
Abstract
Heart diseases are major causes of morbidity and mortality in Western society. Gene therapy approaches are becoming promising therapeutic modalities to improve underlying molecular processes affecting failing cardiomyocytes. Numerous cardiac clinical gene therapy trials have yet to demonstrate strong positive results and advantages over current pharmacotherapy. The success of gene therapy depends largely on the creation of a reliable and efficient delivery method. The establishment of such a system is determined by its ability to overcome the existing biological barriers, including cellular uptake and intracellular trafficking as well as modulation of cellular permeability. In this article, we describe a variety of physical and mechanical methods, based on the transient disruption of the cell membrane, which are applied in nonviral gene transfer. In addition, we focus on the use of different physiological techniques and devices and pharmacological agents to enhance endothelial permeability. Development of these methods will undoubtedly help solve major problems facing gene therapy.
Every important research success brings new questions.
Introduction
Despite significant potential, the key rate-limiting step toward advancing cardiac gene therapies to the clinic is the establishment of safe and efficient delivery systems. In fact, most development efforts do not account for or address this critical need in early preclinical studies due to the much less complex scale of small animal models. Recently, however, there have been key advances in direct (e.g., injection at desired site) and transvascular (e.g., via arterio-venous system) gene delivery methods that are applicable to larger organisms in a clinical context. It should be noted that a wide variety of techniques have already been designed and applied for cardiac gene delivery in the clinical setting. However, despite the advancement of sophisticated minimally invasive percutaneous systems, inefficient delivery to molecular targets and subsequently inefficient gene transfer remains the outstanding problem. Ultimately, a viable cardiac gene therapy platform must have the following attributes: (1) the ability to transfer the therapeutic gene to all regions of the heart; (2) allow for the successful transduction of the targeted percentage of myocytes; (3) the appropriate level of genome copies within the transfected cardiomyocytes; and (4) establish a predictable relationship between quantifiable gene transfer and efficacy levels while determining if regional or global gene expression is sufficient to treat the selected disease.
In this article, we review (i) existing biological barriers for myocardial gene transfer, comparing direct and transvascular gene delivery strategies; (ii) cellular uptake and intracellular trafficking of viral versus nonviral vectors; (iii) engineered physical and mechanical methods for the enhancement of the cell's permeability kinetics; (iv) the most commonly used physiological and pharmacological techniques for regulation of gene expression and modulation of endothelial permeability; and (v) the relative success of these strategies in improving the efficiency of vector-mediated cardiac gene delivery.
Myocardial Gene Transfer and Existing Biological Barriers
Development of effective gene delivery systems capable of transfecting a wide variety of somatic tissues has become crucial for further progress in gene therapy. The available methods of cardiac gene delivery are typically classified by the site of injection, interventional approach, and the variation of cardiac circulation during transfer (Katz et al., 2011). Many clinical trials in cardiology and cardiac surgery have involved injecting the virus particle directly (intramyocardially) into the intended myocardial region. From an application point of view, this procedure is attractive due to its simplicity and lack of safety concerns. Furthermore, this approach has the ability to maximize concentration of vector in the target tissue and reduce transfer across the endothelial barrier, minimizing systemic spillover (Rapti et al., 2011). However, the transgene expression is limited to the needle track and cannot reach the nuclear envelope because the extracellular matrix restrains the spread of genome particles.
Localized intramyocardial injection of naked DNA to the left ventricle was first demonstrated in 1990 (Lin et al., 1990). The absolute amount of recombinant protein produced by plasmid injection was small given the limited distribution of transduced myocytes. The first use of replication-deficient adenovirus intramyocardially resulted in a 140,000-fold increase in the ratio of recombinant protein produced to the number of genomes injected compared to the injection of plasmid DNA (French et al., 1994).
In this respect, transvascular delivery initially seems more preferable. But virus infectivity is significantly inactivated in the bloodstream by neutralizing antibodies, plasma proteins and blood cells (Müller et al., 2007) (Fig. 1). It was demonstrated that over 90% of the virus dose first binds to erythrocytes, and about 98% of viral DNA in the blood is cell-associated, significantly decreasing viral access to the target tissue (Lyons et al., 2006).

Schematic representation of existing barriers during intramyocardial and transvascular gene delivery.
The basic principles in breaching the cellular biological barriers for different methods of gene delivery have been formulated by Escoffre and colleagues as follows: (i) transgene protection from destruction by nucleases located in the extracellular matrix; (ii) increasing the gene diffusion through the matrix; (iii) improving gene transport across the plasma membrane of target cells; (iv) enhancing intracellular gene migration; and (v) support in penetrating across the nuclear envelope (Escoffre et al., 2010).
Virus-mediated transvascular gene transfer generally must overcome three pathways: (1) blood cells and the capillary layer; (2) the vascular endothelial lining and the extracellular matrix; (3) cell membrane and the nucleus (Sasano et al., 2007). Different gene delivery strategies have their own barriers that must be bypassed (Melo et al., 2004).
Cell membrane permeabilization
Most current gene transfer strategies aim at enhancing transgenic expression, which in turn is determined by a number of exogenous and endogenous factors (Quarck and Holvoet, 2004). One of the most important endogenous stimuli in the transgene expression regulation is cell membrane permeability. The permeability of a cell can be transiently changed by applying various factors, conditions, and external environments (Fig. 2). Regardless of these changes, the viability of the cells should be preserved. It is an undoubted fact that changes in cellular permeability involved complex and multistep processes. For example, kinetic studies of electromediated permeabilization by Teissie et al. (2005) include several stages: (1) trigger step: mechanical stress induces the membrane potential difference; (2) expansion: time-dependent membrane transition after the electric field is maintained at an overcritical value; (3) stabilization: recovery of the cell membrane as soon as the external field decreases below the critical permeabilizing threshold; (4) resealing step: the slow decrease in the number of permeabilized cells with post-trigger incubation time; and (5) memory step: cell viability is preserved but membrane structural and physiological properties recover on a much longer timescale before normalizing. The recovery of the cell after application of the physical disruption is very important and requires further research to determine what portion of cells return to a normal physiological state and which aspects of cell function are permanently compromised.
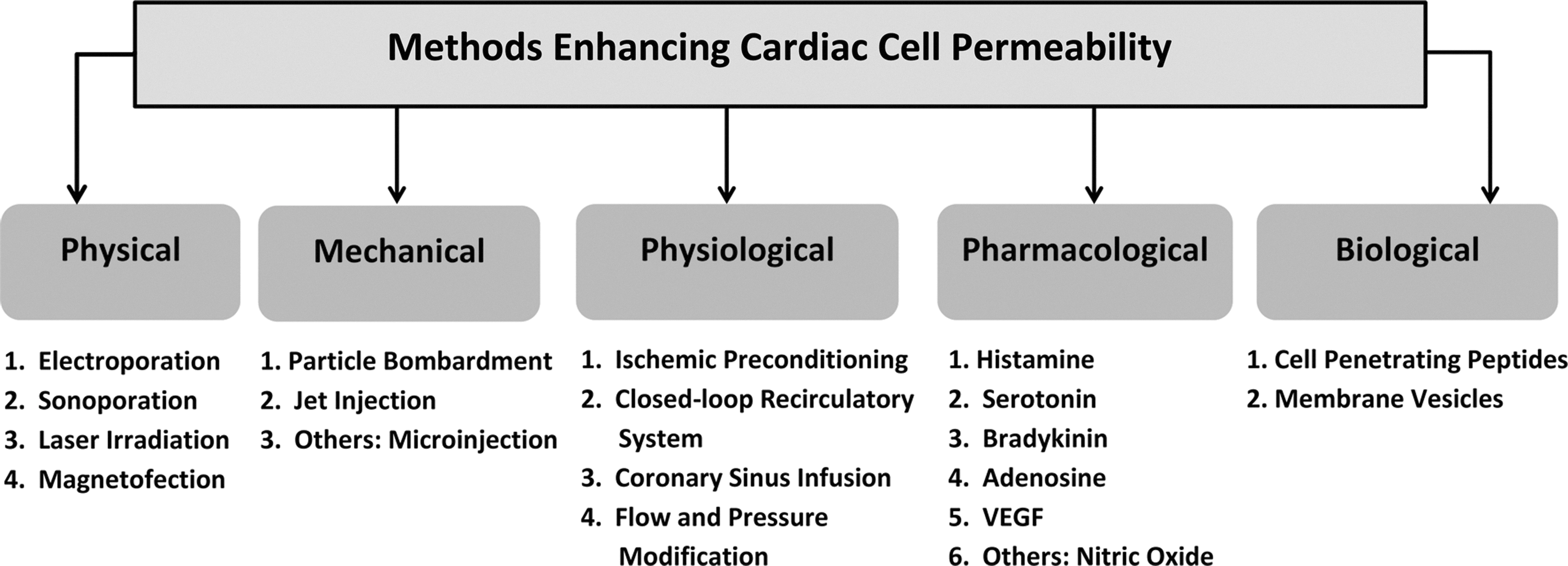
Methods enhancing cardiac cell permeability.
The endothelial barrier
It is known that, in addition to regulating blood perfusion, a major function of vascular endothelia is to provide a semipermeable barrier to control blood–tissue exchange of different macro- and micromolecules, including viruses. The microvascular wall permeability is caused by the presence of long cylindrical pores in the endothelial cells. Molecular movement through the endothelium is governed by Poiseuille's law, where hydraulic conductivity is a function of fluid viscosity and pore radius and therefore depends on molecular size and the state of permeability pathways. Presently, two types of pores in capillary endothelium are described: small pores (4 nm), representing the normal permeability pathways through intercellular junctions, and large pores (25–30 nm, sometimes up to 80 nm), representing permeability pathways for larger molecules occurring between endothelial cells. Moreover, there are endothelial vesicles and transendothelial pores that pass across the endothelial cells. The permeability properties of the barrier are regulated via interactions between endothelial cells, the basement membrane and supported matrix and cells in the surrounding tissue. The mechanism by which the vectors pass through significantly smaller endothelial pores is poorly understood, and the determination of optimal vector dimensions requires further investigation.
The cell membrane barrier
The connection between cells and the environment is the plasma membrane, composed of phospholipid and protein molecules. The plasma membrane ties the cytoplasm of the cell to the external environment, regulating cell permeability and providing a conduit for cell-to-cell communication. Moreover, the cell membrane separates the intracellular components from the extracellular space and is a barrier to free diffusion of plasmid DNA (Stephens and Pepperkok, 2001). The mechanism of this process is based on the capture of plasmid DNA by the plasma membrane's receptors and internalization into target cells (Budker et al., 2000).
Viral vs. nonviral vectors
Currently, no single vector system is optimal for potential gene therapy applications. A perfect vector would be administered by a noninvasive route, target the desired number of myocytes, and express the required amount of transgene product (Kay et al., 2001). Although a plethora of viral and nonviral vectors are now available for cardiac applications, the optimal vector for daily clinical practice remains elusive (Wasala et al., 2011; Tang et al., 2012). In general, recombinant viral vectors represent the popular delivery vehicles of therapeutic genes because of their superior transduction efficiency, favorable cell uptake, relatively easy intracellular trafficking of packaged DNA to the nuclei, ability to escape lysosomic degradation, and facilitation of long-lasting gene expression (Bish et al., 2011). However, they stimulate a specific immune response, and it is therefore impossible to readminister the vector. In addition, retro- and lentivirus could evoke insertional mutations while they integrate into the host genome (Hinkel et al., 2011). In contrast, plasmid DNA molecules are easier to produce pharmacologically and have low immunogenicity and toxicity. However, low levels of gene transfer and short-lived expression still hinder their use (Lechardeur et al., 2005; Medina-Kauwe et al., 2005).
Intracellular trafficking of viral vectors
Establishment of the efficient viral vector depends on knowing the sequences for viral particle assembly, genome packaging, and identification of paths to target cells and nuclei (Coura Rdos and Nardi, 2008). Some vectors are able to integrate into the host genome, while others (like adenoviruses) do not and remain episomal. Persistent transgene expression is provided only by integrating viruses.
The most commonly used viral vectors are based on adeno-associated viruses (AAV), adenoviruses and retroviruses, or lentiviruses. Each type of viral vector has advantages and limitations. To use the adeno-associated viral vector as an example, the main steps of intracellular trafficking occur as follows. Step 1: AAV binds to the surface of the target cell through cell receptors and co-receptors (virus serotype or type of capsid variant used to generate the recombinant virus is critical). Step 2: Receptor-mediated endocytosis of the virus (dependent on the amount and effect of AAV co-receptors on the cell surface and degree of activity of the cellular pathways). The next three steps involve movement to the nucleus. Step 3: Intracellular trafficking through the endosomal compartment and vesicular trafficking. Step 4: Endosomal escape of the virus. Step 5: Intracellular trafficking of the virus to the nucleus and nuclear import. Step 6: Viral uncoating. Step 7: Genome conversion of the single-stranded AAV genome to double-stranded DNA intermediates capable of expressing the encoded gene (Ding et al., 2005; Medina-Kauwe et al., 2005; Coura Rdos and Nardi, 2008; Nonnenmacher and Weber, 2012).
Intracellular trafficking of nonviral vectors
In recent years, much effort has been applied to the development of new nonviral vectors in order to achieve effective and long-lasting gene expression. Their great advantages include low toxicity, noninfectious properties, and capacity to transfer large gene molecules. The major limitation of naked DNA delivery is insufficient level of gene expression and limited distribution. Therefore, the various physical and mechanical methods used are described below. These considerations favor the utilization of synthetic DNA delivery systems using chemical carriers such as polyplex, lipoplex, or a mixture of these—lypopolyplex (Al-Dosari and Gao, 2009). These carriers' properties include the following: condensing DNA into complexes to protect it from nucleases; targeting DNA to target cells; strengthening DNA transfer to the cytosol or nucleus; and slowly releasing DNA to achieve prolonged expression (Lavigne and Gorecki, 2006).
An understanding of the cellular trafficking of plasmid DNA, complexed with synthetic molecules or phospholipids, will help to provide a strategy for overcoming the low efficiency of nonviral gene transfer. After internalization, plasmid DNA must overcome endo-lysosomal entrapment, cytosolic sequestration, and nuclear exclusion. Moreover, DNA can be subjected to metabolic degradation in the endo-lysosomal compartment and in the cytoplasm (Lechardeur et al., 2005). To reach the plasma membrane of target cells, the plasmid DNA must diffuse through the extracellular matrix without being degraded by the extracellular nucleases. This process depends on the amount of nucleases and on the amount of extracellular matrix components like collagen and hyaluronic acid (Escoffre et al., 2010). These factors significantly reduce gene transfer and require at least 105 plasmids per cell in order for a few molecules of DNA to enter the nucleus (Lechardeur et al., 2005). Thus, to increase the efficiency of plasmid DNA transfer, the strategy should combine improving the diffusion into the cells and nucleus, extending the time of gene expression, and limiting the degradation of plasmid DNA.
Minicircle vectors
One of the promising directions for solving these problems is the use of minicircle vectors. Minicircles are supercoiled, minimized plasmids with no bacterial backbone sequences that encode only the therapeutic gene of interest with regulatory sequences. Minicircle DNA has several advantages over both viral-based and conventional plasmid vectors. It has a better safety profile and allows for a larger expression cassette and possibly easier clinical translation. Compared with regular plasmid vectors, minicircles have a significantly higher level and longer duration of transgene expression (Mayrhofer et al., 2009). This was confirmed by data from a study that used short hairpin RNA minicircle vectors injected intramyocardially after ligation of the left anterior descending artery (LAD) in adult mice. Functional studies showed improvement in cardiac function, increased neovascularization, and decreased apoptosis in areas of injured myocardium (Huang et al., 2011).
Physical Methods Enhancing the Cell Membrane Permeabilization
Electric field-based transfection (electroporation)
The basic principle
Electroporation (EP) involves the physical implantation of DNA molecules into cells, using an electric field (Fig. 3A). Currently it is the most commonly used physical method. The technique includes the exposure of the cell membrane to high-intensity electrical field, which destroys the integrity of the cellular barrier. Typically, this damage is temporary and local. During this period the cell membrane becomes highly permeable to exogenous molecules. Using electron microscopy, it has been shown that when the electrical signal reaches the cell, membrane pores can increase in size up to 120 nm in 20 milliseconds (Chang, 1992). In such a condition, the membrane can allow passage of DNA into the cell.

Physiological mechanism
The precise mechanism by which these pores open or close and subsequently take up DNA remains the subject of investigation. It is currently believed that electroporation affects the passive transport of a plasmid when it is in close proximity to the cell surface. Alternatively, it has been suggested that DNA can also actively bind to the cell membrane and then be internalized when the electric field is applied (Gehl, 2003). Previously it had been argued that DNA can itself facilitate pore formation via a direct interaction with the cells (Lurquin, 1997). It has also been suggested that the negative charge of DNA might lead to its electrophoretic movement, thereby easing the transition through cell membrane (Satkauskas et al., 2002). The efficiency of gene transfer by electroporation is influenced by different physical and biological factors. These include pulse duration, electric field strength, DNA concentration, cell size, and type. Different electroporation conditions are required for penetration of molecules of different sizes. In addition, longer pulse durations lead to the creation of larger pores, which stay open longer (Golzio et al., 2002).
In most studies, electroporation increased gene expression by 100-to 1000-fold compared to injection of naked plasmid DNA. Once a critical voltage has been achieved (on the order of 200 V/cm in vivo), membranes become more permeable. Plasmid movement into the cytoplasm was relatively slow and continued after the application of the electrical field ended (Marshall et al., 2010).
Instrumentation
Devices for EP consist of a pulse generator and an applicator, which includes the electrodes. The three-dimensional geometry of the electrodes determines the strength, homogeneity, orientation, and shape of the electrical field, as well as the current flow and the total energy transferred to the target tissue. Different types of electrodes are used in vivo for electrotransfer depending on the characteristics of the target tissue: surface, needle, and catheters for hollow organs. In the case of the meander electrode, an electrical field is generated between positive and negative electrodes with part of the field entering the tissue. This electrode design is the least invasive. With the caliper electrodes, a near-uniform electrical field is generated between the electrode plates. The pattern of electrical pulses also varies considerably between studies ranging from moderate voltage (e.g., 200 V/cm) pulses of tens of milliseconds to high-voltage microsecond pulses.
Limitations
One of the limitations of electrotransfer is that there can be substantial damage associated with the procedure. Sometimes there are difficulties in the placement of electrodes into the tissues, and the high voltage applied might affect the genomic DNA stability (McMahon and Wells, 2004). Another limitation is that transgene expression is not homogeneously distributed in the treated tissue. The distribution of the electric field within the tissue probably affects the efficacy of electroporation during gene transfer (Wells, 2004).
Electroporation-mediated gene transfer to the heart
The uptake of DNA-encoding green fluorescent protein (GFP) or luciferase following electrical shock was demonstrated in embryonic chick hearts. To compare EP with another gene transfer method, adenovirus/GFP studies were performed with the same model. EP was equivalent to an adenovirus dose of 106 transduction units. Heart subjected to six shock triplets expressed a luciferase activity level of 8×106 LU/mg protein, which increased to 13×106 LU/mg protein in hearts subjected to 12 shocks (Harrison et al., 1998). The development of synchronous (with the QRS complex) EP pulse delivery method in pigs allowed for a five-time increase in Vascular Endothelial Growth Factor (VEGF) expression at day 2 post-treatment compared to the injection of plasmids alone. Ventricular fibrillation was absent using this technique (Marshall et al., 2010).
Another study confirmed that EP is a viable approach to deliver plasmid DNA into the myocytes of large animals. A significant increase in luciferase expression was found with application of different electrodes, pulse widths, and electrical fields. Satisfactory expression of GFP within the swine myocardium in vivo was observed. To avoid fibrillation, electric pulses had to be synchronized with electrocardiogram (ECG), and the animal could not be hypokalemic. The important conclusion is that a large amount of DNA is not needed to get significant levels of myocardial expression during electroporation (Hargrave et al., 2012). Repeated long and short electropulses produced GFP expression in the left ventricles of beating rat hearts. Continuous pulses were 300-fold more effective compared to plasmid injections, but the incidence of fibrosis and apoptosis increased. Short electropulses were able to reduce fibrosis by two-fold (Eigeldinger-Berthou et al., 2012).
Interestingly, in a report by Ayuni et al. (2010), naked plasmid DNA was injected into the coronary sinus of male adult rats. The heart was then positioned between plate electrodes and an electric field was applied. Luciferase activity (RLU/mg protein) observed on day 1 after EP-mediated gene transfer was approximately four times higher than in nonelectroporated heart. Still, activity showed a gradual decrease over 7 days (Ayuni et al., 2010).
Ultrasound-based transfection (sonoporation)
The basic principle
Ultrasound (US) has been used in clinics for a relatively long time for both therapeutic and diagnostic purposes. Recently it was found that ultrasound makes biological membranes transiently permeable (Fig. 3B). US covers a broad range of frequencies and wave-forms, but attention has been focused on sinusoidal probes at megahertz frequencies. Applying US to a liquid leads to the formation of vapor-filled bubbles, or cavities, in the solution. The formation and collapse of the ultrasound-induced bubbles is called acoustic cavitation. It has been thought that this mechanism can produce transient membrane permeabilization (Wells, 2004; Mehier-Humbert and Guy, 2005; Villemejane and Mir, 2009).
Physiological mechanism
With respect to gene therapy, it is contested that under conditions of acoustic cavitation mechanical disruption and collapse of active bubbles occurs, and the associated energy release can permeabilize adjacent cell membranes for DNA delivery to the cytoplasm. To promote ultrasound-mediated gene transfection, one usually uses a contrast agent, which consists of elastic and compressible gas-filled microbubbles. Microbubbles can be destroyed by ultrasound waves. Rupture of microbubbles may form localized jets that would function like microneedles, penetrating pathways through the cell membrane. The contrast agents lower the transfection threshold by acting as cavitation nuclei, and a new generation of contrast drugs (such as perfluorocarbon) employs special coating materials to stabilize the bubbles. It was demonstrated in electron microscopy that US led to transient formation of holes (less than 5 μm) in the cell surfaces treated with Optison (octa-fluoropropane contrast agent) (Taniyama et al., 2002). Formation of small membrane pores enhanced the direct transfer of genetic material into cells (Endoh et al., 2002).
Instrumentation
Both diagnostic equipment used in clinical settings and specific ultrasound devices have been used for sonoporation.
Limitations
Optimization of ultrasound-mediated gene transfer depends upon several factors, including transducer frequency, acoustic pressure, pulse and exposure duration, and concentration and formulation of contrast agents.
Sonoporation-mediated gene transfer to the heart
Bekeredjian et al. (2003) showed that US-mediated destruction of microbubbles directs plasmid transgene expression to the rat heart better than viral vectors. Time course evaluation showed high expression in the first 4 days, with a rapid decline thereafter. Interestingly, repeated treatment produced a second peak of gene expression. Naked plasmid DNA and short interfering RNA were transduced into the left ventricles of murine hearts by means of US, using third-generation microbubbles. Results showed significant expression of the marker gene in the subendocardium and antero-septal regions of the myocardium (Tsunoda et al., 2005).
In a rat model of acute myocardial infarction (MI), naked plasmid-encoding hepatocyte growth factor (HGF) was infused through the left ventricular chamber. Immunohistology 7 days later revealed significant myocardial expression of HGF only in the US-mediated microbubble destruction group (Kondo et al., 2004). In another study, rats underwent LAD ligation followed by VEGF 121 injection into the tail vein. Formation of new blood vessels and VEGF protein expression were higher in the group that received US microbubble destruction (Zhigang et al., 2004).
Laser-based transfection
The basic principle
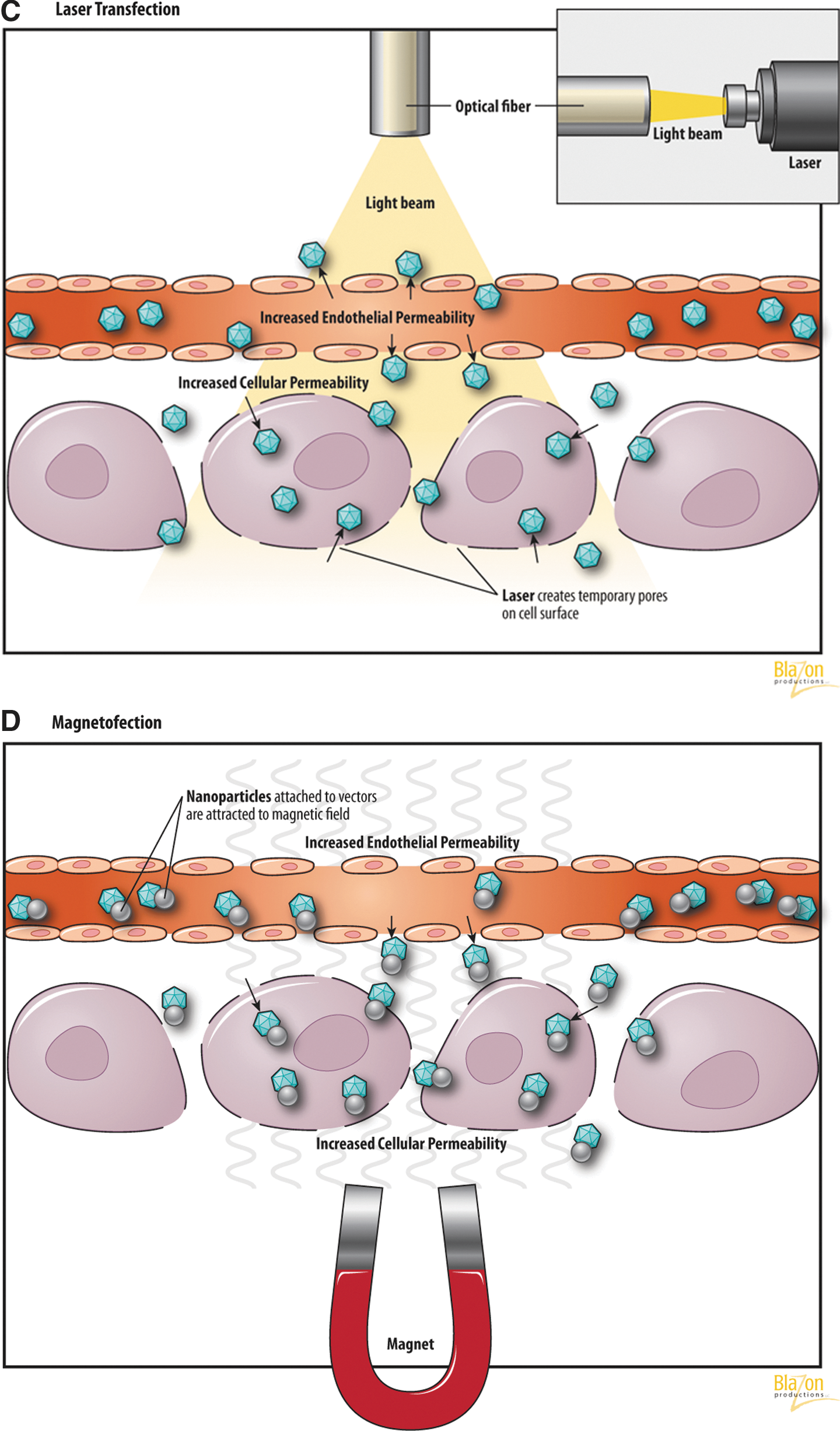
In the last few years several laser systems for cellular biotechnology have been developed (Zeira et al., 2003; Yao et al., 2008). It has been suggested and since confirmed that a highly focused laser beam can produce tiny holes in the cell membrane to facilitate uptake of exogenous DNA (Tsukakoshi et al., 1984) (Fig. 3C). The laser beam is focused onto the target cell via a lens. The permeability of the cell membrane is modified at the site of the beam impact by local temperature changes.
Physiological mechanism
It has been shown that the effect of laser irradiation is not affected by receptors or interactions with the cell membrane. Its efficiency depends upon the difference in osmotic pressure between the cytoplasm and the extracellular matrix (Mehier-Humbert and Guy, 2005). Kurata el al. (1986) used two kinds of laser beams: a pulsed laser to make the holes in cell membranes and a continuous laser to trap the cells. It was demonstrated that lasers create transient pores in the cell membrane approximately 2 μm in diameter that close shortly after the cessation of radiation (Kurata et al., 1986). Laser light can be delivered via modern transcutaneous intravascular catheter-based transfer through optical fibers.
Instrumentation
Laser gene transfer requires a laser source, the power of which is controlled by a pulse generator.
Limitations
This technique for gene transfer has not yet been extensively researched and much remains unknown about the types and extent of damage that is caused by laser irradiation. The laser equipment required for this technique can also be prohibitively expensive.
Laser-based gene transfer to the heart
Laser-targeted transfection of cardiac neonatal rat cells was demonstrated with GFP plasmids. Successful transfection after 48–72 hr was five times higher than in the control (Nikolskaya et al., 2006). Another study showed that the transfection efficiency of plasmid DNA–encoding VEGF was improved with the use of transmyocardial laser revascularization. Moreover, postinfarction wall motion abnormalities were completely reversed within 6 weeks after using this strategy (Sayeed-Shah et al., 1998). On the other hand, a study with catheter-based delivery of plasmid vectors with a reporter gene in porcine ischemic myocardium failed to show that laser injury augments gene expression above levels present with gene transfer alone (Fuchs et al., 2001). This technique requires further research.
Magnetic field-based transfection (magnetofection)
The basic principle
Strong magnetic fields (MF) can also be used to provide an energy source to assist gene transfer when plasmid DNA or virus vectors are coupled with magnetic nanoparticles (Fig. 3D). MF utilizes magnetic nanoparticles made of iron oxide and coated with cationic lipids or polymers to complex with DNA. These magnetic particles are concentrated into the target cells by the influence of an external magnetic field. This process is applicable to viral as well as nonviral gene vectors and can be adapted to DNA, siRNA, mRNA, etc. (Scherer et al., 2002; Kami et al., 2011).
Physiological mechanism
The mechanism of magnetofection appears to be the rapid accumulation of the vector on the target cells. Also it has been shown that the magnetic field can cause changes in venular permeability with extravasation (Plank et al., 2003). Similar to the mechanism of nonviral vector-based gene delivery, the cellular uptake of DNA is accomplished by endocytosis and pinocytosis (a form of endocytosis in which small particles are brought into the cell, forming an invagination, and then suspended within small vesicles). The actual uptake of vectors into the cells depends on increased tissue permeability after application of MF. Advantages of this method include low vector dose, reduced incubation period to achieve transfection, and inert properties of magnetic nanoparticles. Also it should be noted that the nanoparticles are coated with plasma proteins (Plank et al., 2003; Kami et al., 2011).
Instrumentation
Strong magnetic field and magnetic nanoparticles are needed.
Limitations
The equipment required for this technique can be prohibitively expensive.
Magnetofection of the heart
In porcine aortic endothelial cells, MF increased transduction of a luciferase or a B-galactosidase reporter to achieve an efficiency of 37.5% of cells (Krötz et al., 2003). Magnetic nanoparticles combined with plasmid DNA–expressing GFP were injected into mice and directed to the heart. Sections of the heart tissue from the mouse with the magnet showed extensive expression of GFP in comparison with the control mouse without the magnet (Kumar et al., 2010). Magnetic nanobead/polymer/DNA complexes had a 36- to 85-fold higher transfection efficiency under the magnetic field compared with control in vitro. In an in vivo application, the epicardial magnet effectively transferred these complexes to the heart resulting in strong reporter and therapeutic gene expression (Li et al., 2008).
Mechanical Methods
Gene-mediated particle bombardment
The basic principle
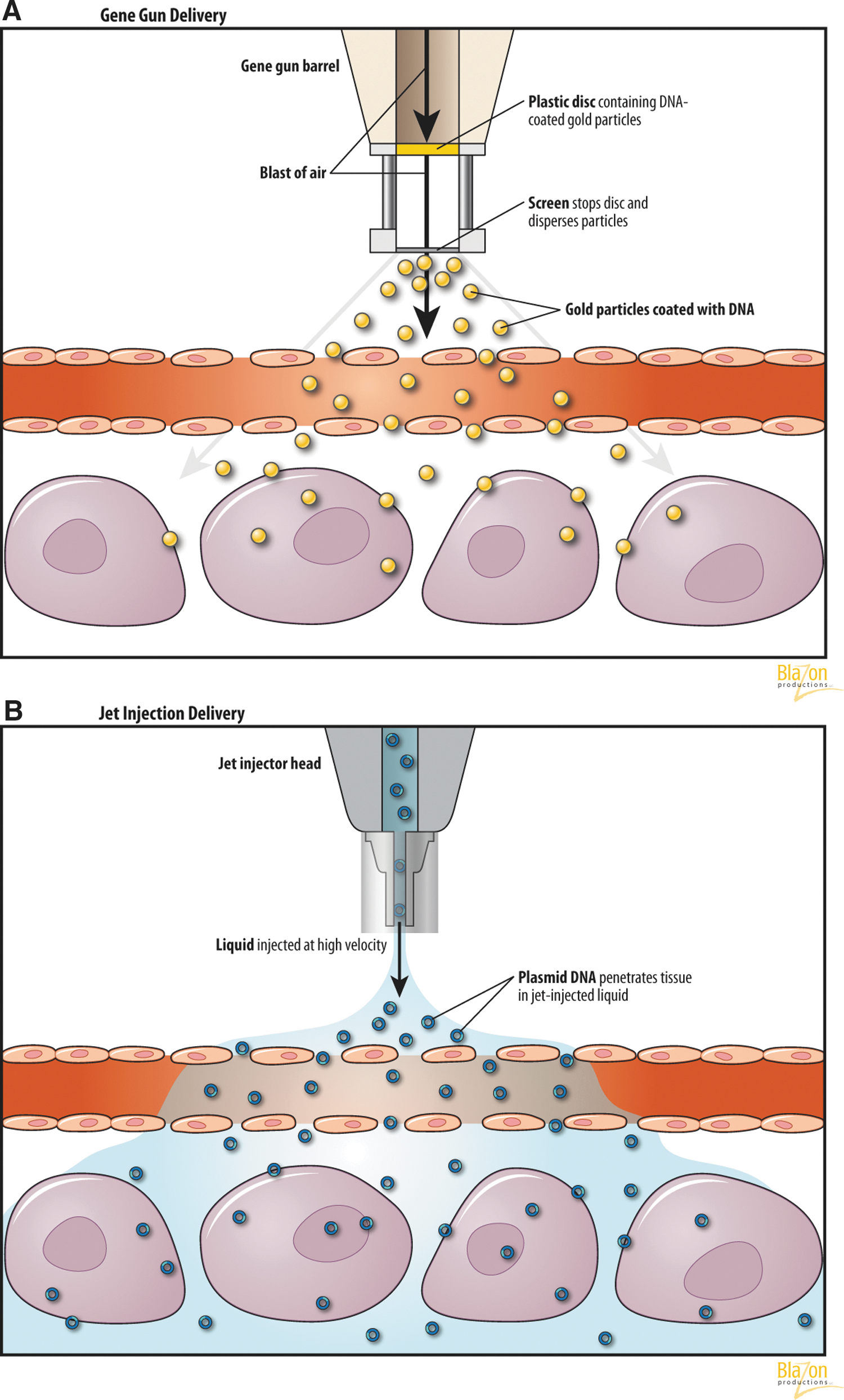
Particle bombardment (also called gene gun or ballistic DNA transfer) utilizes heavy metal particles, introduced with a high velocity pressurized inert gas, into the target cell (Fig. 4A). Naked DNA can deposit onto these particles, and after entering the cell they are gradually released. Acceleration can be achieved by a high-voltage electric spark or a helium pressure gun. For optimal gene expression, the following parameters have been taken into account: (1) the properties, density, and sizes of gold particles for bombardment; (2) the DNA doses; and (3) the discharge voltage for optimal gold particle penetration. Using a submicrogram amount of DNA per bombardment, 1000 to 10,000 copies of DNA can be delivered to each target cell (Yang and Sun, 1995).
Physiological mechanism
DNA-coated particles are brought into the cell through the holes in the plasma membrane, resulting in enhanced gene expression. Some authors believe that it may be possible to employ a gene gun to transfect cells that are relatively resistant to other delivery systems (Klein et al., 1987; Yang et al., 1990). In this method, DNA or RNA adhere to metal particles (gold or tungsten), then the DNA-particle complex is accelerated and shot into the target tissue. Uncoated metal particles could also be delivered through a solution containing DNA surrounding the cell, thus picking up the genetic material and proceeding into the living cell.
Limitations
The efficiency of the gene gun transfer could be dependent on cell type, cell growth condition, and gene gun settings.
Particle bombardment transduction into the heart
The study of Nishizaki et al. (2000) demonstrated that plasmid DNA can be introduced into cardiomyocytes using a gene gun and that transfer of the Epstein-Barr virus-based episomal vector results in 6-week gene expression in vivo. A rat model exhibited GFP expression in the beating heart up to 3 weeks after using in vivo helium-gun-mediated gene transfer. These authors believe that gene gun technology has advantages over other methods in terms of safety, noncytotoxicity, the small amount of DNA needed, and the short time required to perform gene transfer. Moreover, it is independent of target cell type (Matsuno et al., 2003).
Jet injection
The basic principle
A ballistic method jet injection is performed using a high-speed pressurized gas, usually CO2. The injection creates pores in cellular membranes and allows for intracellular gene transfer (Fig. 4B). The penetration power depends on two factors: the applied pressure of the gas and the tissue's resistance (Al-Dosari and Gao, 2009). Levels of gene expression by jet injection are 50-fold higher than by conventional needle injection (Ren et al., 2002). The jet injection gene transfer is usually well tolerated without side effects. This method is described only for nonviral-based techniques of gene transfer and uses a high pressure (usually 1–3 bars) device to force microdroplets of liquid into the tissues. Micromolecules of different substances, such as nucleic acids diluted in liquid, are injected ino target tissues without a needle. Pressure used is typically less than 3–4 bars, and the velocity of the droplets range from 100 m·s−1 to 200 m·s−1. The velocity of the fluid in the jet injection contributes to the distribution in the tissues, whereas the diameter of the jet and the injected volume limit the penetration depth (Arora et al., 2007). The efficiency of this method depends on nozzle diameter (150–300 μm), velocity of the liquid jet, and distance between the nozzle and surface of the tissue (Rajaratnam et al., 1994).
Physiological mechanism
The penetration of the injected molecules inside the cells is a consequence of the pressure caused by the liquid. The high pressure of the jet creates a hole in the tissue surface; the depth of this hole is increased due to the accumulation of fluid. The use of intramuscular jet injection of DNA combined with electroporation was demonstrated to be feasible in a mouse model (Horiki et al., 2004).
Instrumentation
Jet injector devices vary according to the velocity of injection and injection volume: high (more than 100 μL) and low (20–30 μL).
Limitations
Mechanical injury to the cardiomyocytes is possible because of the high pressure of the gas. This process also has an unknown transfection efficiency and inflammatory response.
Jet injection into the heart
We and others could not find publications on the applications of this method for cardiac gene therapy (Al-Dosari and Gao, 2009; Villemejane and Mir, 2009).
Physiological Methods
Ischemic preconditioning
Ischemic preconditioning (IP) is an adaptive endogenous mechanism activated after brief periods of interruption of coronary flow, which provides the heart protection against myocardial injury. In recent years, considerable progress has been made toward the identification of potential cell triggers, intracellular signaling cascades, and the end-effectors involved in IP. There is ample evidence suggesting a role of endogenous paracrine mediators, released during ischemia and acting on local receptors, as triggers of IP. It has been shown in various species that the involvement of many mediators such as adenosine, acetylcholine, catecholamines, angiotensin, bradykinin, endothelin, etc., stimulate specific receptors on the cardiomyocyte cell membrane.
Most of the above listed substances directly target the cardiac vascular endothelium after oxidative stress induced by ischemia, which are associated with ultrastructural changes of myocytes followed by endothelial activation with cell attachment and disruption of intercellular clefts (Skepper et al., 1998). Increased circulating levels of pro-inflammatory cytokines (e.g., interleukins and tumor necrosis factors) with activation of kinase cascades are a clear manifestation of host defense against myocardial injury. This is important in the signal transduction pathways of ischemic preconditioning, resulting in phosphorylation and activation of different cell transmitters in coronary arterial smooth muscle and endothelium (Minamino et al., 1995).
Does ischemic preconditioning have an effect on myocardial gene transfection? There are many described techniques to transduce myocardium after ischemic preconditioning (Figs. 5 and 6). Logeart et al. (2000) showed that short periods of ischemia are required to obtain significant myocyte transduction during single-pass gene delivery in the isolated rat heart model and in vivo when adenoviruses were delivered downstream of an occluded artery. Myocardial IP with 1-min occlusion of two main coronary arteries improved marker gene expression (Hayase et al., 2005). Temporary cardiac arrest of 2 min allowed the transfection of 18% of cardiomyocytes, whereas extending time to 5 min resulted in a cardiac transfection of 43% of cells (Ding et al., 2004). A pig model demonstrated that retrograde delivery during 10 min of ischemia increased reporter gene expression compared with control, and the use of two periods of ischemia resulted in more homogeneous transmural expression (Boekstegers et al., 2000). These and other data confirm that IP is involved in the effectiveness of gene transfer.
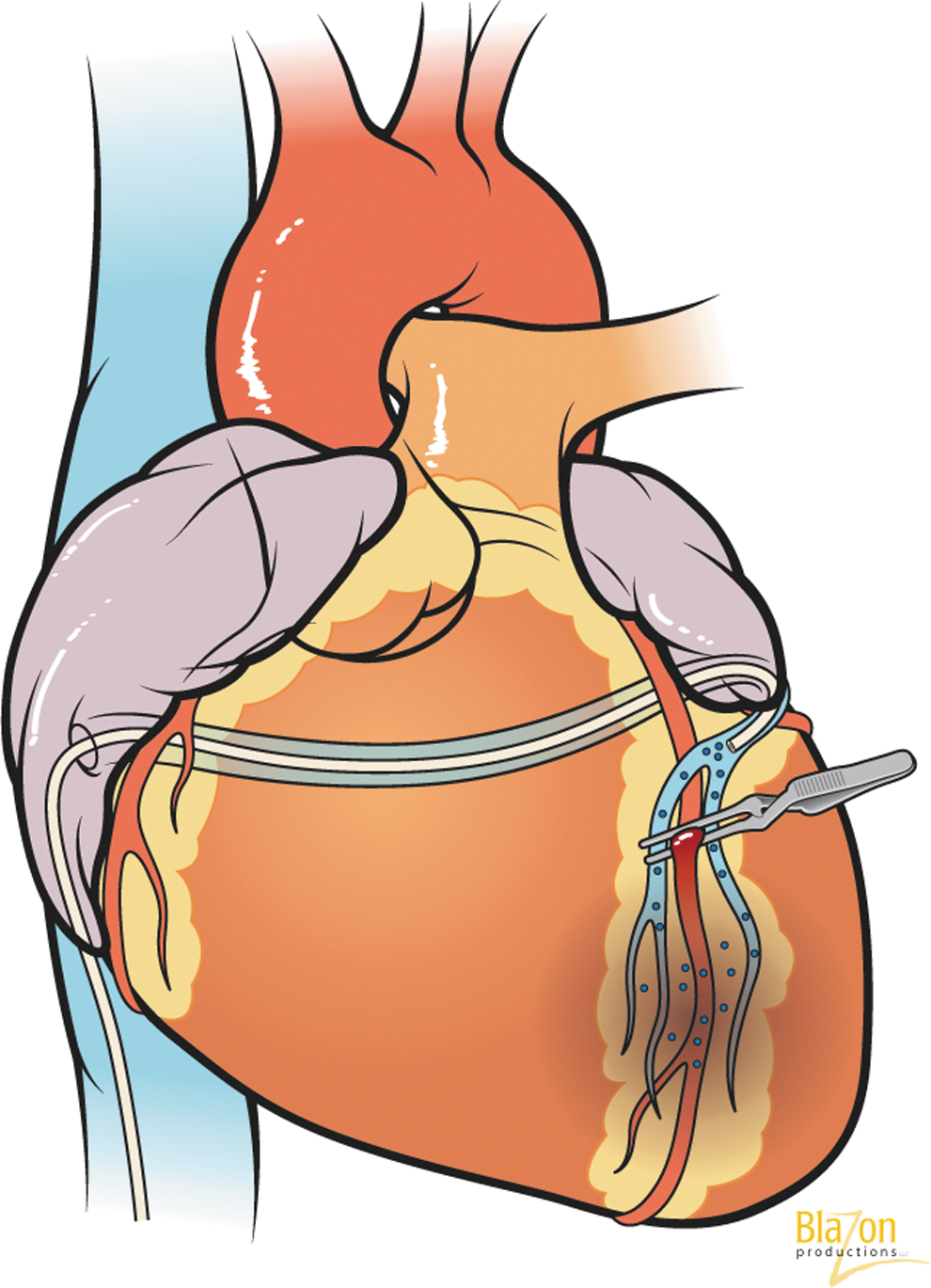
Left anterior descending artery occlusion during transvascular intracoronary retrograde gene delivery through coronary sinus (creation of ischemic preconditioning).
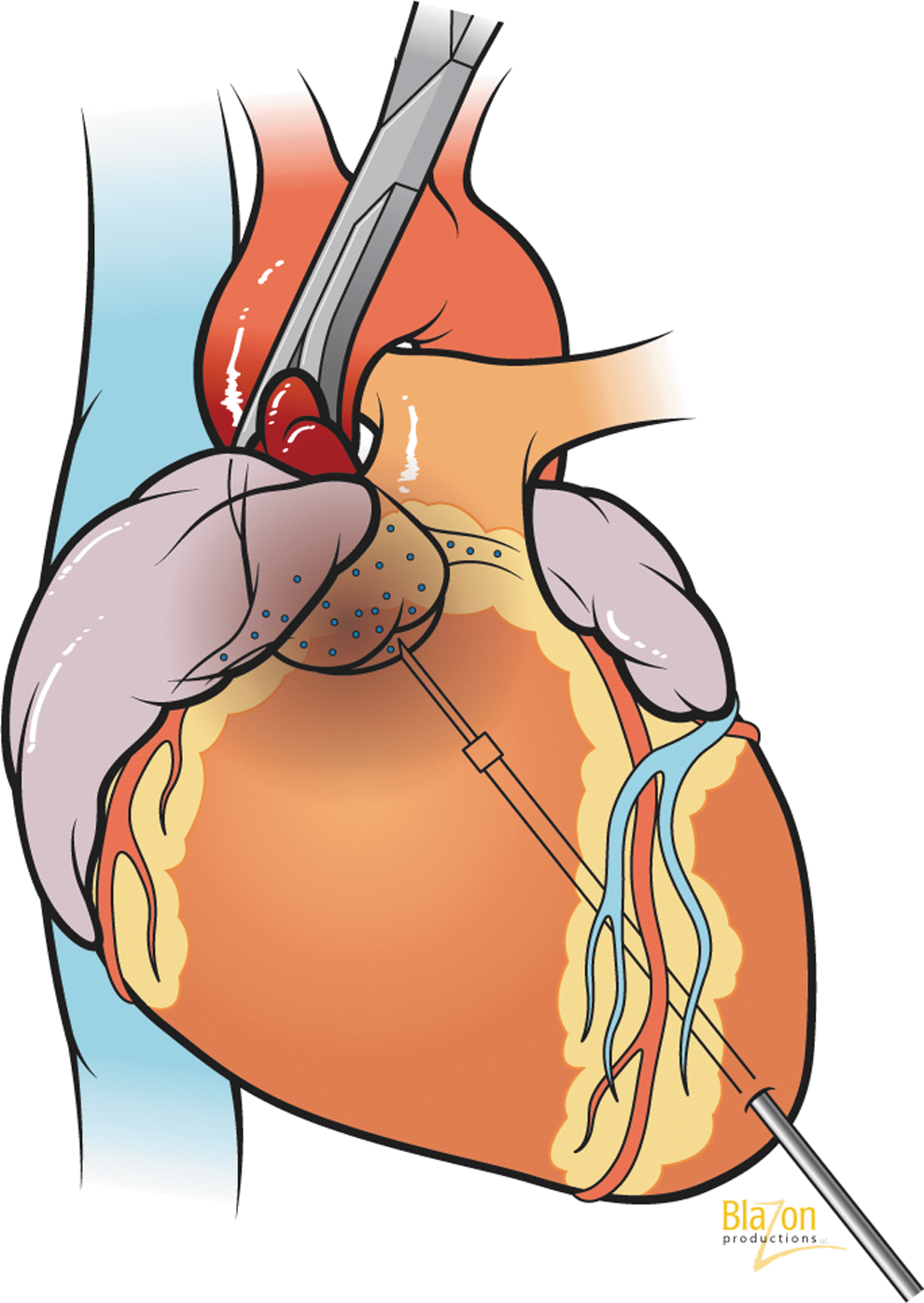
Ascending aortic cross-clamping during transvascular intracoronary antegrade gene delivery: catheter inserted into left ventricle apex and gene delivered under aortic valve. Cross-clamping directs the vector through coronary arteries only (creation of ischemic preconditioning).
Closed-loop recirculatory systems
Separation of cardiac circulation from the systemic, and creation of a “closed-loop“ recirculatory system, can effectively increase transduction efficiency and modulate endothelial permeability in coronary vasculature (White et al., 2011; Tilemann et al., 2012). Bridges et al. (2002) first used this system during cardiopulmonary bypass (CPB) (Fig. 7). The rationale for using CPB is that when you work on a stopped heart, you can use different routes for gene delivery, vary the contact time between gene construct and vascular endothelium, use any agents for endothelial permeability, and control temperature and ionic composition of the perfusate. A percutaneous catheter-based “closed-loop” configuration was also developed recently (Kaye et al., 2007) (Fig. 8). There is no doubt about the bright prospects of this direction and future use of such systems in clinical settings.

Cardiopulmonary-based closed-loop recirculatory system. The system includes two separate circuits that allows for complete cardiac isolation and provides recirculation only in the coronary vasculature. Other advantages are increased time of recirculation, possibility for washing out the vector, and retrograde transcoronary sinus gene delivery.
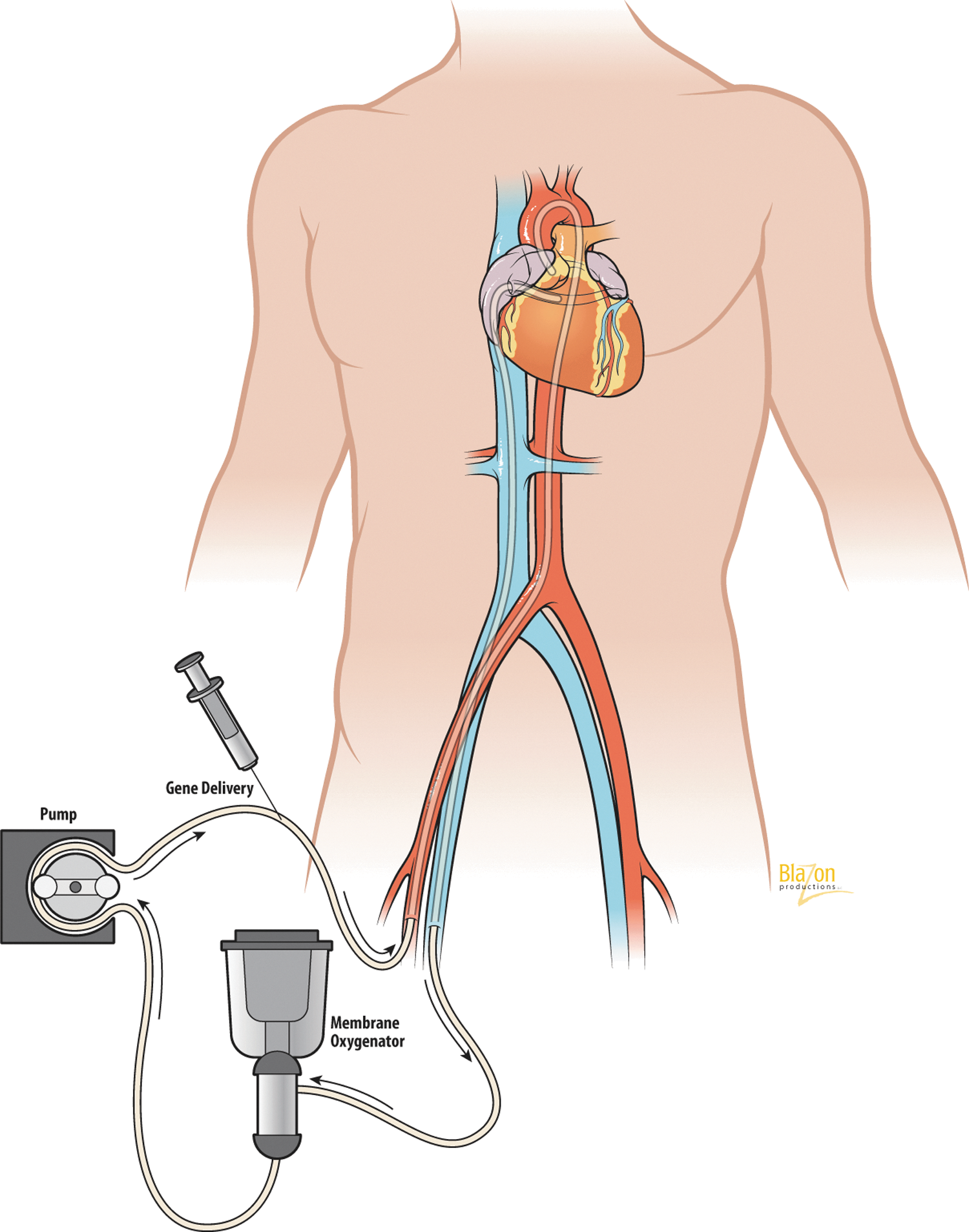
Percutaneous catheter-based closed-loop recirculatory system. Coronary venous blood is drained from the coronary sinus. Following oxygenation, the blood is returned to the left main coronary artery via a roller pump. The gene of interest is delivered into the antegrade limb of the circuit.
Coronary sinus gene infusion
The coronary sinus has become a clinically important structure due to its access for different cardiac procedures. Retrograde coronary venous drug delivery can preserve myocardium during ischemia and has been used clinically to deliver oxygenated blood during unstable angina or high-risk coronary angioplasty. It was proven that this method of perfusion accelerates coronary thrombolysis, preserves global and regional myocardial function during open-heart surgery, and limits infarct size. Pressure-regulated infusion into the anterior cardiac vein through the coronary sinus substantially increases reporter gene expression in the targeted territory and provides homogenous beta-galactosidase expression compared to intramyocardial injection (Raake et al., 2004). The use of a pig model showed the advantage of this delivery route as compared to antegrade delivery. The authors explained this phenomenon by showing that the passage time of a marker gene increased more than 10-fold when delivered retrograde as opposed to antegrade (Boekstegers et al., 2000). Later, these data were confirmed by other authors (Hou et al., 2003; White et al., 2011). Retrograde gene delivery changes the hydrostatic and osmotic pressure, thereby increasing the capillary filtration ratio in the venous part of the capillary bed. Another advantage is the ability to overcome the resistance of precapillary sphincters located before arterial capillaries (Katz et al., 2012).
Flow and pressure modification
To evaluate the dependence on coronary flow rate, the hearts were perfused with virus-containing solution for 120 min at 10–40 ml/min. Infection was enhanced by a factor of four when the flow rate was increased from 10 ml/min to 30 ml/min or greater. The improvement of gene transfer at higher flow rates was independent of perfusion pressure. A more likely cause of the flow dependence is the presence of precapillary sphincters that open in response to increasing flow rates. Flow can open the surface areas for virus delivery and shorten diffusion distances to myocytes (Donahue et al., 1997). Similar results in a piglet model were obtained after adenoviral/β2adrenoreceptor (βAR) delivery. Left ventricular βAR expression was significantly higher at flow rates of 120 cc/min and 80 cc/min compared to 10 cc/min (Emani et al., 2003). Also, the importance of the myocardial perfusion gradient on expression of marker genes was shown in murine coronary vascular endothelium after transfection of AAV and adenoviral vectors (Champion et al., 2003). Increasing flow and pressure caused enhancement of microvascular permeability and thus more efficient virus distribution within the capillary layer (Sasano et al., 2007).
Pharmacologic Methods
The use of pharmacological agents was apparently one of the first methods with described effects on capillary permeability. In the mid-90s, scientists began to use various vasoactive mediators to increase cardiac gene transduction. It was based on the well-known ability of different neurotransmitters (histamine and serotonin) and active peptides (bradykinin) to increase endothelial permeability by means of paracellular leakage of plasma fluid and protein. Later it was found that releasing inflammation stimuli such as thrombin, VEGF, or activated neutrophils can cause dissociation of cell–cell junctions between endothelial cells, as well as cytoskeleton contraction, leading to widened intercellular spaces that facilitate transendothelial flux (Kumar et al., 2009). These agents act at the level of the post-capillary venules to increase permeability by causing endothelial contraction and disruption of tight junctions.
The role of extracellular calcium was also evaluated in terms of enhancing virus access to myocytes at the level of endothelial junctions. It was found that a reduction of the extracellular calcium concentration would increase the surface area available for virus passage between endothelial cells. In addition, the regulation of calcium transient is necessary for maintaining the connection between myocytes and the extracellular matrix (Lipskaia et al., 2010). Changes in cellular calcium balance can cause a disruption of the extracellular matrix, which allows more efficient access of virus to the myocytes.
Logeart et al. (2001) injected various pharmacological agents 5 min before intracoronary delivery of adenoviral vectors encoding luciferase or β-galactosidase. Changes in the rabbits' coronary vascular permeability were studied with fluorescein isothiocyanate-labeled 70,000-Dalton Dextran. The results indicate that histamine, serotonin, and VEGF-165 pretreatment increased fluorescence by ∼100% in tissue samples of the targeted area versus no pretreatment group, whereas bradykinin did not increase sample fluorescence.
The role of numerous permeabilizing drugs was examined in a piglet model with intracoronary infusion of adenovirus or adeno-associated viral construct. The authors demonstrated that the best effect on the permeability of endothelial barrier was achieved with the administration of a phosphodiesterase-5 inhibitor and infusion of VEGF, nitroglycerin, and adenosine. The application of these agents allows for gene transduction in ∼80% of cells within the target zone (Sasano et al., 2007). An important observation was that the modulation of vascular permeability markedly reduced the virus exposure time required to obtain infection in greater than 90% of cardiac myocytes (Donahue et al., 1998). Currently more than 30 vasoactive substances affecting capillary permeability are known. Listed below are the main agents that have been used in cardiac gene therapy: 1. Histamine is a biologically active substance, chemically classified as an amine and stored primarily in the mast cells. It triggers the inflammatory response and has long been known as a factor contributing to microvascular permeability. Histamine-induced endothelial barrier dysfunction is caused by paracellular leakage from intercellular gaps. The study of Ikeda et al. (2002) evaluated the feasibility of restoring a δ-sarcoglycan in cardiomyopathic hamsters. It was demonstrated that using a solution with histamine undoubtedly increases microvascular permeability, allowing the achievement of homogeneous marker gene expression in ∼77% of left ventricular myocytes and helping to restore sarcoglycan in the myocyte membrane. 2. Serotonin is a monoactive neurotransmitter that is primarily found in the enterochromaffin cells of the gastrointestinal tract and in the central nervous system. It was suggested that increases in permeability induced by serotonin differ from those of other substances. The data show that serotonin can induce opening in venular endothelium (Michel and Kendall, 1997). The use of serotonin in a Langendorff model of rabbit hearts allows for more adenoviral transfection in cardiomyocytes in comparison with other agents (Donahue et al., 1998). These data are consistent with the view that serotonin pretreatment increases the permeability of the capillaries, resulting in enhanced attachment of viral vectors to myocardial cells (del Monte and Hajjar, 2003). 3. Bradykinin is a pharmacologically active nonapeptide kinin formed from kininigen by the action of kallikrein. Bradykinin works on blood vessels mostly through the release of prostacyclin and nitric oxide. It is a potent endothelium-dependent vasodilator. Permeabilization of the endothelium by bradykinin is accomplished by the formation of intercellular gaps, permitting the passage of macromolecules (Ehringer et al., 1996). The positive effect of bradykinin on permeability changes in the coronary vasculature after adenoviral delivery was also shown (Donahue et al., 1998). 4. Adenosine is an endogenous nucleoside occurring in all cells of the body. Intravenously administrated adenosine clears via cellular uptake by vascular endothelial cells and erythrocytes. Adenosine plays an important role in various biochemical processes including energy transfer and permeability properties (Jacobson et al., 2006). Stimulation of myocardial adenosine receptors enhances cardioprotection of preconditioning (Vogt et al., 1998). It has been suggested that injection of adenosine into the coronary arteries increases permeability and decreases heart rate during adenoviral transfer of phospholamban (Tsuji et al., 2009). Pretreatment of the animals with intracoronary delivery of adenosine was also used in other studies (Wright et al., 2001; del Monte and Hajjar, 2003; Hayase et al., 2005; Rengo et al., 2009). 5. Vascular endothelial growth factor (VEGF) is a glycoprotein originally identified as a vascular permeability factor due to its ability to increase microvascular permeability to plasma proteins (Bates and Curry, 1997). It was demonstrated that VEGF increases capillary and venular leakage as a result of opening endothelial intercellular junctions as well as induction of fenestration in the venular part of the endothelium (Roberts and Palade, 1995). Subsequently, its role in angiogenesis has been extensively studied (Tio et al., 1999). Delivery of human recombinant VEGF showed a three-fold increase in coronary vascular permeability that was assessed by Evans Blue accumulation (Wright et al., 2001). Using an ex vivo model of coronary perfusion in rabbits, it was found that a dose-response relationship between VEGF and the efficiency of adenoviral transfer to the heart existed. In addition, inhibitors of nitric oxide synthase and guanylate cyclase prevented this effect (Nagata et al., 2001). The increased transduction efficiency after VEGF gene transfer or coapplication of adeno-associated viral vector with VEGF was also demonstrated in other studies (Raake et al., 2008; White et al., 2011; Vera Janavel et al., 2012).
Cell-Penetrating Peptides
Most of the molecules entering cells via the receptor-mediated endocytosis pathway become trapped in endosomes and are ultimately destroyed in the lysosomes by numerous enzymes. A promising approach for overcoming this pathway utilizes cell-penetrating peptides (CPP), also known as protein transduction domains, which have the ability to move independently of membrane receptors and cell-type specificity. Current gene research focuses on two CPPs: Tat, originating from HIV type 1, and pAntp, isolated from the transcription factor Antennapedia. They can transport plasmid DNA, siRNA, liposomes, etc., across plasma membranes (Tang and Hammond, 2007; Koren and Torchilin, 2012).
Bian at al. (2007) engineered a protein consisting of the transcription factor GATA4 and the cell-penetrating protein VP22. Cardiac fibroblasts transfected with this construction were transplanted into the rat's infarct-border zone 1 month after LAD ligation. Animals that received treatment with CPP demonstrated increased left ventricular (LV) fractional shortening and reduced fibrosis of the infarcted myocardium (Bian et al., 2007). The enhanced transfection of the gene delivery using Tat-lipoplexes was demonstrated in a rat infarct model in vivo and in hypoxic cardiomyocytes in vitro (Ko et al., 2009).
Membrane Vesicles
In recent years, studies have led to the hypothesis that membrane vesicles (about 30–100 nm in size) secreted by a variety of cell types may represent a novel method of intracellular communication and exchange of genetic information. Exosomes, a type of membrane vesicle, release into the extracellular environment upon fusion of multivesicular endosomes with the cell surface (Fevrier and Raposo, 2004). Taking into account their nanoparticle size, exosomes can be utilized as a novel delivery platform for gene therapy that can transport various biological substances ranging from proteins to mRNA and miRNA. It has been demonstrated that exosomes can be used as vehicles for delivering siRNA and miRNA that can inhibit cancer cell growth and proliferation (Tan et al., 2012).
The therapeutic potential of exosomes as transport vehicles was shown in mice neuronal cells. The authors targeted the BACE1 enzyme (essential for the generation of β-amyloid, which is important to stopping the progression of Alzheimer's disease). It was confirmed that exosome-mediated siRNA delivery can cross the blood–brain barrier, thus controlling BACE1 expression levels (Alvarez-Erviti et al., 2011). Although we did not find studies that use membrane vesicles for cardiac gene transfer, undoubtedly the discovery of exosomes has opened a completely new paradigm for gene therapy potential in the treatment of heart diseases.
Conclusions
1. Enhancement of myocardial gene transfer is not possible without an understanding of existing biological barriers. Depending on the route of delivery (transvascular or intramyocardial), one should take into consideration the need to overcome the various barriers.
2. Development of an optimal gene delivery technology depends on the molecular mechanisms of intracellular trafficking, transcription, and the translation of viral and nonviral gene vectors.
3. Among the physical and mechanical methods of improving the cellular permeability, the greatest effect is achieved by using electro- and sonoporation. Study of the other methods requires additional research.
4. The optimal, clinically translatable technique for cardiac gene transfer probably should incorporate ischemic preconditioning, creation of a sophisticated minimally invasive “closed-loop“ recirculatory system, a coronary sinus route of gene injection with flow and pressure adjustment, application of different pharmacological mediators, cell-penetrating peptides, and membrane vesicles.
Footnotes
Acknowledgments
We thank Anne Olson for the excellent illustrations in this article. We are grateful to Richard Hillesheim, BS, and Richard D. Williams, BS, for editorial assistance. This work was supported by grants from the National Heart, Lung and Blood Institute, NIH 1-R01-HL083078-01A2, and the Gene Therapy Resource Program of the National Heart, Lung and Blood Institute, NIH P30-DK047757.
Author Disclosure Statement
No competing financial interests exist.