
Abstract
A novel member of the human β3-galactosyltransferase family, the β3GalT7 gene (AY277592, EC2.4.1.-) was first isolated and cloned by our laboratory. To further study its functions, we constructed a prokaryotic expression system of β3GalT7 and obtained anti-β3GalT7 polyclonal antiserum by immunizing rabbit with purified β3GalT7 protein. Using the antiserum, the expression of β3GalT7 in various tissues and cell lines was analyzed by Western blot and immunochemical assays. Immunochemistry analysis showed the enzyme was expressed significantly higher in some tumor tissues than in normal tissues, indicating its biofunction in tumorogenesis. By immunofluorescence, the enzyme was observed highly accumulated in cytoplasm around nuclear membrane, implying that β3GalT7 may play an important role in the assembly of galactose in RER and Golgi.
Introduction
In the past few years, rapid advances in cDNA cloning achieved utilizing databases from genome and expressed sequence tag (EST) projects have led to the discovery of a large number of novel glycosyltransferase genes. With the aid of bioinformatics technology, a novel member of human β3-galactosyltransferase family—β3GalT7—was cloned from a human lung cDNA library(6) by our research group through homology screening in the NCBI database of EST with human β3GalT1 gene probe (GenBank accession no. AF117222). The new gene β3GalT7 containing an open reading frame (ORF) of 1191 bp was mapped to chromosome 19q13.2. The encoded protein includes a signal peptide and a galactosyltransferase domain with molecular weight and the isoelectric point of 43.3 kDa and 8.66, respectively. Northern blot indicated that β3GalT7 was highly expressed in lung, throat, and ileum and relatively lower in tongue, breast, womb, and testis.
Glycosyltransferases can be grouped into functional subfamilies based on similarities of sequence, which reflect their enzymatic character, donor specificity, acceptor specificity, and specific linkage between donor and acceptor. Compared with 6 β1,3-galactosyltransferases (β3Gal-T1,T2,T3,T4,T5,T)(7–11) and 5 β1,3-N-acetylglucosaminyltransferases (β3Gn-T1,T2,T3,T4,T5)(12–14) in the CaZy31 family, we found that the new gene was highly homologous, so it was named β3GalT7.
To further study its functions, we constructed a prokaryotic expression system of β3GalT7, obtained its polyclonal antiserum, analyzed its expression in several different cancer and normal tissues and cell lines, and detected its subcellular location through fluoroimmunocytochemistry. We demonstrated that β3GalT7 was abnormally expressed in malignant cells and tissues, indicating the relationship with malignancy and contributions to tumor biocharacteristics.
Materials and Methods
Cell lines and reagents
Bacterial strain Escherichia coli BL21 harboring recombinant vector β3GalT7-pGEX-6p-1 was produced by our laboratory.(6) Lung adenocarcinoma cell line SPCA-1, stomach adenocarcinoma cell line SGC-7901, mouse embryo fibroblasts cell line 3T3, and chronic marrow leukemia cell line K562 were purchased from American Type Culture Collection (Manassas, VA). The cells were cultured in RPMI 1640 (Gibco-BRL, Grand Island, NY) supplemented with 10% fetal calf serum (Hyclone, Logan, UT), 100 U/mL penicillin, and 100 mg/mL streptomycin.
Prokaryotic expression and purification of β3GalT7 protein
Comfirmed by DNA sequencing, the verified E. coli BL21/β3GalT7-pGEX-6p-1 clones were cultured at 37°C in 5 mL LB medium (100 mg/mL ampicillin) overnight. The next day β3GalT7 protein expression was induced by the addition of 1−2 mM isopropyl β-D-thiogalactoside (IPTG) to actively growing bacteria in LB medium at an A600 of 0.4-0.6. Incubation was continued for a further 8 h at 30°C. Then the bacteria were harvested by centrifugation. Overexpressed GST-β3GalT7 was purified by batch purification using Glutathion-Sepharose 4B beads equilibrated in the sonication buffer (50 mM Tris-HCl, 300 mM NaCl, 10% Glycero, 7 mM 2-ME [pH 7.8]). Elution was performed by incubating the bound resin with 5 mM reduced glutathione in 50 mM Tris-HCl (pH 8.6), 200 mM KCl, and 7 mM 2-ME for 10 min at room temperature. Purified by GST chromatography, the GST-β3GalT7 protein was quantified by SDS-PAGE electrophoresis.
Preparation and purification of polyclonal antibody
Twelve- to 14-month-old female New Zealand rabbits (Beerkape, Shanghai, China) were subcutaneously injected with purified GST-β3GalT7 protein in emulsified Freund's complete adjuvant (Sigma Chemical, St. Louis, MO) (1 mg/kg). Before immunization, 1∼2 mL of venous blood were sampled as control serum. Additional two to three boosters were applied at 3-week intervals with GST-β3GalT7 and incomplete Freund's adjuvant (Sigma Chemical) mixtures. Ten days after the last injection,(15) the antiserum was isolated and purified by 50% saturated ammonium sulfate and 33.3% saturated ammonium sulfate.
Western blot analysis
Total proteins of lung adenocarcinoma cell line SPCA-1, stomach adenocarcinoma cell line SGC-7901, large cell lung carcinoma H46, and chronic myelogenous leukemia cell line K562 were extracted and quantified by Bradford method.(16) Then, 30 μg protein from each cell lysate and purified GST-β3GalT7 fusion protein were boiled for 5 min with Laemmli buffer and loaded onto 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE 10%). By a semi-dry transfer system (Bio-Rad, Hercules, CA), the proteins were transferred onto PVDF membrane and stained with rabbit β3GalT7 polyclonal antibody followed by goat anti-rabbit IgG-AP. The protein was visualized by using the BM Chemiluminescence Western Blotting Kit (Boehringer, Mannheim, Germany) according to the manufacturer's protocol. The low-molecular-weight protein marker was the product of MBI (Burlington, Canada).
Tissue and cell immunochemistry
Immunochemistry analysis was performed to detect β3GalT7 protein expression in human stomach adenocarcinoma, cervical adenocarcinoma, and lung adenocarcinoma cell line SPCA-1. All the tissues were fixed with 10% neutral formalin, embedded in paraffin, and processed as 5-mm sections. Four or five adjacent ribbons were collected for histopathological analysis (hematoxylin and eosin stain) and immunohistochemical staining. Histopathological diagnosis for tumor tissues and nontumor tissues was performed according to cellular morphological changes and tissue architecture using established criteria. After dewaxing, endogenous peroxidase activity was blocked with 0.3% hydrogen peroxide for 5 min at room temperature, followed by incubation with normal non-immune goat sera for 30 min. Subsequently, sections were incubated with rabbit β3GalT7 antiserum (1:100 dilution) overnight at 4°C. Staining was continued with a biotin-conjugated antiprimary antibody, a streptavidin-peroxidase, and diaminobenzidine (Maxin-Bio Co., Fuzhou, China). Negative controls were run by replacing the primary antibody with PBS or normal mouse IgG1 (BD PharMingen, San Diego, CA). Results of β3GalT7 protein staining were categorized as either negative or positive based on the presence of clear brown color staining in the cytoplasm and cell membrane. Specimens with more than 10% positive cells were graded as positive.(16)
Immunofluorescence for subcellular localization of β3GalT7 protein
Stomach adenocarcinoma cells SGC-7901 and lung adenocarcinoma cells SPCA-1 were seeded on glass cover slips in 6-well dishes, then incubated overnight in RPMI 1640 with 10% fetal calf serum. Cells were fixed in 100% ice-cold acetone and saturated for 15 min in TBS-T (0.1%) containing 3% BSA. Rabbit anti-β3GalT7 polyclonal antiserum (1:200 dilution) was applied as the first antibody and incubated for 1 h. Rhodamine-labeled mouse anti-rabbit IgG (1:1000 dilution) was used as the second antibody. Cover slips were then rinsed with distilled water, and visualized by the Axioskop2 confocal microscope (Zeiss, Gottingen, Germany).
Results
Harvesting and identification of β3GalT7 antiserum
SDS-PAGE of whole bacteria lysates showed that GST-β3GalT7 fusion protein expression was highly induced by 1.5 mM IPTG for 8 h. The reducing SDS-PAGE analysis showed that molecular weight of the fusion protein was about 69 kDa, including 26 kDa of GST protein (Fig. 1). Therefore the molecular weight of β3GalT7 should be 43 kDa, which agrees with the predicted value from the gene sequence. New Zealand rabbits were then immunized with this protein to prepare polyclonal antiserum. After the purification, the titer of β3GalT7 antiserum turned out to be 1:64.
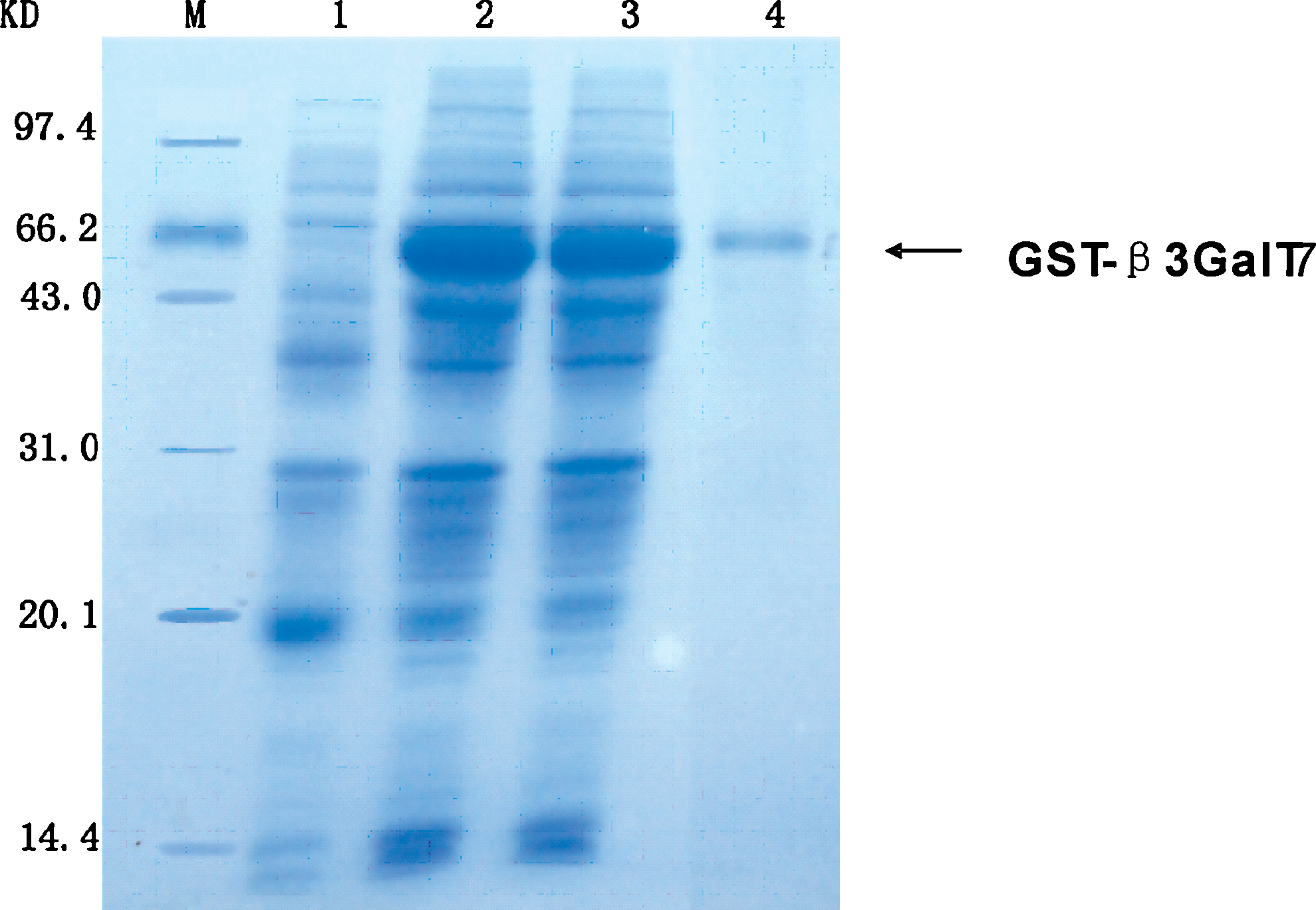
Expression of recombinant β3GalT7 in Escherichia coli strain BL21(DE3). Coomassie blue-stained SDS-PAGE gel of the β3GalT7 expression construct. M, molecular mass markers; lane 1, induced by IPTG; lanes 2 and 3, induced by adding IPTG to a final concentration of 1 mmol/L and 1.5 mmol/L, respectively; lane 4, purified GST-β3GalT7 protein.
Expression of β3GalT7 in tumor cell lines and binding specificity of antiserum
Different tumor cell lines were grown in tissue culture. Proteins were extracted from each cell line and then Western blot was performed. With the purified rabbit anti-β3GalT7 antiserum as first antibody and goat anti-rabbit IgG-Ap as the second one, Western blot showed that β3GalT7 was expressed in all four tumor cell lines SPCA-1, SGC-7901, H460, and K562. A clear band could be visualized illustrating highly specified reaction between purified β3GalT7 protein and the antiserum (Fig. 2).

Western blot analysis of β3GalT7 protein in four tumor cell lines. Lane 1, purified β3GalT7 protein (as positive control); lane 2, lung adenocarcinoma SPCA-1; lane 3, stomach adenocarcinoma SGC-7901; lane 4, large cell lung carcinoma H460; lane 5, chronic myelogenous leukemia K562.
Expression of β3GalT7 protein in human malignant tumor tissues
Tissue immunochemistry was carried out with the purified antiserum to detect β3GalT7 protein in human stomach adenocarcinoma, cervical adenocarcinoma, and lung adenocarcinoma cell line SPCA-1. As shown in Figures 3 and 4, in the stomach adenocarcinoma, the β3GalT7 expression was ++, normal stomach +. β3GalT7 expression had not been detected in normal cervix tissues, while in cervical cancer, it was ±. In 70% lung adenocarcinoma, β3GalT expression was +++. These results were consistent with RT-PCR results, demonstrating high expression of β3GalT in lung tumor. It was also indicated that β3GalT7 expression in the stomach and cervix tumor is obviously higher than that in normal tissues (Figs. 3 and 4).
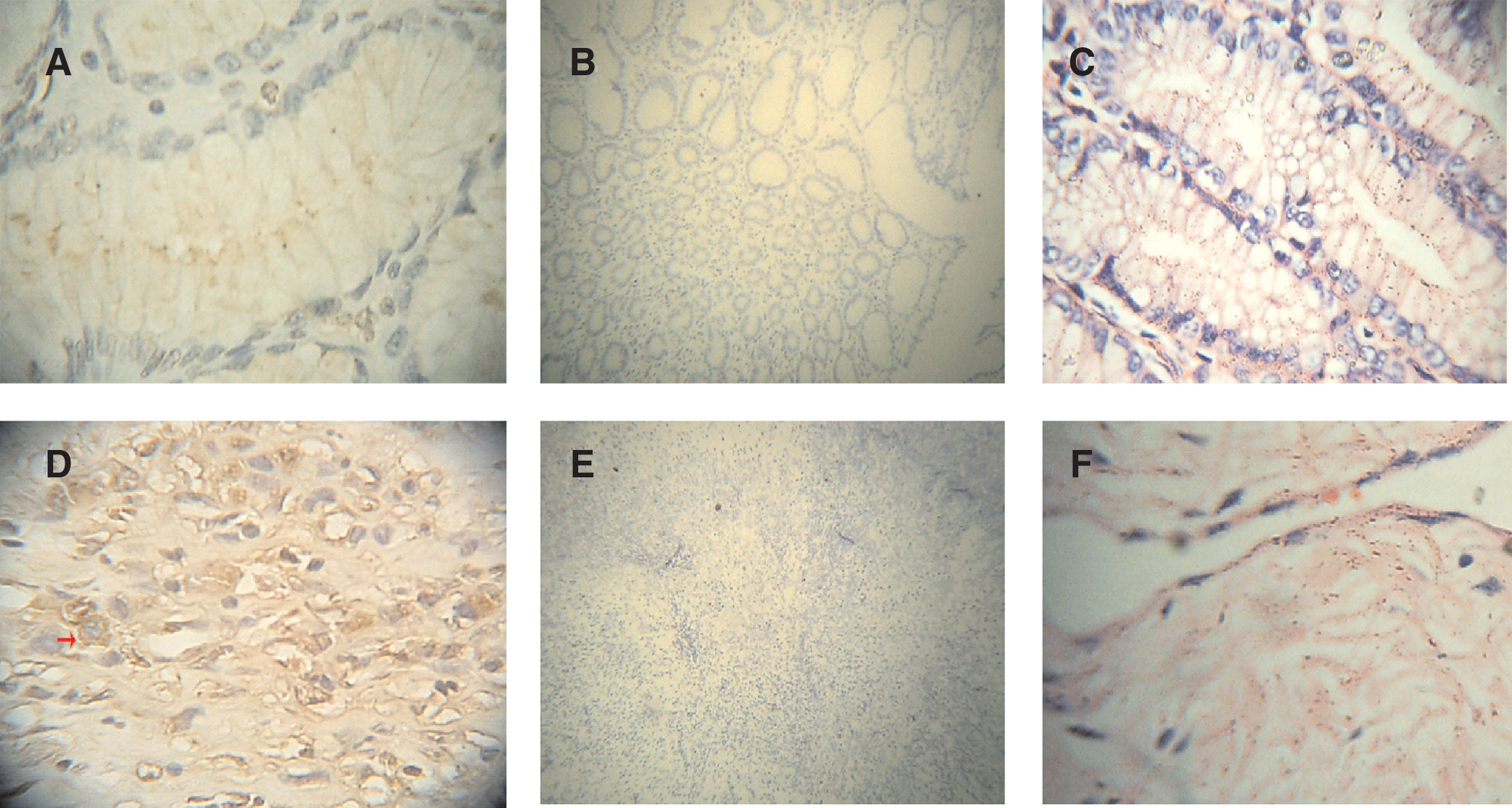
Immunochemistry analysis of β3GalT7 protein in human stomach adenocarcinoma tissue and normal tissue. (
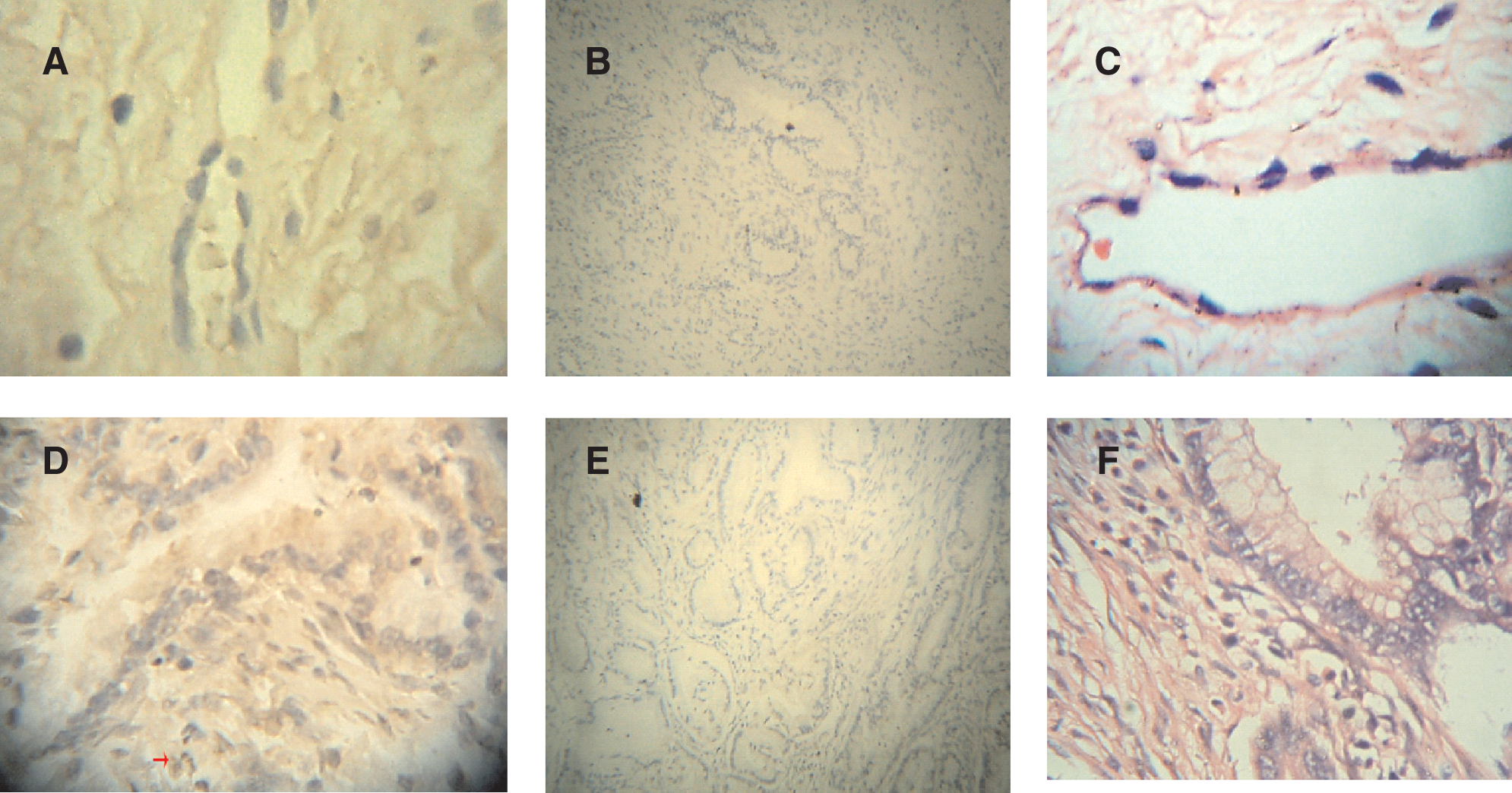
Immunochemistry analysis of β3GalT7 protein in human cervical adenocarcinoma tissue and normal tissue. (
Cell immunochemistry was employed to localize the protein. β3GalT7 protein was dyed brown, located around the nuclear membrane and the cytoplasm. The cell nucleus was dyed blue by hematoxylin (Fig. 5). Control group hybridized with rabbit serum sampled before immunization appeared negative.
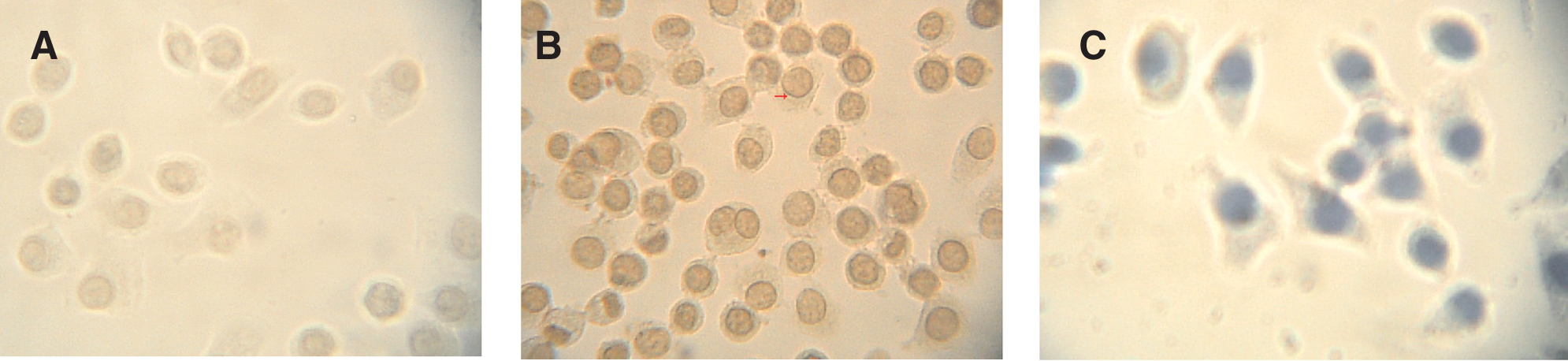
Immunochemistry analysis of β3GalT7 protein in human lung adenocarcinoma SPCA-1. (
Subcellular distribution of β3GalT7 in cancer cell lines
Cell immunofluorescence results indicated cytoplasmic accumulation of the emzyme β3GalT7 around the nuclear membrane (Fig. 6).
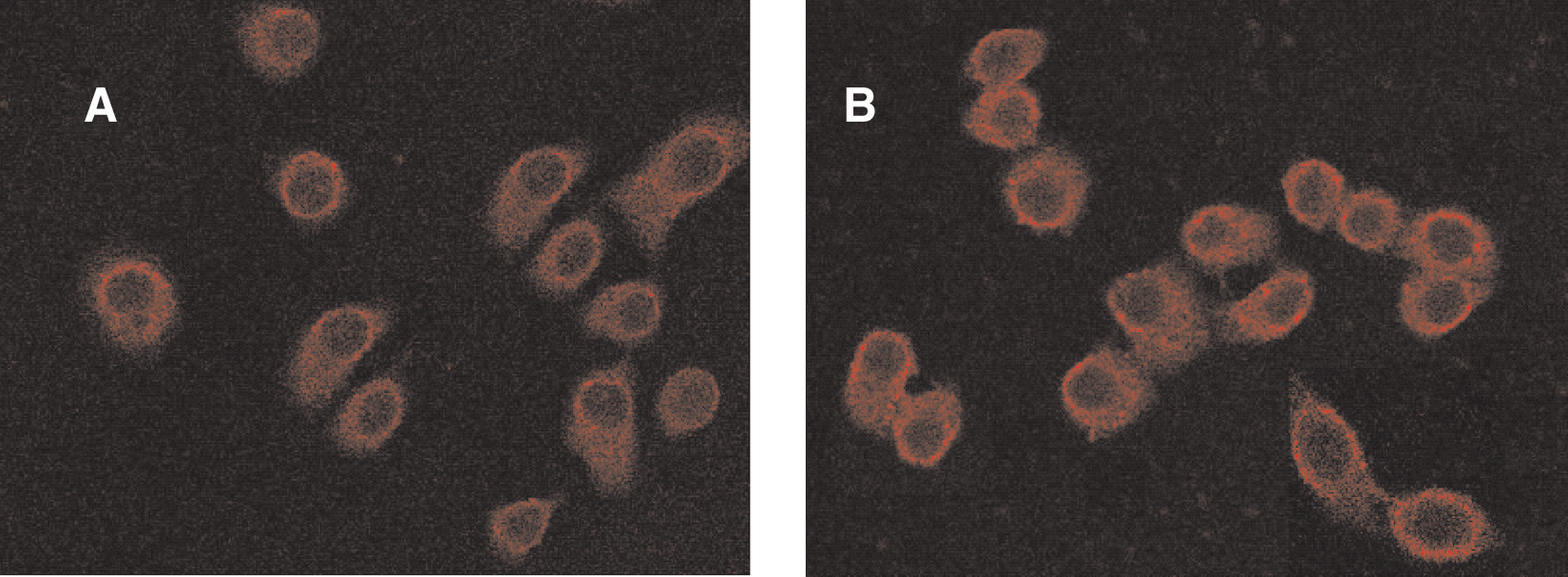
Fluoroimmunocytochemistry analysis of the subcelluar localization of β3GalT7 protein. (
Discussion
Glucose and galactose are essential metabolic substrates of all mammalian cells for energy demand. Increased need for glycolysis and galactose uptake for ATP production is observed in tumor cells, particularly in cells lacking oxygen supply. Because galactose is transported from blood to tumor, galactosed molecules must be delivered across galactose transporters of the vascular endothelium and tumor cells. Galactosyltransferase plays an important role in this process, and its bioactivities are closely correlated with tumor bioactivities.
The new gene of β3-galactosyltransferase β3GalT7 was first cloned and reported by our laboratory in 2004.(6) Having the β3-galactosyltransferase motifs, this gene was supposed to belong to β3-galactosyltransferase family and was designated as β1,3-galactosyltransferase-7 (β3GalT7) without evaluating its enzymatic characterization.(6) In 2005, Ishida and colleagues cloned the same gene from a human colon cancer cell line and named it β3Gn-T8, based on its position in a phylogenetic tree and enzymatic activity.(5) Thus, both β3GalT7 and β3Gn-T8 are authorized for this enzyme. In their study, Ishida et al. reported that the enzyme has a 1,3-GlcNAc transferase activity toward tetraantennary N-glycans and can extend polylactosamine on N-glycan. By quantitative real-time PCR, it was reported that the transcripts of β3Gn-T8 were abundant in most colorectal cancer cell lines and tissues. The enzyme may be involved in malignancy by synthesizing polylactosamine on β1-6 branched N-glycan in colon cancer.(5,17) To further detect the distribution of β3GalT7 and explore its biocharacteristics in tumorogenesis, we obtained purified protein and specific antibody of β3GalT7.
In genetic engineering, prokaryotic expression system has been a routine strategy for years to obtain target proteins(18–20) due to the following advantages: clear genetic background, easy to control, short expression cycle, high output, and low cost. The protein expressed by prokaryotic cells is usually in the form of non-fusion protein, fusion protein, secretion expression. The N or C terminal of fusion protein was fused with some non-target amino acid residue. In this way, we obtained the pure GST-β3GalT7 fusion protein. Its apparent molecular weight is 69 kDa, which was consistent with our prediction. This indicated that we have successfully constructed the recombinant vector of β3GalT7 and expressed the enzyme. After that, we obtained its high titer antiserum, which is indispensable for further research. Western blot (Fig. 2) demonstrated that: (1) the antiserum had high specificity to β3GalT7 and (2) SPCA-1, SGC-7901, and other cancer cell lines express β3GalT7.
For many years, antibody has been widely used in protein detection, localization, and quantitative analysis. Using β3GalT7 anti-serum, immunochemistry was employed and discovered that this enzyme was expressed significantly higher in some tumor tissues than in normal tissues, suggesting that β3GalT7 might, like its family member β3GalT5, play an important role in the tumorigenesis and invasion of tumor through participation in the synthesis of tumor-related antigen Slea. Furthermore, cell immunofluorescence analysis was used to locate the enzyme at the subcellular level. The results (Fig. 6) demonstrated that β3GalT7 protein was distributed in cytoplasma and was highly accumulated around the nuclear membrane, implying that, like other glycosyltransferases, β3GalT7 may also play an important role in the assembly of glycan in RER and Golgi.(21)
In conclusion, we have obtained the protein β3GalT7 and its antiserum, and that has laid a foundation for further functional research of this new gene.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (30670462) and the Undergraduate Scientific and Technological Innovation project of Jiangsu Provincial Education Department (CZ09B–030Z).