorial use of HYA and ET was responsible for the unexpectedly high levels of IL-1β detected on day 4, but we confirmed a further decrease in this cytokine at all the following time points. No significant release of IL-1β was observed in mouse spleen (Fig. 4B, bottom). Similar to TNF-α and IL-1β, we detected an early rise in IL-6 production 3 hr after skeletal muscles were treated with ET and HYA whereas no cytokine production was observed when HYA was injected in the absence of ET. The combined treatment (ET plus HYA) was a potent stimulus for IL-6 release (Fig. 4C). In the spleen IL-6 levels were at the limit of detection of the assay (Fig. 4C, bottom).
Th1 cytokines (IL-2 and IFN-γ) and Th2 cytokines (IL-4 and IL-10) were measured at the same time points in both muscle and spleen of treated and untreated mice and no appreciable amount of any of these molecules was revealed (Figs. 5 and 6).
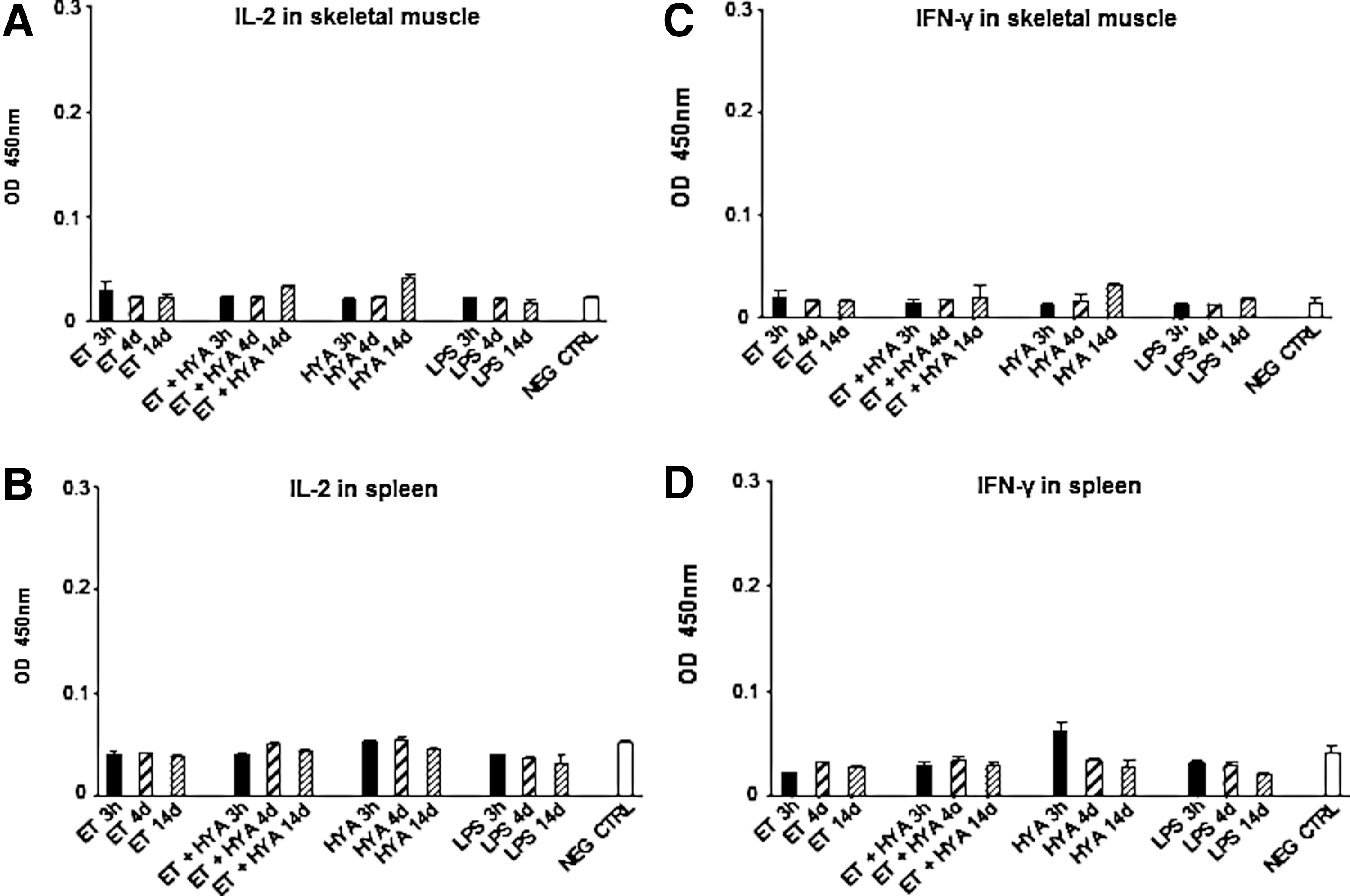
Measurement of Th1 cytokine levels in skeletal muscle and spleen of treated mice.

Measurement of Th2 (IL-4 and IL-10) cytokine levels in skeletal muscle and spleen of treated mice.
Discussion
Plasmid DNA vaccines, in contrast to traditional vaccines, are characterized by low immunogenicity when administered in the target site as unformulated injections. Their weakness is due mainly to the lack of adjuvant components, which are usually present in conventional vaccines and are responsible for innate immunity stimulation. Several strategies have been developed to increase DNA vaccination potency. Some are based on the introduction into the DNA backbone of immune modulator genes encoding cytokines, chemokines, helper T cell epitopes, or molecules targeting innate immunity, such as agonists of pattern recognition receptors (PRRs) (Hutnick et al., 2011); other strategies rely on DNA complexation within nanoparticles or polymers that mimic the particulate nature of conventional vaccines (Azad and Rojanasakul, 2006; Singh et al., 2006; Chiarella et al., 2007, 2008a; Fraser et al., 2007).
One of the most investigated strategies is based on the exploitation of chemical or physical devices aimed at facilitating DNA delivery into target organs. In particular, skeletal muscle electrotransfer has emerged as a safe and effective technology, leading to increased uptake of plasmid DNA and enhancement of the immune response because of activation of proinflammatory pathways and recruitment of inflammatory cells (Khan et al., 2005; Chiarella et al., 2008b; Hirao et al., 2008).
Our group and others have already demonstrated that ET is a delivery system with adjuvant effect on the skeletal muscle of animals immunized with naked DNA vaccines. ET creates local inflammation, promoting recruitment of monocytes/macrophages, polymorphonuclear leukocytes, and APCs favorable for the immune response to the vaccine (Durieux et al., 2004; Gronevik et al., 2005a; Ahlen et al., 2007; Chiarella et al., 2008b; Liu et al., 2008).
In terms of delivery efficacy, skeletal muscle ET is highly potentiated by preinjection of HYA, an enzyme hydrolyzing hyaluronic acid, an ubiquitous component of the extracellular matrix, resulting in increased gene expression, a higher number of transfected fibers, with an acceptable degree of tissue damage (McMahon et al., 2001; Gollins et al., 2003; Molnar et al., 2004; Evans et al., 2008). Despite the fact that several research groups have employed the combined use of HYA and ET in vaccination, no one has investigated the effect of HYA injection on skeletal muscle at the cellular and biochemical levels or determined whether this treatment influenced the inflammatory reaction caused by ET.
Here we defined the cellular events and chemical mediators involved in the first 2 weeks after ET treatment with and without HYA and in absence of DNA, comparing at various time points the skeletal muscles of electrotransferred mice. The results of the histological analysis of skeletal muscle sections indicate that infiltration of inflammatory cells into the extracellular space of muscle was induced by ET and enhanced by HYA. The enzyme amplified the effect of ET, favoring migration of a higher number of polymorphonuclear cells and APCs in comparison with application of ET alone. In the absence of ET, HYA was not able to develop the proinflammatory context necessary for the recruitment of inflammatory cells.
As proved by our data, inflammatory cell infiltration induced by ET, in the presence and in the absence of HYA, was not early but rather an intermediate event. It took 4 days to occur after treatment and, as previously shown (Chiarella et al., 2008b), the cells recruited to the muscle site were mostly F4/80 macrophages and dendritic cells with the MHC-II and CD11c phenotype. The only difference observed between ET plus HYA and ET stands in the earliest migration (day 4) of macrophages and MHC-II-positive cells induced by the combination of ET plus HYA compared with ET alone.
From this analysis we identified two time windows characterizing the kinetics of inflammatory cell migration in skeletal muscle treated with ET and ET plus HYA. In the first time window (days 4–7) we observed the highest number of F4/80-, MHC-II-, and CD11c-positive cells into the damaged muscle. In the second time window (days 7–14) we observed a substantial decrease in the number of these cell phenotypes in favor of muscle regeneration. These data are supported by other experimental observations (Arnold et al., 2007) and are important for the optimization of DNA vaccination protocols performed with ET and HYA. We conclude that at least 14 days is the appropriate interval for DNA vaccine administration into skeletal muscle because this time gap is sufficient to create the proinflammatory environment necessary for APC recruitment and uptake of DNA vaccine. On the other hand, the regenerating process of skeletal muscle starting in the second week after treatment may also favor DNA vaccine uptake by proliferating skeletal muscle cells (Peng et al., 2005).
When we looked at the biochemical mediators released in electrotransferred muscles, we observed that proinflammatory cytokines TNF-α, IL-1β, and IL-6 were released early, within the first 3 hr of treatment, confirming ET as being the primum movens of cytokine production.
Skeletal muscle TNF-α release was not affected by HYA alone whereas the early production of TNF-α, IL-1β, and IL-6 was influenced by the combination of ET plus HYA. In particular, HYA and ET were separately responsible for the early increase in IL-1β whereas the unexpectedly high level observed on day 4 seemed to depend on the combination of the two agents. Nevertheless, immediate IL-6 release participated in the local inflammatory reaction although we have at present no evidence indicating whether this cytokine was produced independently or in response to IL-1β and TNF-α (Akira et al., 1993).
Taken as a whole, these data indicate that HYA is a good enhancer of ET adjuvant because (1) it is not highly immunogenic; (2) it does not negatively affect skeletal muscle morphology; and (3) it favors APC infiltration and increases the release of proinflammatory cytokines in combination with ET.
In the light of these data, we strongly support the use of HYA to potentiate DNA vaccine efficacy, and we recommend at least 14 days as the optimal boosting interval to consider in DNA electrotransfer protocols in the field of veterinary and human medicine.
Footnotes
Acknowledgments
The authors are grateful to the following scientists of the University Campus Bio-Medico of Rome for providing access to their laboratory instruments: Sergio Morini, Simone Carotti, Flavio Keller, Ramona Marino, Alessandra Micera, and Bijorn Omar Balzamino. The authors thank the staff of the Animal Facility at the Sacro Cuore Catholic University of Rome for assistance in animal care. The authors thank Dr. G. Contini for critical revision of the manuscript. This work was supported by Energy for Research, a sponsor group of the Laboratory of Molecular Pathology and Experimental Oncology.
Author Disclosure Statement
The authors state no conflict of interest.