
Abstract
Corneal transplantation (keratoplasty) is the most common type of tissue replacement in the world. The increased rate of graft rejection after keratoplasty is a central problem for repeated transplantations and in inflamed host corneas. It has been shown that apoptosis of grafted epithelium has a role in corneal allograft rejection. This study focused on the T-cell response triggered in BALB/c mice after allogeneic corneal transplantation with and without anti-apoptotic p35-transduced epithelium. To restrict p35 expression to the epithelial cells, modified allogeneic composite grafts were created. As a result, it was found that the proportion of alloreactive CD4+ T cells in postoperatively removed cervical lymph nodes was reduced in the p35-transduced group compared to the allogeneic control group. Diminished priming of the CD4+ T cells was supported by significantly decreased proliferation and lower interferon gamma secretion when compared to allogeneic engraftments. The reduced priming of CD4+ lymphocytes is the first confirmation of the functionality of p35 in the epithelium of corneal grafts to alter the development of the recipient's immune response. Thus, modification of allosensibilization seems to be a promising tool for reducing graft-mediated immune response following corneal transplantation.
Introduction
T
In this study, it was hypothesized that overexpression of the anti-apoptotic p35 protein in corneal epithelial sheets reduces the priming of T cells in draining lymph nodes (LNs) after transplantation. It was assumed that inhibition of apoptosis with anti-inflammatory effects promotes the persistence of mechanisms regulating the immune-privileged status of the cornea, ultimately resulting in an increase of graft survival.
Materials and Methods
Animals
Eight- to twelve-week-old male C57BL/6 (B6; n = 15) and B/c (n = 26) mice were obtained from Harlan Laboratories. The mice were hosted in the Central Animal Laboratory, University Duisburg-Essen, and treated according to guidelines established by the Association for Research in Vision and Ophthalmology (ARVO) Statement for the Use of Animals in Ophthalmic and Vision Research. Anesthesia was performed subcutaneously using ketamine and xylazine at a dose of 120 mg/kg and 20 mg/kg of body weight, respectively.
Corneal transduction and preparation of composite graft
The following donor corneas were utilized: B/c epithelium (EPI) + B/c stroma-endothelium (S-E; n = 5); B6 EPI + B6 S-E (n = 5); B6 EPI expressing the empty vector + B6 S-E (n = 4); and B6 EPI expressing p35 + B6 S-E (n = 4). B/c was used as a host. In addition, naive B/c mice were enrolled as surgically untreated controls. Donor corneas with a diameter of 2.0 mm were excised from naive B/c and B6 mice and transduced either empty pHAGE-CMV-IZsGreenW vector or p35-carrying pHAGE-CMV-p35-IZsGreenW (Harvard Gene Therapy Initiative). After determination of best transduction conditions, vectors were utilized at a concentration of 3 × 105 IU/mL applied in Roswell Park Memorial Institute (RPMI) medium (Thermo Fisher Scientific, Carlsbad, CA) containing 5% fetal calf serum (FCS; PAA, Linz, Austria) and hexadimethrine bromide (8 μg/mL; Sigma–Aldrich, St. Louis MO) at 37°C for 1 h. The transduced corneas were incubated in 20 mM of ethylenediaminetetraacetic acid (Thermo Fisher Scientific) for 45 min, and thereafter the epithelium was removed and placed immediately on S-E obtained from the same strain. All prepared composite donor corneas were left to dry for 1 min to achieve better epithelial sheet attachment. The tissues were preserved in Optisol-GS medium (Bausch & Lomb, Irvine, CA) at 4°C, and the transplantations were performed within 24 h. Before surgery, the fixation of epithelium onto the stroma was ensured visually using a microscope.
Confocal microscopy of transduced epithelium
For the transduction experiment, treated and untreated corneas were subjected to confocal microscopy. Corneas were fixed in 3.7% paraformaldehyde for 10 min and stained with DAPI (1 μg/mL; Sigma–Aldrich) for 10 min. The samples were mounted with VECTASHIELD® mounting medium (Vector Laboratories, Inc., Burlingame, CA) and sealed with clear nail polish. The slides were stored in 4°C and analyzed utilizing a Leica DM IRE2 confocal fluorescence microscope (Leica Microsystems, Wetzlar, Germany).
Transplantation and assessment of composite grafts
The cornea was trephined with a diameter of 1.5 mm in recipient mice and removed. The composite button with a diameter of 2.0 mm was placed on graft bed and sutured with interrupted 11-0 nylon sutures (Ethicon, Somerville, NJ). The temporary lid closure with nylon sutures for 2 days was carried out to protect the grafted cornea. Corneal sutures were removed at day 8 after transplantation. Transplanted corneas were observed according to published standardized opacity grading score: 0, clear graft; 1+, minimal superficial non-stromal opacity; 2+, minimal deep stromal opacity with the pupil margin and iris vessels visible; 3+, moderate deep stromal opacity with only the pupil margin visible; 4+, intense deep stromal opacity with the anterior chamber visible; and 5+, maximum stromal opacity with total obscuration of the anterior chamber. 17 According to the opacity scores, the cumulative survival of grafts was analyzed using the Kaplan–Meier method. To differentiate changes between transplantation groups, failure was defined as an opacity score of >3.
Cells counts and characterization of T-cell subsets in LNs
Three weeks after transplantation, two ipsilateral cervical LNs of every treated animal were removed, and single-cell suspensions were prepared in RPMI medium supplemented with 10% FCS. The red blood cells were lysed osmotically, and viable cells were determined using trypan blue (Thermo Fisher Scientific) exclusion assay in the Neubauer chamber (Glaswarenfabrik Karl Hecht, Sondheim, Germany). The Fc-receptor blockage was performed with anti-mouse CD16/CD32 antibody (clone: 93; eBioscience, San Jose, CA) and isolated lymphocytes were stained with the following antibodies: anti-mouse CD4-FITC (clone: GK1.5; 2.5 μg/mL), CD8-FITC (clone: 53–6.7; 5 μg/mL), CD25-PE (clone: PC61.5; 1.25 μg/mL), CD69-PE (clone: H1.2F3; 2.5 μg/mL), and Foxp3-PECy5 (clone: FJK-16S; 10 μg/mL; eBioscience) according to the manufacturer's instructions. Viable and death cells were discriminated by propidium iodide (PI; Sigma–Aldrich) and analyzed on a FACSCalibur flow cytometer with the CellQuest Pro software (BD Biosciences, Franklin Lakes, NJ).
Mixed lymphocyte reaction and bromodeoxyuridine proliferation assay
Single-cell suspensions from cervical LNs of the recipient and naive B/c were prepared as described above. The CD90.2+ cells were separated via positive selection using CD90.2 magnetic microbeads (Miltenyi Biotec, Bergisch Gladbach, Germany). Naive B6 spleen was removed, and the erythrocytes were lysed. Thereafter, the CD90.2– splenocytes were collected via depletion by CD90.2 magnetic microbeads. The purity (>90%) of CD90.2+ and CD90.2– cells was confirmed by anti-mouse CD90.2-FITC antibody (clone: 30-H12; Miltenyi Biotec) with flow cytometry. Viable T and non-T cells were enumerated in hemocytometer by trypan blue exclusion assay and diluted to a concentration of 106 cells per 1 mL in RPMI medium with 10% FCS for subsequent mixed lymphocyte reaction (MLR). Kinetics of MLR-induced cell proliferation was measured at 24, 48, 72, and 96 h using ratios 1:1, 1:5, and 5:1 of co-cultured naive B/c CD90.2+ and naive B6 CD90.2– cells according to the manufacturer's instructions using the bromodeoxyuridine (BrdU) proliferation assay (Roche, Basel, Switzerland; Millipore, Billerica, MA). Briefly, the labeled BrdU was detected by anti-BrdU monoclonal antibody and incubated with anti-mouse peroxidase conjugated secondary antibody. Thereafter, tetramethylbenzidine (TMB) solution was added, and the reaction was stopped with sulfuric acid (H2SO4; Carl Roth, Karlsruhe, Germany). Optical density (OD) was measured using the Sunrise reader (450 nm; Tecan, Männedorf Switzerland). Subsequently, MLR-induced proliferation was performed using recipient CD90.2+ and naive B6 CD90.2– splenocytes at a ratio of 1:1 for 96 h.
Flow cytometry
The MLR cultures of all study groups were collected after 96 h of cultivation, and the Fc-receptor binding sites were blocked with anti-mouse CD16/CD32 antibodies. Next, CD4+ cells were characterized with the anti-mouse CD4-FITC, CD25-PE, CD69-PE, and Foxp3-PECy5 antibodies using a FACSCalibur flow cytometer with CellQuest Pro software. PI-positive dead cells were excluded from the determinations.
Enzyme-linked immunosorbent assay
To measure the cytokine response in MLR cultures, sandwich enzyme-linked immunosorbent assay (ELISA) was employed. Supernatants were collected after 96 h, and the mouse interferon gamma (IFN-γ) and IL-10 ELISA reagent sets (eBioscience) were enrolled according to the manufacturer's instructions. Briefly, the capture antibody was detected with anti-mouse IFN-γ and IL-10 antibody, both conjugated with biotin, and detected with horseradish peroxidase–coupled avidin. Then TMB solution was utilized, and the incubation was stopped with H2SO4. The microplate absorbance reader was applied for measurement of OD at 450 nm. The recombinant protein of IFN-γ and IL-10 (eBioscience) was used for generating standard curve of respective cytokine based on the manufacturer's protocol.
Detection and measurement of induced apoptosis
In addition to transplantations of p35-modulated composite grafts in animal model, the anti-apoptotic effect of p35 was tested in murine corneal epithelial cells. Murine corneas were transduced with either empty pHAGE-CMV-IZsGreenW vector or p35-carrying pHAGE-CMV-p35-IZsGreenW (3 × 105 IU/mL) according to the protocol described in the section “Corneal transduction and preparation of composite graft.” Apoptosis was induced using the topoisomerase II inhibitor Etoposide at 3 ng/mL for 6 h, provoking premature cell death via the intrinsic apoptotic pathway by specific stimuli. To detect apoptosis, terminal deoxyribonucleotidyl-transferase (TdT)-mediated deoxyuridine-5′-triphosphate-digoxigenin (dUTP) nick-end labeling (TUNEL) assay (Roche) was used according to the manufacturer's instruction to measure DNA breaks in epithelial cells within murine corneas. Vital nuclei were visualized using To-Pro 3 iodide (Thermo Fisher Scientific). Corneas were washed and mounted with VECTASHIELD® mounting medium for fluorescence. IZsGreen expression and TUNEL positivity were detected by a Leica TSC-SP2 confocal laser scanning microscope (Leica Microsystems).
Statistical analysis
Data are presented as the mean ± standard deviation (SD). p-Values for statistical analyses were obtained with a two-tailed t-test using by SPSS Statistics for Windows v24.0 (IBM Corp., Armonk, NY). A p-value of <0.05 was considered statistically significant. The Kaplan–Meier method was used to evaluate cumulative probability of graft survival.
Results
Methodical setup of composite grafts and transduction efficacy
First, the B6 EPI p35 + B6 S-E composite graft was successfully prepared (Fig. 1a and b). The transduction and expression of the baculovirus encoded anti-apoptotic gene p35 was indirectly confirmed by evaluating the expression of fluorescent reporter protein IZsGreenW, as both genes are coordinately transferred. The epithelium was analyzed by confocal fluorescence microscopy to confirm the presence of IZsGreenW (Fig. 1c). Figure 1d demonstrates the postoperative appearance of transplanted allogeneic B6 EPI p35 + B6 S-E composite graft in B/c at day 0. All transplantation groups are graphed in Fig. 1e.
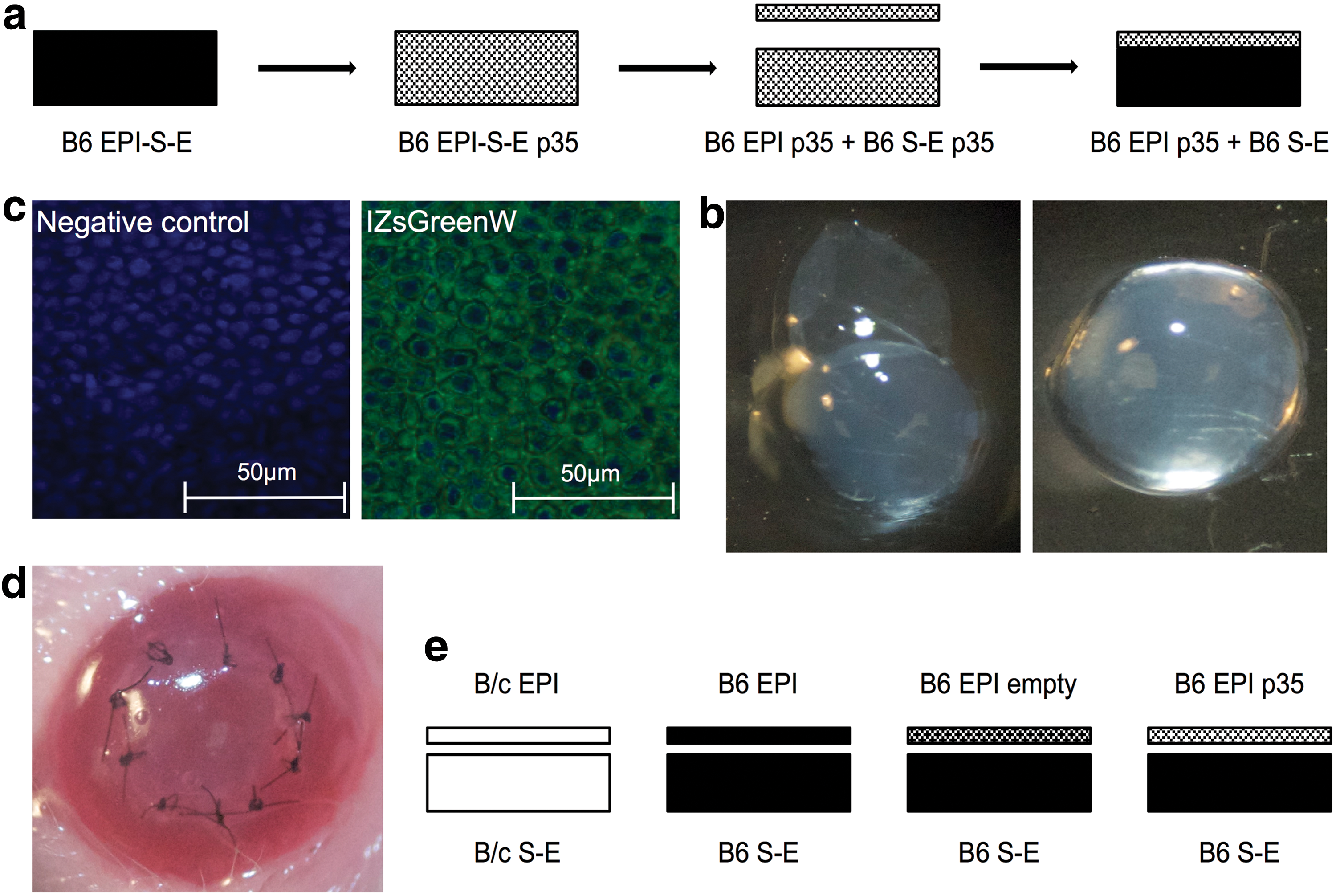
The preparation of B6 epithelium (EPI) p35 + B6 stroma-endothelium (S-E) composite graft and transplantation model. Harvested B6 corneas were incubated with pHAGE-CMV-p35-IZsGreenW lentiviral vector (3 × 105 IU/mL) in the presence of hexadimethrine bromide for 1 h. Transduced corneal layers were treated with ethylenediaminetetraacetic acid for 45 min, and subsequently the intact sheet of epithelium was peeled off and transplanted on untreated S-E of B6
Post-transplantation status of composite grafts
During the monitoring of the transplanted corneas, the opacity grading scores described above were applied. Three weeks after transplantation of B6 EPI p35 into B6 S-E and of B/c EPI into B/c S-E groups, an average score was measured (moderate deep stromal opacity) of 3.3 ± 0.5 and 2.8 ± 0.4, respectively, while the allogeneic transplants B6 EPI into B6 S-E and B6 EPI empty into B6 S-E showed a higher average score (intense deep stromal opacity) of 3.6 ± 0.5 and 3.8 ± 0.5, respectively (Table 1). In addition, Fig. 2a demonstrates opacity scores of syngeneic and allogeneic corneal transplantations during the follow-up time (2–21 days). The average opacity grading score was significantly lower in syngeneic group compared to allogeneic engraftments, except for p35-modulated transplantations. Based on opacity scores, the graft failure was less observed in p35-treated mice compared to other allogeneic transplantations in a Kaplan–Meier survival analysis (Fig. 2b).
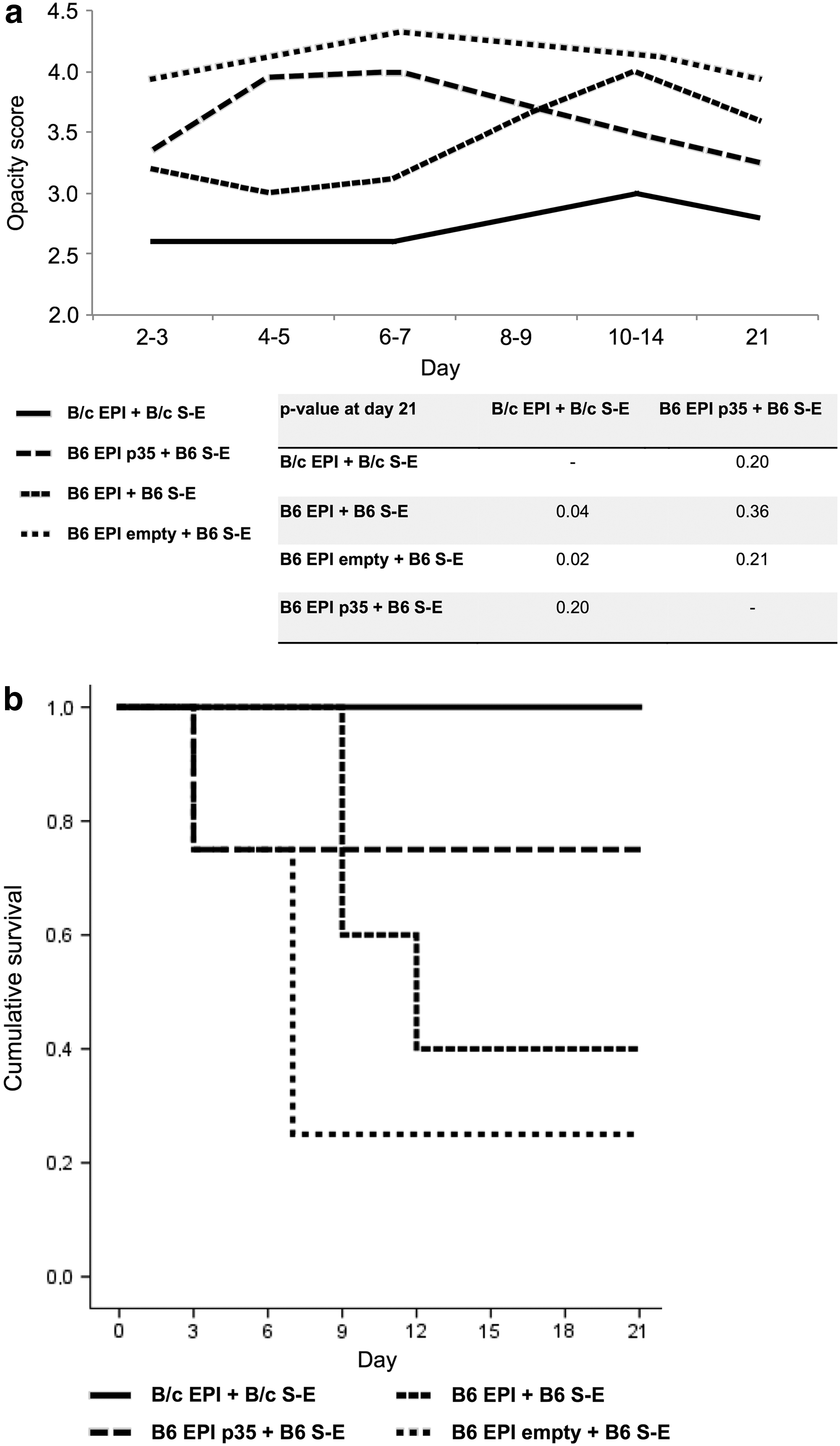
Opacity of the grafted cornea as an indicator for graft survival. Transplanted composite grafts of B/c EPI + B/c S-E (n = 5), B6 EPI + B6 S-E (n = 5), B6 EPI empty + B6 S-E (n = 4), and B6 EPI p35 + B6 S-E (n = 4) were studied for opacity according to published standardized opacity grading score: 0, clear graft; 1+, minimal superficial non-stromal opacity; 2+, minimal deep stromal opacity with the pupil margin and iris vessels visible; 3+, moderate deep stromal opacity with only the pupil margin visible; 4+, intense deep stromal opacity with the anterior chamber visible; and 5+, maximum stromal opacity with total obscuration of the anterior chamber. The score was estimated for each composite graft, and the average opacity scores of transplantation groups were represented as a curve during the 3-week follow-up time. The score was significantly lower in the syngeneic group compared to B6 EPI + B6 S-E and B6 EPI empty + B6 S-E transplantations (p = 0.04 and p = 0.02, respectively), except for B6 EPI p35 + B6 S-E engraftments (p = 0.20)
The post-transplantation corneal and lymph node measurements 3 weeks after surgery of treated animals and naive B/c mice
p-Values were calculated (two-tailed t-test) in comparison with B6 EPI p35 + B6 S-E.
B/c, Balb/c; EPI, epithelium; S-E, stroma-endothelium; SD, standard deviation; Tregs, regulatory T cells.
Cell count and T-cell phenotyping in postoperatively removed LNs
Based on the assessment of corneal status, it was presumed that the total leukocyte count in cervical LNs was increased in allogeneic transplants with rejection signs. Cervical LNs of mice were removed, and the mean total number of viable leukocytes per two ipsilateral cervical LNs was counted (Table 1). The mean total number of cells did not differ significantly between B6 EPI p35 + B6 S-E and the other allogeneic transplantations. Determination of the T-cell subtypes in isolates of naive B/c and the different allogeneic and syngeneic transplanted mice revealed no alterations with respect to the CD4+ (Table 1) and CD8+ cell fractions (data not shown). Additionally, early and late activation markers in CD4+ cells, namely CD69 and CD25, respectively, and the amount of regulatory T cells (Tregs) were determined. The percentage of CD69, CD25+ leukocytes, and Tregs did not alter within analyzed study groups (Table 1). Thus, the post-transplantation CD4+ and CD8+ fractions without allogeneic stimulation did not reveal any differences with respect to T-cell frequencies between investigated naive B/c, allogeneic, and syngeneic groups.
MLR-induced cell proliferation
To determine the optimal conditions for subsequent studies with respect to incubation time and the ratio of co-cultivated cells, the kinetics of T-cell proliferation were studied using the BrdU approach. Magnetically separated naive B/c CD90.2+ cells from cervical LNs and naive B6 CD90.2– splenocytes were co-cultivated at 1:1, 1:5, and 5:1 ratios for 24, 48, 72, and 96 h. Highest induction of T-cell proliferation was observed at 96 h MLR using the 1:1 ratio (Fig. 3a). Interestingly, nearly no differences in cell proliferation were observed when the ratio of stimulated and stimulating cells was 1:5 and 5:1 (Fig. 3a). Next, MLR was used as an alloantigen recall assay to assess the T-cell priming capability after transplantation. Therefore, magnetically separated CD90.2+ cells from cervical LNs of all recipients and naive B/c with naive B6 splenocytes were stimulated at a 1:1 ratio for 96 h. In comparison with allogeneic transplantations, the B6 EPI p35 + B6 S-E group showed significant lower proliferation. Its proliferation was nearly as low as seen in the naive and syngeneic transplantation groups (Fig. 3b). In addition, allogeneic B6 EPI + B6 S-E and B6 EPI empty + B6 S-E had equivalent proliferation capability (Fig. 3b).
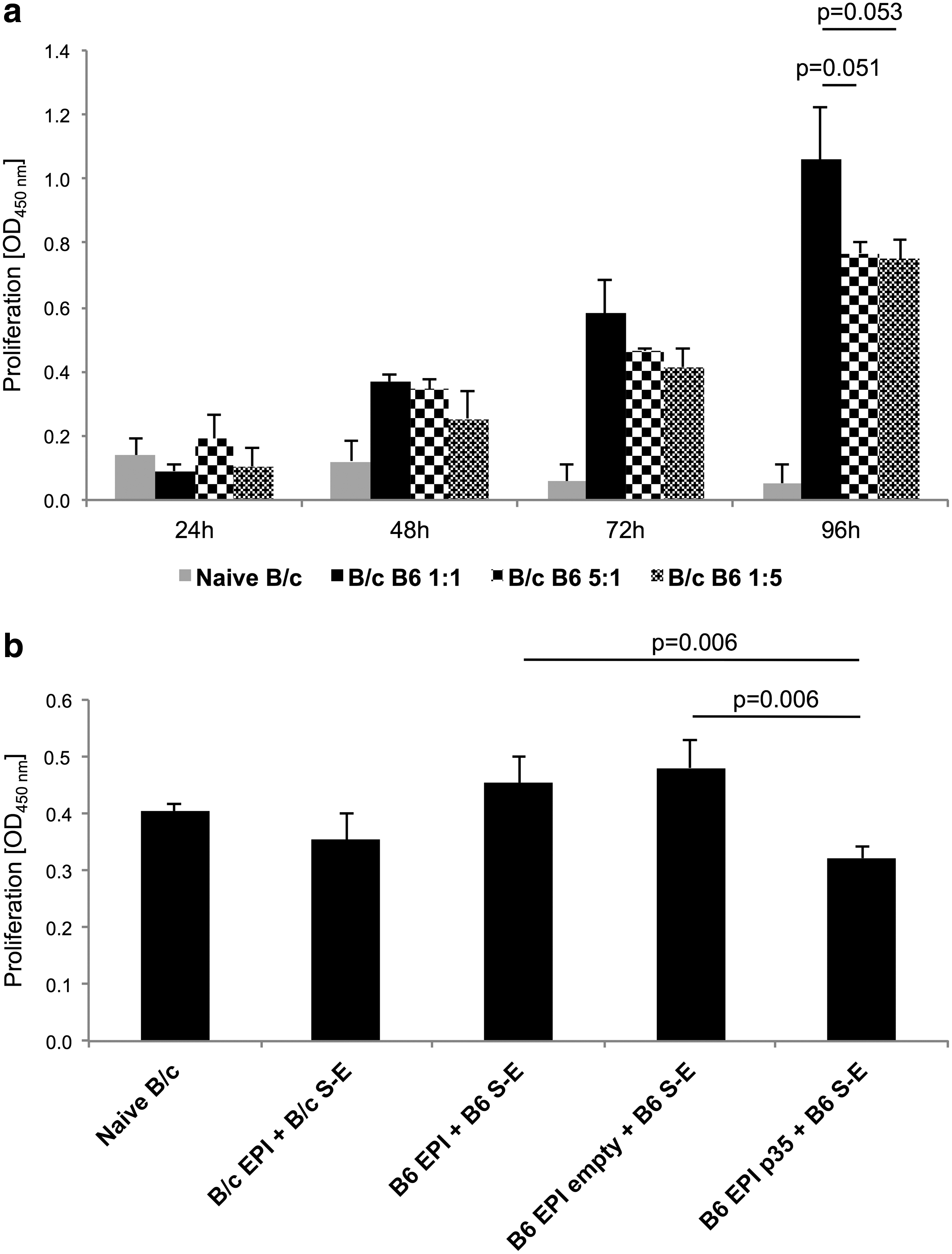
The kinetics and post-transplantation proliferation tests in MLR cultures. Naive B/c T cells from cervical lymph nodes (LNs) were stimulated with B6 splenocytes in different stimulatory ratios (1:1, 1:5, and 5:1) for up to 96 h. The bromodeoxyuridine (BrdU) proliferation assay was performed at 24, 48, 72, and 96 h by colorimetric enzyme-linked immunosorbent assay (ELISA) to determine the proliferation kinetics after co-culture of B/c CD90.2+ and B6 CD90.2– cells. The strongest stimulatory effect, although not statistically significant compared to ratios 1:5 and 5:1 (B/c: B6), was measured at a ratio of 1:1 (B/c:B6) at 96 h
Phenotyping CD4+ cells in MLR cultures
Next, the fraction of viable CD4+ cell subsets in 96 h MLR cultures were determined utilizing flow cytometry. The total proportion of CD4+ cells in isolated naive B/c, B/c EPI + B/c S-E, and B6 EPI p35 + B6 S-E cell cultures was 50%, 51%, and 52%, respectively, while in B6 EPI + B6 S-E and B6 EPI empty + B6 S-E groups, the frequency of CD4+ cells increased to 63% and 66%, respectively (Fig. 4a). The proportion of Tregs just slightly increased in the case of B6 EPI + B6 S-E, whereas the Tregs numbers of the other treatments remained comparable to the status before MLR. The measured Tregs frequency was: naive B/c 4%, B/c EPI + B/c S-E 3%, B6 EPI + B6 S-E 10%, B6 EPI empty + B6 S-E 2%, and B6 EPI p35 + B6 S-E 3% (Fig. 4b). Concomitantly, during evaluation of CD4+ cells, the cell surface expression of CD69 and CD25, as well established markers for activation, was determined. In CD4+ cells, fairly similar amounts of CD69+ cells were observed in non-transplanted (17%), syngeneic (13%), as well as allogeneic transplanted mice (B6 EPI + B6 S-E 13%, B6 EPI empty + B6 S-E 16%). However, in CD4+ cells of B6 EPI p35 + B6 S-E, the amount of CD69+ cells decreased to 6% (Fig. 4c). The frequency of CD25+ also decreased on CD4+ cells of the B6 EPI p35 + B6 S-E–treated animals. Here, flow cytometric analysis confirmed the presence of CD4+CD25+ on the cells isolated from MLR cultures as follows: naive B/c 12%, B/c EPI + B/c S-E 12%, B6 EPI + B6 S-E 18%, B6 EPI empty + B6 S-E 14%, and B6 EPI p35 + B6 S-E 7% (Fig. 4d). In addition to decreased proliferation, the p35 transduction in composite grafts demonstrated the reduced priming capacity of CD4+ cells on the basis of cell phenotype, leading to a reduction of CD4+ cells and lower expression of CD69 and CD25 in the cell membrane if compared to other allogeneic transplantations.
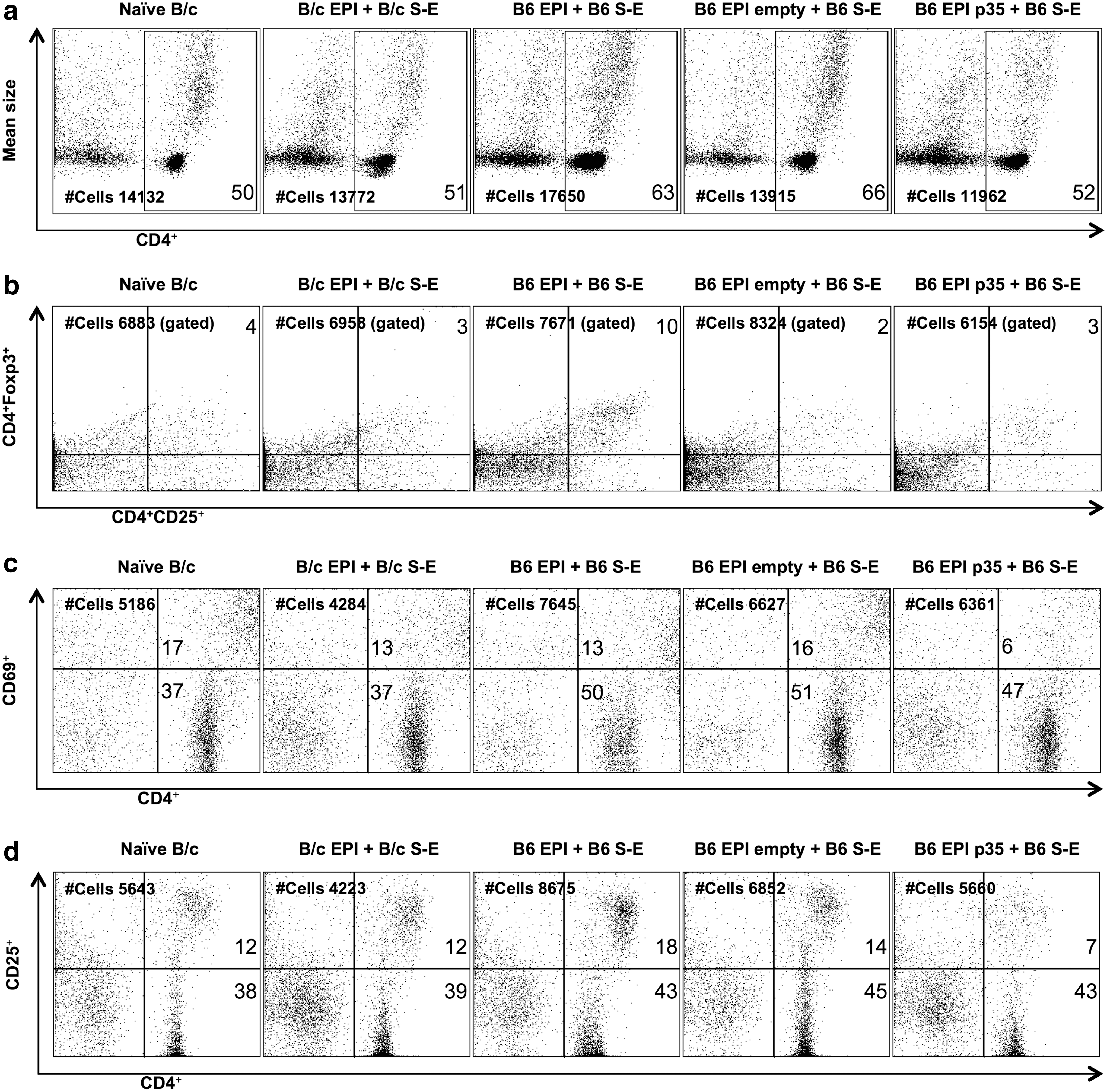
The phenotype of CD4 subpopulations in mixed lymphocyte reaction (MLR) cultures. Three weeks after transplantation, single-cell suspensions were prepared from ipsilateral cervical LNs of B/c EPI + B/c S-E (n = 5), B6 EPI + B6 S-E (n = 5), B6 EPI empty + B6 S-E (n = 4), B6 EPI p35 + B6 S-E (n = 4), and from naive B/c mice. Thereafter, magnetically sorted T cells were co-cultured with CD90.2– cells from naive B6 spleen for 96 h. Subsequently, anti-mouse CD4-FITC, CD25-PE, CD69-PE, and Foxp3-PECy5 antibodies were used for staining of MLR cultures. Viable cells (PI negative) were gated and CD4 subpopulations were analyzed by flow cytometry. The frequency (%) of CD4+ cells in the allogeneic p35-transduced group (52%) demonstrated a diminished proportion of cells compared to the B6 EPI + B6 S-E (63%) and B6 EPI empty + B6 S-E (66%) groups (
Cytokine secretion
The immunomodulatory cytokine IFN-γ plays a central role in establishing alloimmunity. 18 On the other hand, IL-10 is one of the major inhibitory cytokines that influences the functions of Tregs. 19 Four days after stimulation with B6 alloantigen, significantly higher IFN-γ concentrations were found in the MLR supernatants of allogeneic genetically unmodified composite transplantations compared to allogeneic p35 transplantations (Fig. 5a). The post-MLR IFN-γ level in isogeneic transplantations and naive B/c used as control groups was analogous to the B6 EPI p35 + B6 S-E group (Fig. 5a). The lower concentration of IFN-γ in the p35-transduced group supported the outcomes of proliferation analysis and flow cytometric evaluation of CD4+ cells. However, B6 stimulation with splenocytes for 96 h did not show significant differences in the IL-10 concentrations in the study groups analyzed (Fig. 5b).
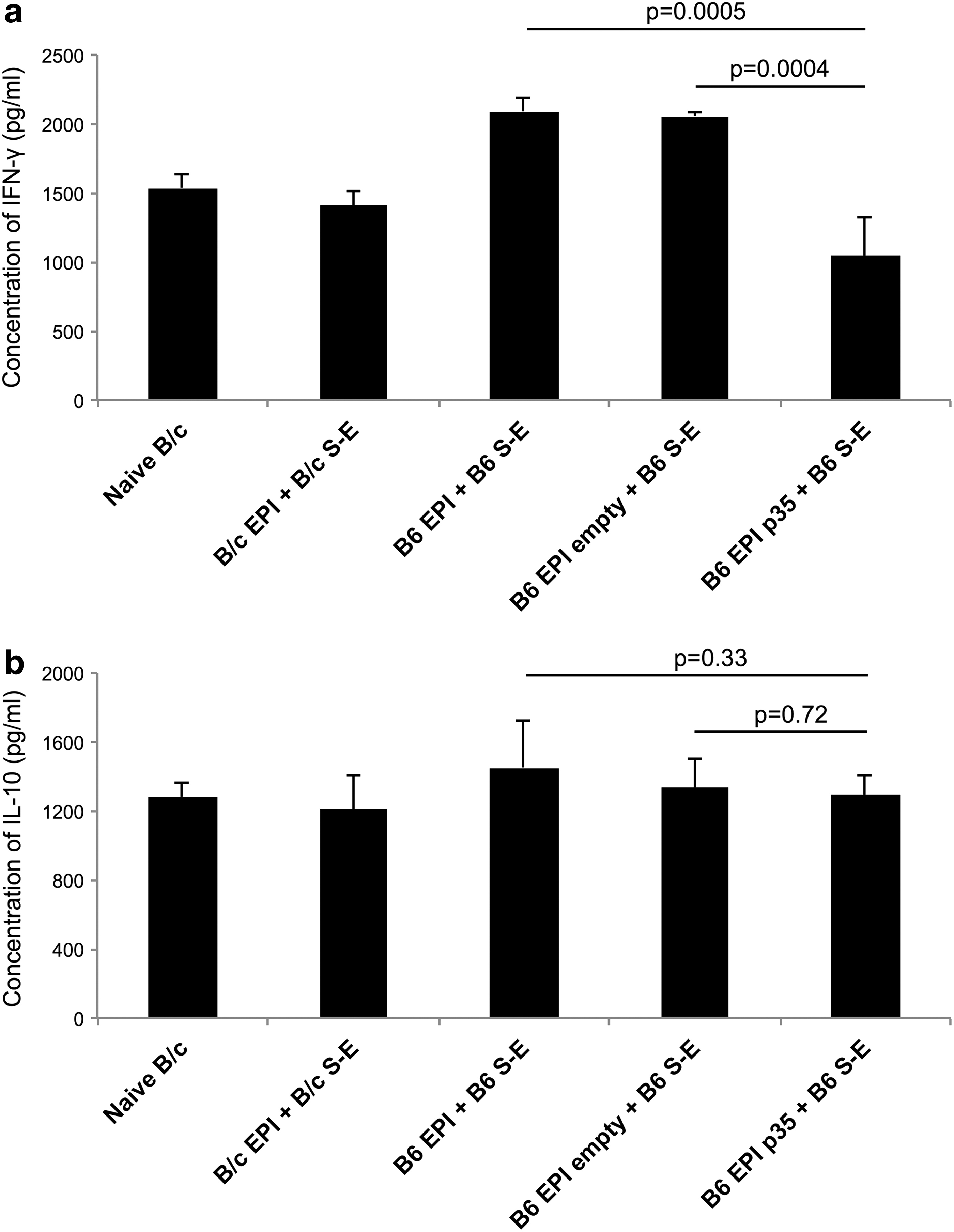
The concentrations of interferon (IFN)-γ and interleukin (IL)-10 in MLR cultures. Cells from cervical LNs were isolated 3 weeks after transplantation of B/c EPI + B/c S-E (n = 5), B6 EPI + B6 S-E (n = 5), B6 EPI empty + B6 S-E (n = 4), B6 EPI p35 + B6 S-E (n = 4), and naive B/c. The CD90.2+ cells were magnetically sorted and stimulated with naive B6 splenocytes as an alloantigen recall assay for 96 h. The ELISA was employed for MLR cultures to detect IFN-γ
Anti-apoptotic effect in corneal epithelial cells
The p35 overexpression protected murine corneal epithelium against programmed cell death induced by etoposide. The confocal laser scanning microscope images demonstrated significantly less TUNEL-positive cells in p35-transduced epithelial cells compared to epithelial cells expressing IZsGreenW only (Fig. 6). In addition to a confirmed reduction of T-cell priming in LNs modulated by p35, the results showed the intracellular functionality of p35 to downregulate apoptosis in corneal epithelial cells.
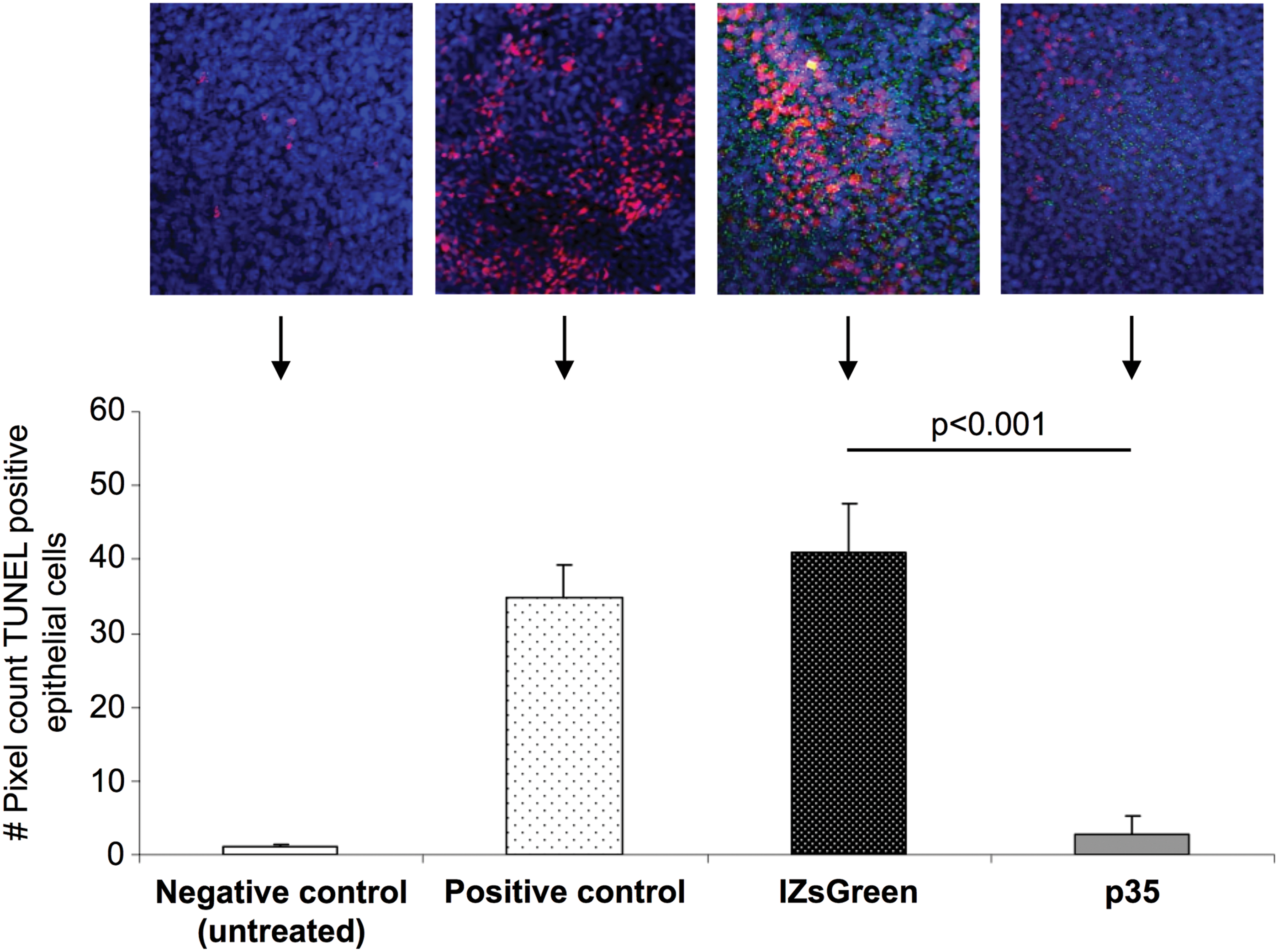
Anti-apoptotic protein expression in corneal epithelium leads to protection against a strong apoptotic inducer. To determine whether overexpression of p35 results in a protection of corneal epithelium against apoptosis, murine corneal epithelial cells were transduced with empty pHAGE-CMV-IZsGreenW vector and p35-carrying pHAGE-CMV-p35-IZsGreenW (3 × 105 IU/mL) followed by treatment with etoposide at 3 ng/mL for 6 h. Vital nuclei were visualized using To-Pro 3 iodide, and immunohistochemical evidence for DNA strand breaks was obtained by identifying terminal deoxyribonucleotidyl-transferase-mediated deoxyuridine-5′-triphosphate-digoxigenin nick-end labeling positivity in confocal laser scanning microscope (40 × ). Epithelial cells expressing p35 showed significantly less apoptosis compared to epithelial cells expressing IZsGreenW only (p < 0.001). The significance was verified with a two-tailed t-test. Color images available online at
Discussion
Rejection of the transplanted cornea is triggered by CD4+ cells and is associated with developing DTH to alloantigens. 17,20 Earlier studies by Fuchsluger et al. showed the effect of p35 protein in corneal epithelium on decreasing post-transplantation DTH, thus improving transplant survival significantly 7 weeks after surgery in a mouse model. 14 However, the allosensitization modulated by p35 on the cellular level in cervical LNs has remained unclear. The current study transplanted mouse donor corneas transduced with the p35 and analyzed various immunological aspects. Corneal epithelium was used, as it is a primary trigger of alloimmunogenicity. 11,21 The results confirmed the hypothesis that donor epithelial transduction with p35 decreases later allosensitization of the recipient by decreased priming of CD4+ cells per se in LNs in comparison with other allogeneic transplantations. To the authors' knowledge, the alteration of T-cell priming due to p35 transduction after tissue transplantation has not been investigated before. The allogeneic corneal graft survival was significantly improved using gene therapy with the anti-apoptotic gene Bcl-xL, where the treatment was directed to endothelial cells to make the transplant more “resistant” and avoid graft failure. 22 Thus, previously described Bcl-xL treatment was affecting the efferent loop of the alloimmunity when p35 was dealing with allorecognition in the donor epithelium.
In spite of the fact that LCs with MHC class II molecules expressing capability have been identified in the cornea, the prevailing opinion is that the initiation of the post-transplantation rejection mainly occurs in an indirect way where the “uptake” of alloantigens occurs via recipient APCs. 23,24 Thus, the migration of inflammatory cells into the graft and pericorneal angiogenesis of draining vessels is crucial for initiating the immune response.
In the present study, 3 weeks after surgery, the average opacity grading score was ≥2 in all transplantation groups. According to previously published data, 17 all the grafts were considered to have a “rejection reaction” not a “graft failure.” The opacity grading score described above was developed for a conventional engraftment, and therefore composite corneal transplantations may demonstrate higher opacity scores 3 weeks after surgery. As p35-modulated grafts did not demonstrate significantly higher opacity scores (indicators of graft survival) compared to syngeneic transplantations, this contributes to the postulated hypothesis of reduced allorecognition.
The cellular composition of ipsilateral neck LNs removed 3 weeks after transplantation was not different with respect to T-cell subpopulations between the naive and different forms of transplantations. It should be mentioned that the mean total number of cells per removed two LNs was increased in surgically manipulated mice. This could be explained by the ongoing immune responses, since cervical LNs are the first site of sensibilization after transplantation. 25 The priming of T cells in LNs includes expression of activation markers, cytokine secretion, and proliferation. 26 MLR was used for the assessment of allosensibilization to study T-cell activation. When measuring the MLR outputs, the analysis of BrdU-monitored proliferation kinetics provided the best result at a 1:1 ratio of CD4+ cells and stimulatory cells with a 96 h incubation time. Post-transplantation alloantigen recall assay demonstrated a decreased proportion of CD4+ cells in the p35-transduced group compared to other transplantations. The expression of early (CD69) and late (CD25) markers after MLR was reduced on the surface of CD4+ cells isolated from genetically modified animals compared to simultaneously transplanted allogeneic groups. The low activity of Tregs in MLR was expected in non-activated T cells, with the exception of the allogeneic empty vector group where the proliferation of CD4+ cells was confirmed by BrdU assay. In addition, reduced allosensibilization in the p35 group was also supported by decreased IFN-γ concentration and reduced proliferation of the cells in BrdU analysis. These results support the argument that corneal epithelium is one of the sources of allosensibilization, and that p35 exerts a suppressive effect on immune reaction in case of intraepithelial expression. The exact mechanisms between p35-expression in corneal epithelial cells and reduced priming of T cells in host cervical LNs are not proven. Additionally, the effect of p35 expression is defined because of the limited life-span of corneal epithelial cells. After keratoplasty, donor-derived epithelium will be replaced over time with epithelial cells of the recipient.
In order to describe the potential mechanisms of p35 effects on epithelium, the multifunctionality of p35 must be considered. First of all, p35 is known as an anti-apoptotic protein, as confirmed in this study, and its overexpression inhibits caspases that are the key mediators of the apoptotic response. 27,28 In mammals, p35 is capable of inhibiting initiator caspases, specifically caspase-1, -8, and -10, and executioner caspases-3, -6, and -7. 29 Due to inhibited caspases, p35 is effective in affecting the inhibition of apoptosis induced via the mitochondrial as well as via tumor necrosis factor (TNF) and Fas-pathways. It has been demonstrated that p35 is also capable of inhibiting nitric oxide–mediated apoptosis in mouse macrophages by decreasing the release of cytochrome C from mitochondria. 30 Furthermore, p35 might function as an antioxidant by scavenging reactive oxygen species (ROS) that are produced in case of inflammation and cause ROS-mediated apoptosis. 31 Moreover, it has previously been shown that p35 expression in corneal endothelial cells leads to significant reduction of cell death via inhibiting the intrinsic and extrinsic apoptotic pathways. 32
With regard to the anti-inflammatory effects of p35, the inhibition of the proteolytic activity of ICE (caspase-1) should be stated. 27 Specifically, pro-inflammatory cytokine IL-1β that belongs to the superfamily of IL −1 has been synthesized from biologically inactive precursor, pro-IL-1β, which has to undergo cleavage by caspase-1 in order to attain functional activity. 33 The corneal epithelium has the ability to express IL-1β constantly, 34 –36 while the continuous expression of the IL-1 receptor occurs in corneal stromal and epithelial cells. 37,38 Due to inflammatory stimuli, the migration of limbal LCs into the cornea is associated with secretion of IL-1β. 39 Moreover, if an iatrogenically induced inflamed B/c cornea was treated with the topical IL-1 receptor antagonist, reverse expression of IL-1β appeared, and a positive effect in promoting allograft survival and inhibited neovascularization of the cornea were observed. 39 –41 Acting as a pro-inflammatory cytokine, IL-1β affects the activation of vascular adhesion molecules, including intracellular adhesion molecule-1. 42 Upregulation of several CXC chemokines, such as IL-8 and GROα, which act as a chemoattractants for polymorphonuclear leukocytes, is also controlled by IL-1β. 43 In addition, IL-1β mediates murine CXC chemokines such as macrophage-inflammatory protein (MIP)-2 and KC, and it affects corneal angiogenesis by inducing vascular endothelial growth factor-A and cyclooxygenase-2/prostanoids. 44,45 Earlier studies have proven that intradermal injection of IL-1β in B/c mice produces multiple-fold expression of IL-1α, IL-1β, MIP-2, IL-10, TNF-α, and MHC class II in LCs, indicating that the component deriving from APCs might be important for triggering primary immune responses in the skin. 46
In summary, corneal transplantation is inevitably associated with trauma caused by the handling of the transplant, both pre- and intraoperatively. Epithelial damage leads to rapidly occurring apoptosis of the underneath keratocytes, irrespective of the nature of the trauma. 47,48 Taking into account the baculovirus anti-apoptotic p35 functions to epithelium, both caspase-dependent and caspase-independent anti-apoptotic processes need to be considered, along with their anti-inflammatory effects. These effects—when acting in conjunction—lead to reduced allosensibilization after transplantation in comparison with other allogeneic transplantations. Although we know that decreased priming of CD4+ cells in cervical LNs is derived from the epithelium of the composite graft, triggered by p35 expression, further experiments are needed to demonstrate the exact mechanisms responsible for immune sensibilization on a molecular level in corneal epithelial cells. The p35 gene therapy with its multifunctional impact has the potential to protect grafted transplants from host-versus-graft reactions, especially in high-risk engraftments. The inhibition of graft failure by modulated immune responses would be one option to decrease the need for re-transplantations.
Footnotes
Acknowledgments
The authors are grateful to Dr. Johannes Schwartzkopff for his advice on the animal experiments, and Dr. Pait Teesalu for his expertise in applying for funding. This study was supported by the Archimedes Foundation (Kristjan Jaak Scholarship, M.P., and DoRa 6 Scholarship, M.P.), the University of Tartu Foundation (Andreas and Elmerice Traks Scholarship, M.P., and Liisa Kolumbus Scholarship, M.P.), and the Bausch & Lomb Sicca-Förderpreis (M.P.).
Author Disclosure
No competing financial interests exist.