
Abstract
The hematopoietic zinc finger protein, Hzf, is induced in response to DNA damage or by Arf tumor suppressor in a p53-dependent manner. Recent studies have revealed that Hzf is an RNA-binding protein that regulates localization and translation of specific mRNA. The RNA-binding activity of Hzf is required for the functions of cerebellar purkinje cells and adipocytes, although their molecular mechanisms underlying the mRNA regulation largely remain unknown. To further investigate the molecular function of Hzf, we raised two rat monoclonal antibodies (MAb) against a peptide corresponding to the C-terminal region of the mouse/human Hzf protein. Both MAbs reacted with the native protein expressed in mammalian cells, and were highly efficient in detecting endogenous Hzf by immunoblotting, immunoprecipitation, and immunofluorescence. These MAbs should therefore be useful for further analysis of molecular functions of the Hzf protein and for identification of Hzf-binding proteins.
Introduction
Hzf protein contains three typical C2H2-type zinc-finger domains that can potentially serve as nucleic acid – binding motifs. Iijima et al. reported that Hzf associates through its zinc-finger motifs with the type 1 inositol 1,4,5-triphosphate receptor (IP3R) mRNA in cerebellar purkinje cells where it regulates the intracellular localization and translation of the mRNA.(5) We have shown that expression of Hzf is induced during adipocyte differentiation; cells with impaired Hzf expression are defective in undergoing efficient adipogenesis.(6) Hzf physically interacts with the C/EBPα mRNA to enhance its translation in adipocytes, and consistent with the role of C/EBPα in glucose metabolism, Hzf-null mice show a reduced insulin sensitivity that is reminiscent of human type 2 diabetes.
We now describe the production of two rat monoclonal antibodies (MAbs) directed to the mouse Hzf protein and demonstrate their utility for immunoblotting, immunoprecipitation, and immunofluorescence analyses. The availability of unrestricted quantities of high-affinity MAbs should facilitate further analysis of Hzf function and identification of Hzf-binding molecules.
Materials and Methods
Cells and culture conditions
NIH-3T3 fibroblasts, an Arf-inducible NIH-3T3 derivative cell line MT-Arf,(7) 3T3-L1 and 293T cells were maintained in Dulbecco's modified Eagle's medium (DMEM, Wako Pure Chemical Industries, Osaka, Japan) supplemented with 10% fetal calf serum (FCS) and penicillin/streptomycin. Peritoneal macrophages, P3U1 myeloma cells, and hybridoma cells were maintained in GIT medium (Nihon Pharmaceutical, Osaka, Japan) without antibiotics.
Immunization
A mouse/human Hzf C-terminal peptide (LHPAPGPIRTAHGPILFSPY) conjugated and unconjugated to keyhole limpet hemocyanin (KLH) was purchased from ThermoFisher Scientific (Sedanstrasse, Germany). This peptide sequence was utilized to establish rabbit Hzf polyclonal antisera in the previously study.(3) Fifty μg of KLH-conjugated Hzf peptide in Gerbu adjuvant (Gerbu Biotechnik, Gaiberg, Germany) were injected into the footpad of 6-week-old female Wister rats. Three boosters of 50 μg of Hzf peptide in Gerbu adjuvant at 10-day intervals were applied. Test bleeds obtained from the tail vein 1 week after each booster injection were screened for the presence of antibodies by enzyme-linked immunosorbent assay (ELISA) as described below. Once a satisfactory immune response was achieved, the animal was rested at least 10 days prior to a final injection. Four days after the final injection, the animal was sacrificed, lymph nodes were removed, and lymphoid cells were prepared for hybrid fusion.
ELISA screening
Polystyrene 96-well plates were coated with Hzf C-terminal peptide overnight at 4°C and blocked with 0.1% w/v bovine serum albumin (BSA) in PBS for 1 h at 37°C. The culture supernatants of hybridomas or serum from immunized animals were added and incubated for 1 h at 37°C. After washing three times with PBS, plates were incubated with alkaline phosphatase conjugated goat anti-rat IgG antibody (1:5000, Jackson ImmunoResearch, West Grove, PA) for 1 h at 37°C. After washing three times with PBS, plates were incubated with the substrate solution (Bio-Rad Laboratories, Hercules, CA). The absorbance was measured at 405 nm by microplate reader to obtain positive clones.
Cell fusion, hybridoma screening, subcloning, and isotyping
Single cell suspensions of rat lymphoid cells were fused with myeloma cells P3U1, at a ratio of 5:1, using GenomeONE-CF (Ishihara Sangyo, Osaka, Japan) according to the manufacturer's protocol. Hybridomas propagated on the X-ray irradiated peritoneal macrophages were selected in GIT medium supplemented with hypoxanthine-aminopterin-thymidine (HAT) (Sigma, St. Louis, MO). Supernatants were harvested from individual wells exhibiting hybridoma growth when cells were approximately 10% confluent and were tested by ELISA. Positive cultures were subcloned by limiting dilution (0.8 cell to 1 well in 96-well plate) in GIT medium supplemented with hypoxanthine-thymidine (HT, Sigma) and retested for immunoreactivity. The culture medium was gradually changed to GIT medium without HT and macrophage. Immunoglobulin (Ig) subclasses were determined using Rat Monoclonal Antibody Isotyping Test Kit (Serotec, Raleigh, NC).
Purification of MAbs
SCID mice pretreated with Pristane (Wako Pure Chemical Industries Ltd.) for 10 days were intraperitoneally injected with two positive hybridoma clones (231-9 and 391-18, 1 × 107 cells/clone). Two to 3 weeks later, ascitic fluid was obtained from these mice, and immunoglobulins were precipitated by slow addition of (NH4)2SO4 to a final concentration of 0.313 g/mL. Proteins were recovered by centrifugation, resuspended in 2.5 mL of PBS, and desalted by PD-10 desalting column (GE Healthcare UK, Buckinghamshire, United Kingdom). For some analyses, purified 231-9 was biotinylated using ECL Protein Biotinylation System (GE Healthcare UK).
Retroviral expression plasmid, retroviral vector production, and infection
293T cells were transfected with retroviral expression plasmids (MSCV p19ARF, Flag-Hzf, pSUPER retro sh-Hzf, and control plasmids(3)) together with helper retrovirus plasmids as previously described.(8) Culture supernatants were harvested 24 − 60 h after transfection, pooled, and stored on ice. Exponentially growing cells in 10-cm-diameter culture dishes were infected with 3 mL of fresh virus-containing supernatant in complete medium containing 8 μg/mL polybrene (Sigma). Infection was confirmed either by flow cytometric assay for green fluorescent protein expression or by selection for drug resistance.
Immunoblotting
Cells were lysed by addition of RIPA buffer (10 mM Na-phosphate [pH 7.2], 150 mM NaCl, 2 mM EDTA, 0.1% SDS, 1% Na-deoxycholate, 1% NP-40, and protease inhibitors [Roche, Mannheim, Germany]) and sonicated on ice for 20 s. Insoluble materials were pelleted by centrifugation at 15,000 g for 10 min at 4°C, and protein in the supernatant was quantified by BCA assay (Pierce, Rockford, IL). Twenty μg of proteins were separated on polyacrylamide gels containing SDS and transferred to polyvinylidene difluoride (PVDF) membranes (Millipore, Bedford, MA). Membranes were blocked in TBS-T (10 mM Tris [pH 7.4], 150 mM NaCl, and 0.1% Tween-20) containing 5% non-fat milk powder (Wako Pure Chemical Industries). Membranes were probed for 1 h with primary antibodies in TBS-T containing 5% non-fat milk powder, washed three times with TBS-T, and then incubated with appropriate secondary antibodies conjugated to horseradish peroxidase (HRP, GE Healthcare UK). After several washes with TBS-T, antibodies were visualized by incubation with Immobilon Western (Millipore). The Hzf protein was detected by two primary antibodies (231-9 and 391-18 or biotinylated 231-9) purified in our laboratory. Commercial primary antibodies were used to detect the Flag epitope tag (M2-HRP, Sigma), p19ARF (5C3-1, Santa Cruz Biotechnology, Santa Cruz, CA), C/EBPδ (C-22, Santa Cruz Biotechnology) and β-tubulin (Sigma).
Immunoprecipitation
Cell lysates were prepared using immunoprecipitation (IP) buffer (50 mM N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid [HEPES, pH 7.5], 150 mM NaCl, 1 mM EDTA, 2.5 mM EGTA, 1 mM dithiothreitol, 0.1% Tween-20) containing 10% glycerol and protease inhibitors, and sonicated on ice for 5 s. Insoluble materials were pelleted by centrifugation at 15,000 g for 10 min at 4°C, and protein in the supernatant was quantified by BCA assay (Pierce). Five μg of MAb-containing IP buffer (231-9 or 391-18) was incubated together with 20 μL of 50% slurry of protein G-Sepharose (GE Healthcare UK) and 500 μg of cell lysate in a total volume of 500 μL, and rotated at 4°C for 12 h. Immunoprecipitates were recovered by brief centrifugation and washed three times with 1 mL of IP buffer before being boiled for 5 min in 2 x Laemmli SDS gel sample buffer.
Immunofluorescence
Cells cultured on coverslips were washed three times in PBS, fixed for 15 min in 4% paraformaldehyde, washed in PBS, and permeabilized with 0.5% Triton X-100 in PBS for 15 min. All subsequent steps were performed at room temperature. Fixed coverslips were washed three times in PBS, blocked for 1 h with 10% FCS in PBS, rinsed three times PBS, and incubated for 1 h with 10 μg/mL of Hzf MAb (231-9 or 391-18) in PBS containing 10% FCS. Following three rinses in PBS, coverslips were incubated for 1 h with Cy3-labeled goat anti rat IgG (Jackson ImmunoResearch) in PBS containing 10% FCS to visualize primary antibodies. Coverslips were washed three times in PBS and mounted onto microscope slides using Vectashield containing DAPI (Vector Labs, Burlingame, CA). Stained cells were imaged with a fluorescent microscope (Zeiss, Göttingen, Germany).
Results
Establishment of rat MAbs against Hzf
Rats were immunized with a mouse Hzf C-terminal peptide (LHPAPGPIRTAHGPILFSPY) conjugated to KLH. The peptide has been previously utilized to obtain rat polyclonal antisera.(3) By ELISA assay using the peptide without KLH, we screened sera from immunized animals. Responding animal was sacrificed, and their lymphoid cells were subjected to hybrid fusions.
After dilution of fused cells into microtiter plates containing macrophages and selection in HAT medium, proliferating hybridomas were observed in 1004 of 1248 wells. In the first round of hybridoma screening by ELISA assay, 10 hybridoma supernatants reacted strongly with Hzf peptides. Cells growing in these wells were subjected to the limiting dilution for the second ELISA screening. Nine clones were obtained in the second screening, and all of these clones produced antibodies of the IgG2a isotype that reacted with the Flag-tagged Hzf protein expressed in NIH-3T3 cells by immunoblotting (data not shown). In the following analyses, two clones (231-9 and 391-18) were utilized.
Immunoblotting
We next checked whether the MAbs (231-8 and 391-18) can detect endogenous Hzf proteins by immunoblotting. Lysates prepared from NIH-3T3 cells infected with control or p19ARF expression retroviruses together with sh-Hzf viruses were separated on denaturing-polyacrylamide gel, transferred to membrane, and blotted using the Hzf MAbs. As a positive control, bacterially produced His-Hzf protein was used.(6) As shown in Figure 1A, both 231-9 and 391-18 MAbs were able to detect the endogenous Hzf proteins even in the absence of p19ARF, and retroviral expression of p19ARF resulted in further accumulation of the Hzf proteins as expected. In sh-Hzf-infected cells in which endogenous Hzf is efficiently suppressed,(3,6) Hzf proteins were hardly detectable by both MAbs, confirming the specificity of these MAbs. Additionally, both MAbs were able to detect the endogenous Hzf proteins in 3T3-L1 cells. The induction of Hzf proteins by adipogenic stimulation was clearly detected by both MAbs in control virus-infected 3T3-L1 cells but not in Hzf-knockdown cells (Fig. 1B), further demonstrating that these MAbs are highly efficient for detecting endogenous Hzf proteins by immunoblotting.
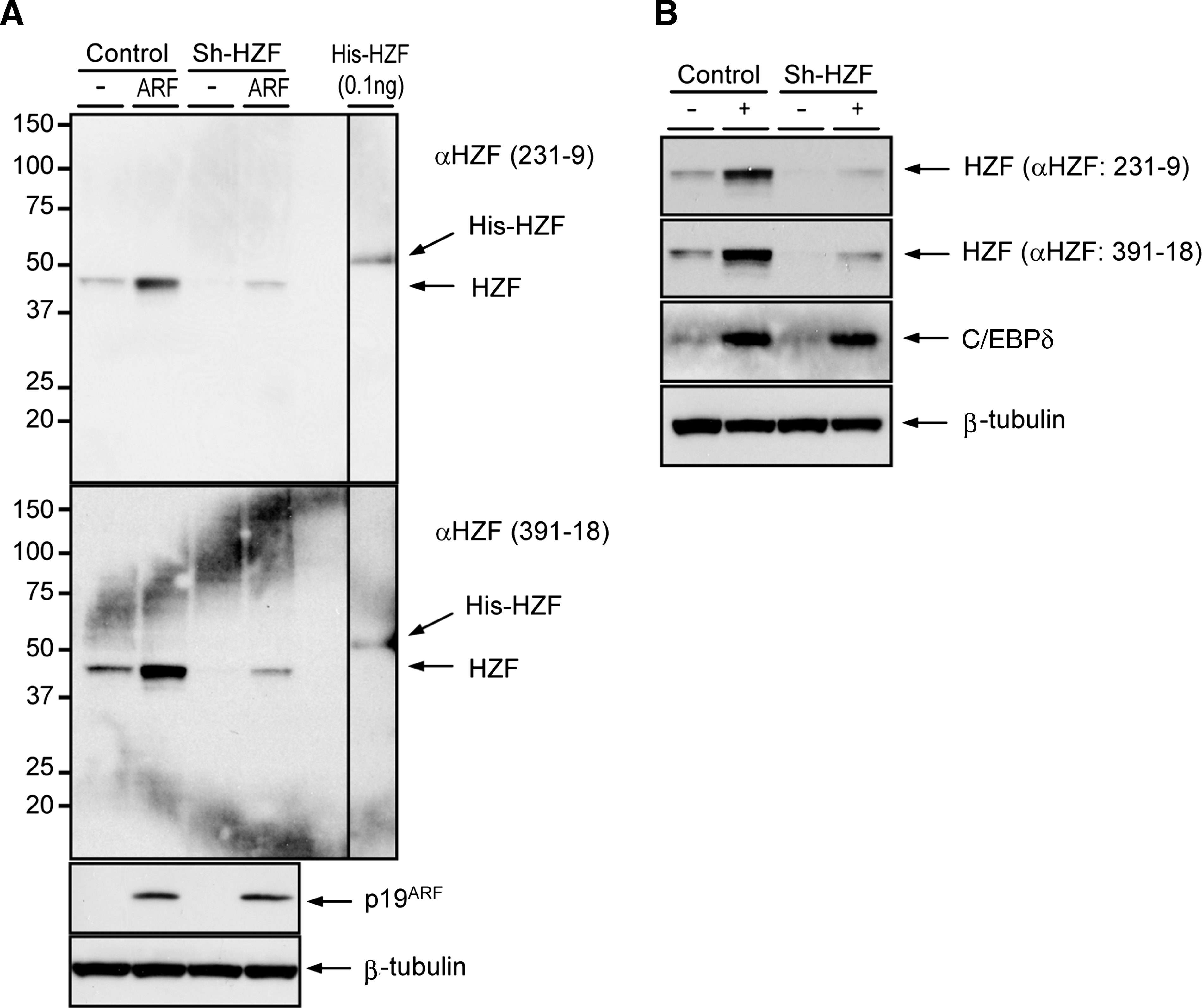
Immunoblotting of Hzf. (
Immunoprecipitation
To check whether these MAbs can immunoprecipitate Hzf protein, NIH-3T3 cells were infected with retroviruses encoding Flag-tagged mouse full-length Hzf. Lysates were mixed with 5 μg of 231-9 or 391-18 MAbs or without antibody as a negative control, and protein G-sepharose was added. Immunoprecipitated proteins were subsequently subjected to the immunoblotting using Flag antibody or biotinylated 231-9 Hzf MAb. Flag-tagged Hzf protein was precipitated in the presence of 231-9 or 391-18 MAb, but not in the absence of antibody (Fig. 2). Rabbit polyclonal Hzf antibody, we previously established,(3) was unable to precipitate Hzf protein (data not shown), indicating that murine Hzf protein can be precipitated efficiently by these MAbs compared with rabbit polyclonal antibody. Although both 231-9 and 391-18 MAbs are IgG2a subtype, only 231-9 MAb had reactivity with approximately 40 kDa protein when Flag-tagged Hzf protein was detected by M2-HRP (Fig. 2A), suggesting the possibility that 231-9 and 391-18 MAbs are not derived from identical clone.
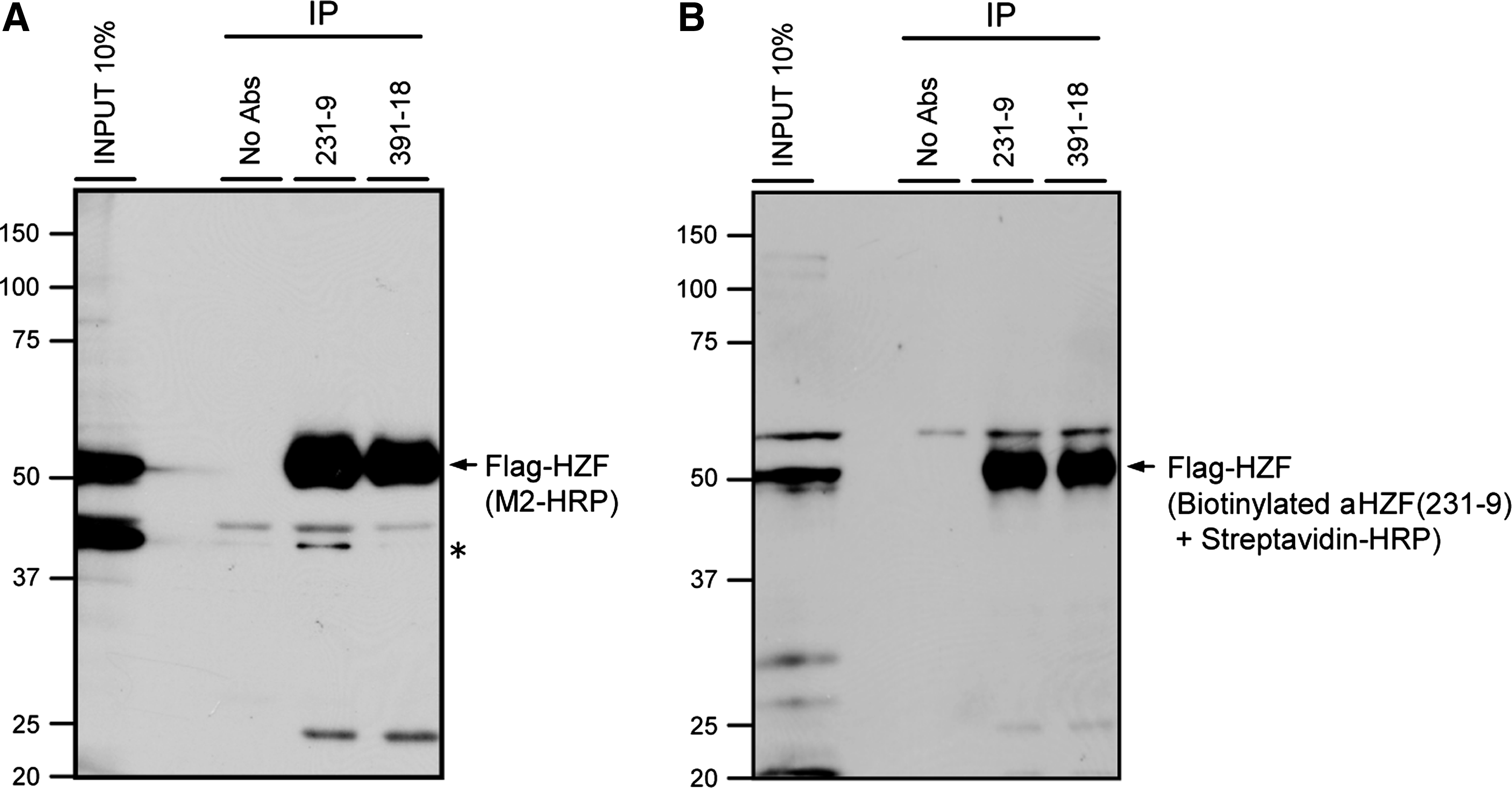
Immunoprecipitation of Hzf. Lysates of NIH-3T3 cells infected with Flag-tagged Hzf retroviruses were immunoprecipitated with MAb 231-9 or MAb 391-18. Immune complexes were recovered using protein G-sepharose, resolved on 10% polyacrylamide gels containing SDS, and probed with HRP-conjugated antibody to the Flag epitope (
Immunofluorescence
To determine whether the MAbs can microscopically localize Hzf within fixed cells, NIH-3T3, and its derivative Arf-inducible cell line, MT-Arf cells were grown on coverslips, fixed with paraformaldehyde, incubated with MAb 231-9 or 391-18, and then reacted with goat anti-rat IgG conjugated to Cy3. MT-Arf cells express exogenous p19ARF under the control of methallothioneine promoter, and the p19ARF protein can be induced by the addition of zinc sulfate into culture media, which consequently leads to the induction of endogenous Hzf proteins. Both MAbs specifically stained nucleoplasm in zinc-induced MT-Arf cells (Fig. 3). In addition, Hzf protein was weakly detected in uninduced MT-Arf cells with both MAbs or parental NIH-3T3 cells treated or untreated with zinc (data not shown), although no signal was detected without primary MAbs. These results were consistent with our previous results using rabbit polyclonal Hzf antibody that indicate Hzf protein is accumulated in the nuclei of MT-Arf cells treated with zinc sulfate.(3)
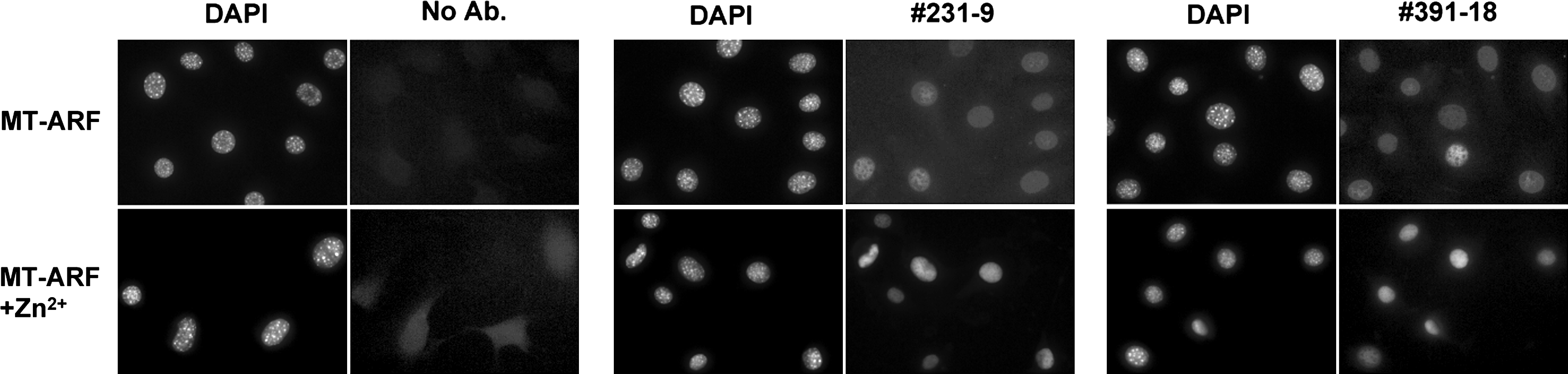
MAbs detected Hzf by immunofluorescence. MT-Arf cells grown on coverslips were treated or untreated with 80 μM zinc sulfate for 24 h, fixed with 4% paraformaldehyde, and subjected to immunofluorescence analysis using MAb 231-9 or 391-18. Cells were counterstained with DAPI.
Discussion
In this study, we generated rat MAbs 231-9 and 391-18 against Hzf C-terminal peptide. Compared with a rabbit polyclonal antibody previously established,(3) these MAbs were highly efficient in detecting endogenous and recombinant Hzf protein by immunoblotting and immunofluorescence. In addition, Hzf proteins were efficiently precipitated with these MAbs. The Hzf peptide used for immunization contains conserved region between mouse and human sequence, therefore these MAbs should also detect human Hzf protein.
Hzf interacts with IP3R and C/EBPα mRNAs through its C2H2 type zinc finger domains to regulate their localization and translation.(5,6) The precise mechanism underlying the mRNA regulation is currently unknown, and an identification of Hzf-interacting proteins will lead to the elucidation of molecular mechanism for mRNA regulation. To this end, the newly developed MAb in this study may be useful as these MAbs can precipitate Hzf protein from cell lysates with much more efficiency than previously developed rabbit polyclonal antibody. Furthermore, it will be possible to identify other RNA molecules targeted by Hzf through the purification of the Hzf complex using these MAbs in combination with DNA microarray.
We and others have reported that Hzf is a direct transcriptional target of p53 tumor suppressor.(3,4) Thus, expression of Hzf protein may be disturbed in cancer cells in which p53 pathway is inactivated. In addition, our recent observation indicated that Hzf plays an essential role in adipocyte differentiation and insulin-mediated glucose metabolism.(6) Human Hzf gene resides on chromosome 12q13, a locus involved in susceptibility to type 2 diabetes,(9,10) suggesting a possibility that Hzf level is disturbed on insulin-resistant patients. The MAbs might be, therefore, a useful tool, not only to study the molecular mechanism of Hzf-mediated RNA regulation, but also for the diagnosis of cancer and metabolic diseases such as type 2 diabetes.
Footnotes
Acknowledgments
We thank Drs. Martine F. Roussel and Charles J. Sherr of St. Jude Children's Research Hospital for providing MT-Arf cells and p19ARF and Hzf expression plasmids. We also wish to thank Hiroyuki Kawagishi for technical help.