
Abstract
Katanin is a microtubule severing protein composed of two subunits, P60 and P80. The P60 subunit severs microtubules and P80 controls the microtubule severing activity of P60. Katanin promotes spindle shortening and severing processes in mitotic and meiotic spindles. In neurons, the level of P60-katanin was found to be very high in actively growing axons and at the tips of growing neuronal processes and in dendritogenesis. Inhibition of P60-katanin increases microtubule length throughout the neuronal cell body and mitotic/meiotic spindles, indicating that katanin is one of the key proteins for organizing microtubular structure in a variety of cell types. In this study, we produced 1G6, the first monoclonal antibody against P60-katanin. Recombinant P60-katanin (RecP60) protein, which was produced based on a specific region of rat P60-katanin, was expressed in Escherichia coli and used as antigen. 1G6 recognized the endogenous P60-katanin in Western blot analysis and immunocytochemistry in which monoclonal antibody would increase the range of applications. Since P60-katanin polyclonal antibodies have been used in studies until now, obtaining the hybridoma cell, the unlimited source of a monoclonal antibody against P60, results in an advantage in investigating P60-katanin functions.
Introduction
Studies suggest that katanin is responsible for changes in microtubule dynamics during mitosis. Katanin is concentrated at a microtubule-dependent structure at mitotic spindle poles, suggesting katanin may sever microtubules from their centrosomal attachments in non-neuronal cells.(5) Inhibition of katanin inhibits spindle-shortening processes in fibroblast mitotic spindles and Caenorhabditis elegans meiotic spindles.(6) Katanin may have other functions distinct from its role in cell division. In neurons, the level of P60-katanin was found to be very high in axons actively growing toward their targets, but then it decreased when the axon reached its target and stopped growing.(7) High P60-katanin levels were also found at the tips of growing neuronal processes at certain developmental stages and in dendritogenesis. Blocking the function of P60-katanin prevents microtubule release from the centrosome and profoundly increases microtubule length throughout the neuronal cell body, indicating that katanin-mediated microtubule severing is responsible for the release of microtubules from the neuronal centrosome.(8,9)
In this study, we produced the first monoclonal antibody against P60-katanin using rat sequence-based RecP60 protein as antigen. Since P60-katanin polyclonal antibodies have been used until now,(5,9) obtaining the hybridoma cell (the unlimited source) of the antibody against P60 results in an advantage for monitoring the behavior of endogenous P60 in cells and investigating the functions of P60-katanin.
Materials and Methods
Expression and purification of RecP60 (antigen preparation)
The amino acid sequence of the N-terminus part (amino acids 1–194) of the full-length rat P60-katanin (Gene Bank, the European Molecular Biology Laboratory, accession no. AY621629) was selected to make specific and soluble N-terminal His6-tag recombinant P60 protein. To achieve the specificity, the conserved C-terminal AAA domain was excluded from the full length of P60. To solve the solubility and folding problem, hydrophobic regions and cystein residues were excluded (Fig. 1A).
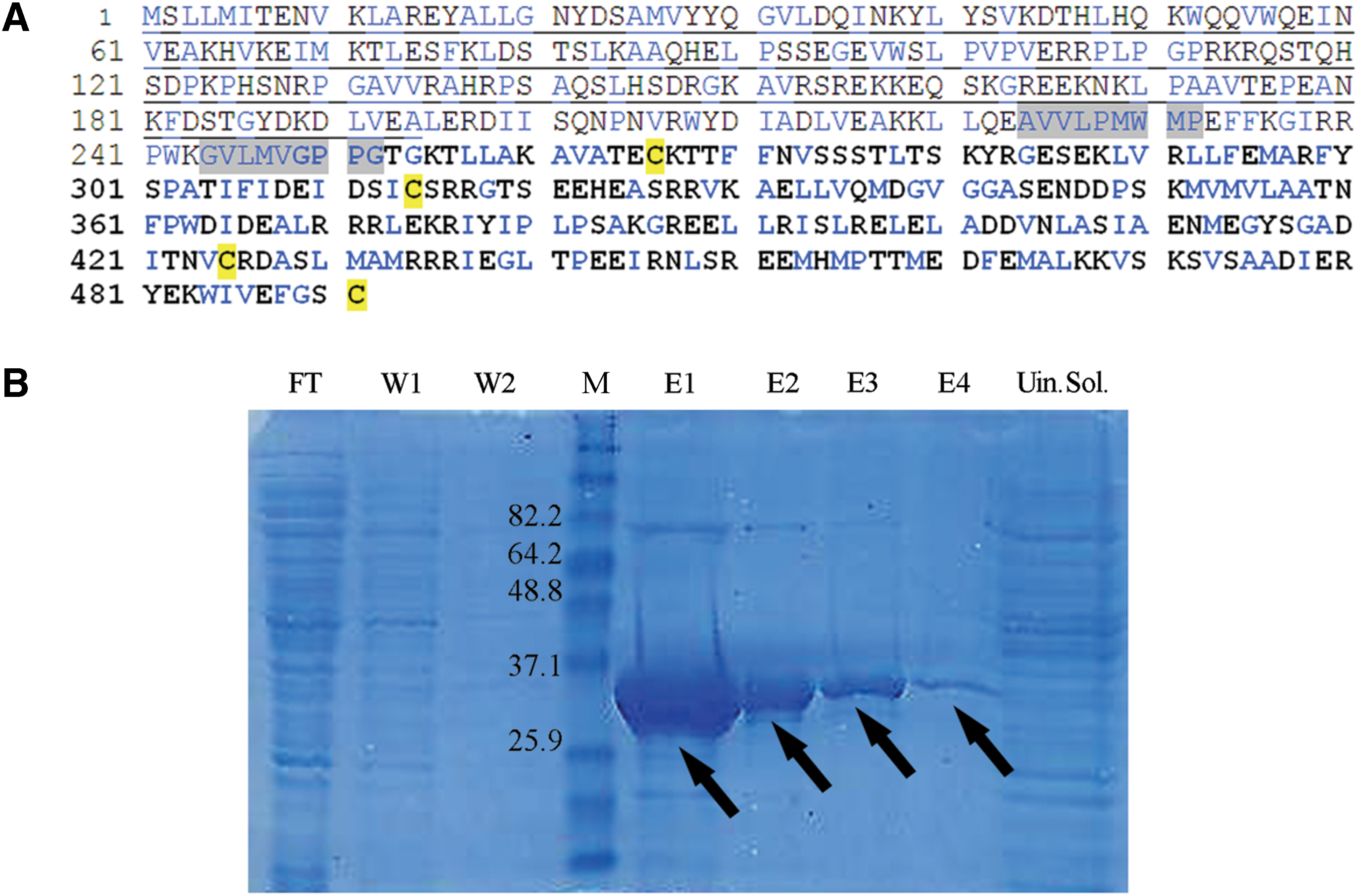
Property of RecP60 antigen.
RecP60, which was inserted in pET30a (Novagen, Darmstadt, Germany) vector, was expressed in BL-21(DE3)pLysS cells (Novagen) under isopropyl-β-D-1-thiogalactopyranoside (IPTG; AppliChem, Darmstadt, Germany) induction. His-tagged RecP60 was purified from the cell lysate by His-affinity resin (Ni-NTA agarose; Qiagen, Hilden, Germany) under native conditions to prepare the RecP60 antigen for immunization. After freezing overnight at −80°C, the cell pellet was resuspended in lysis buffer (50 mM NaH2PO4, 300 mM NaCl, 10 mM imidazole [pH 8.0]). Ni-NTA agarose was added to the supernatant of the lysate and incubated for 1 h. The beads were washed two times with washing buffer (50 mM NaH2PO4, 300 mM NaCl, 20 mM imidazole [pH to 8.0]), and RecP60 was eluted by elution buffer (50 mM NaH2PO4, 300 mM NaCl, 4 M imidazole [pH 8.0]). The subfractions of purification steps were analyzed with SDS-PAGE followed by Coomassie blue staining. Protein concentration was determined by Bradford assay.
Immunizations and analysis of the anti-RecP60 response
BALB/c mice were immunized intraperitoneally two times with 50 μg of RecP60/mouse at 2-week intervals. In the first immunization, RecP60 was injected with Freund's complete adjuvant (Sigma, St. Louis, MO) and in the second, RecP60 was injected with Freund's incomplete adjuvant (Sigma). After immunization, anti-RecP60 antibody response was assayed by indirect ELISA. ELISA plates, coated with RecP60 antigenic protein, were incubated with the mice sera. Anti-RecP60 antibody binding reaction was detected by using an alkaline phosphatase conjugated anti-mouse IgG (Sigma) or anti-mouse polyvalent (IgA, IgM, IgG) immunoglobulins (Sigma) as secondary antibody. Primary-secondary antibody binding reaction was visualized by p-nitrophenyl phosphate hydrolysis reaction. After 1 h, the optical density was read at 405 nm by using a microplate reader.
Mice that developed the IgG response against RecP60 were selected for fusion studies. One week before fusion, a booster immunization was done both intraperitoneally and to four foot pads of the selected mouse with 25 μg of RecP60 dissolved in PBS.
Hybridoma technology and monoclonal antibody production
Spleen and lymphoid cells of boosted BALB/c mouse were fused with FO myeloma cells (CRL-1646, ATCC)(10) in order to obtain hybridoma cells producing anti-RecP60 monoclonal antibodies. Fusion was performed using polyethylene glycol 4000 (PEG 4000, Merck, Darmstadt, Germany). Following fusion procedure, cells were plated in 96-well plates using DMEM with 20% fetal calf serum, 50 μg/mL gentamicin, and 1x hypoxanthine-aminopterin-thymidine (HAT, Gibco, Brooklyn, NY). Supernatants of growing hybridomas were screened with indirect ELISA against purified RecP60. Positive supernatants for RecP60 detected by using anti-mouse polyvalent immunoglobulins were selected, and their antibody producing cell lines were isolated individually by limited serial dilution. To understand whether the obtained monoclonal antibodies (MAbs) were IgG type or not, anti-mouse IgG was used as secondary antibody in the indirect ELISA. Selected hybridomas 1G6, 3H4, 3H6, 7F11, and 9H3 were cultured in large scale and their supernatants were used for further selection to analyze if they were specific for P60-katanin.
Immunocytochemistry
HeLa (human), SHSY-5Y neuroblastoma (human), and NIH/3T3 fibroblast (mouse) cells were grown on coverslips in 33 mm cell culture Petri plates for 48 h in DMEM containing 10% fetal calf serum and penicillin-streptomycin (10 U–10 μg/mL). The cells were fixed and permeabilized with methanol, and they were treated with IgG-type MAbs 1G6, 3H4, 3H6, and 9H3. To visualize these MAbs' recognition of intrinsic P60-katanin, cells were incubated with flourescence-labeled secondary anti-mouse IgG antibodies (Alexa 488, Alexa 546, or Cy5; Invitrogen, Brooklyn, NY).
To further confirm their localization, overlay of P60-katanin and tubulin was analyzed by using 1G6 MAb (monoclonal antibody) and rabbit anti-β tubulin antibody on methanol-fixed HeLa cells. Alexa-488 anti-mouse IgG and Alexa-647 anti-rabbit IgG antibody (Invitrogen) were used as secondary antibodies.
HeLa cells were transfected with the pEGFP-C1 vector (Clontech, Mountain View, CA) carrying full P60-katanin (human). For overexpression of the katanin constructs, Nucleofector (Amaxa Biosystems, Basel, Switzerland) electroporator was used. After nucleofection, cells were plated and fixed-permeabilized with methanol at 4, 8, 12, 24, 48, and 72 h to determine different expression levels of P60-GFP and then treated with 1G6 MAb and chicken anti-GFP antibody. Cy5-conjugated anti-mouse and FITC-conjugated anti-chicken IgY antibodies were used as secondary antibodies, respectively.
Images of the cells were obtained with a TCS SP2 AOBS inverted laser scanning confocal microscope (Leica, Wetzlar, Germany) using a 63x oil immersion objective.
Western blot analysis
For immunoblotting analysis, HeLa, NIH/3T3 fibroblast, SHSY-5Y neuroblastoma, brain, and liver tissues were washed in ice cold 1x PBS twice and homogenized in SDS-containing Laemmli sample buffer. Samples were separated on 10% SDS-PAGE and electrophoretically transferred to nitrocellulose membrane. After blocking the membrane for 30 min with 3% skimmed milk dissolved in PBS, 1G6 was added as a primary antibody and incubated overnight at 4°C. HRP-conjugated anti-mouse IgG was used as secondary antibody. Immunoreactive bands were visualized by addition of Luminol as the substrate and analyzed with CCD camera.
Results
Expression and purification of antigen RecP60
RecP60 corresponding to the N-terminus (amino acids 1–194) of full-length rat P60-katanin (Fig. 1A), fused with an N-terminal His6-tag, was expressed and purified from E. coli by Ni-NTA affinity and used for antigen production. The fractions of purification steps were analysed with SDS-PAGE. Overexpressed RecP60 was highly soluble, detected at the expected molecular weight of35 kDa, and the yield of purification was 4, 4 mg/L of bacterial culture (Fig. 1B).
Generation of monoclonal antibodies against RecP60
The purified RecP60 protein was used to immunize BALB/c mice, and hybridoma cells were generated by fusing the immune spleen and lymphoid cells with the myeloma cell line FO. The antibodies produced by the hybridomas were screened by indirect ELISA. Seven fusions were performed with RecP60-immunized mice. From the fourth fusion only one (3H4) and from the seventh fusion four (1G6, 3H6, 7F11, and 9H3) anti-RecP60 antibody producing hybridoma cells were obtained (Table 1). 1G6, 3H4, 3H6, 7F11, and 9H3 showed high anti-RecP60 immune response visualized after incubating with anti-mouse polyvalent (IgA, IgM, IgG) immunoglobulins. We also showed that 1G6, 3H4, 3H6, and 9H3 MAbs were IgG-type antibodies by using anti-mouse IgG antibody as secondary antibody in ELISA (Table 2). According to results shown in Table 2, 7F11 was not type IgG; it could be either IgA or IgM.
Spleen and lymphoid cells were mixed during seventh fusion.
1G6, 3H4, 3H6, and 9H3 are IgG-type antibodies.
Immunofluorescence staining of mitotic cells with 1G6
Obtained hybridoma cells producing IgG-type antibodies against recombinant P60 katanin, 1G6, 3H4, 3H6, and 9H3, were cultured in large scale, and their supernatants were collected to test whether they recognized endogenous P60 in cells. Only one monoclonal antibody, 1G6, reacted with P60-katanin specifically, which demonstrated the expected P60 interaction with microtubular structure, as shown by previous studies.(5,9) This single staining of intrinsic P60-katanin with 1G6 was tested on HeLa (human), SHSY-5Y neuroblastoma (human), and NIH/3T3 fibroblast (mouse) cells in order to analyze whether 1G6 recognized P60-katanin in different species. It was found that it recognized P60-katanin in both human and mouse. According to the results of single staining with 1G6, 1G6 appeared to stain P60-katanin of HeLa, neuroblastoma, and fibroblast cells, showing the specific localization on the mitotic spindle, consistent with the phases of the mitosis, highly concentrated at the remains of spindle microtubules at telophase and also on astral microtubules (Fig. 2).
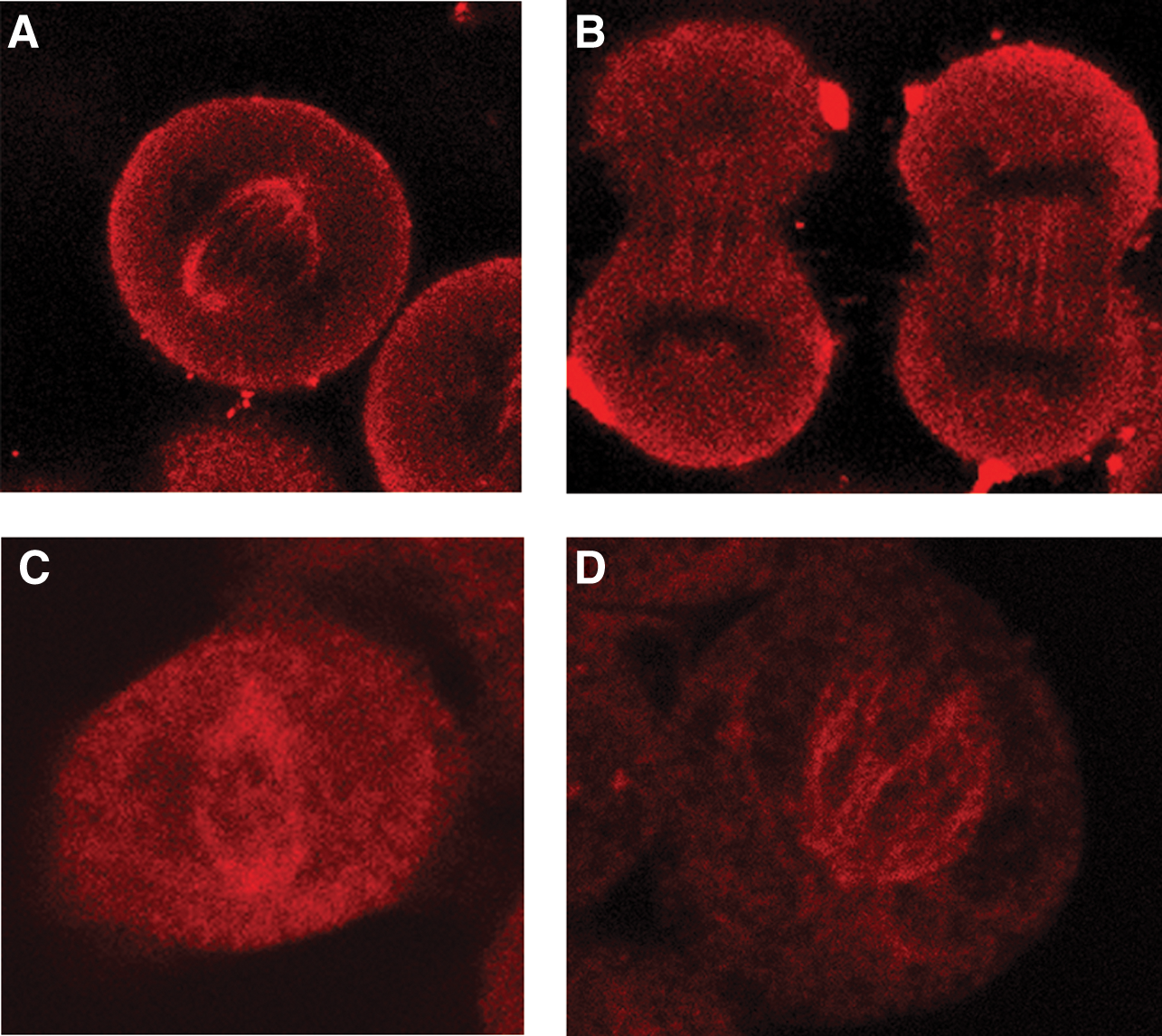
Immunofluorescence analysis of 1G6 recognition of P60-katanin on dividing HeLa, SHSY-5Y neuroblastoma, and NIH/3T3 fibroblast cells. 1G6 staining (red, Alexa 546) was localized on the mitotic spindle and also on astral microtubules
To show the P60-katanin localization on microtubule, 1G6 monoclonal antibody was used in double staining with tubulin. The 1G6 staining merged with β tubulin staining image was such that 1G6 was localized on the mitotic spindle and concentrated specifically on the microtubules towards the spindle poles, which is different from β tubulin staining (distributed on the entire microtubules of mitotic spindle in dividing cells) (Fig. 3).
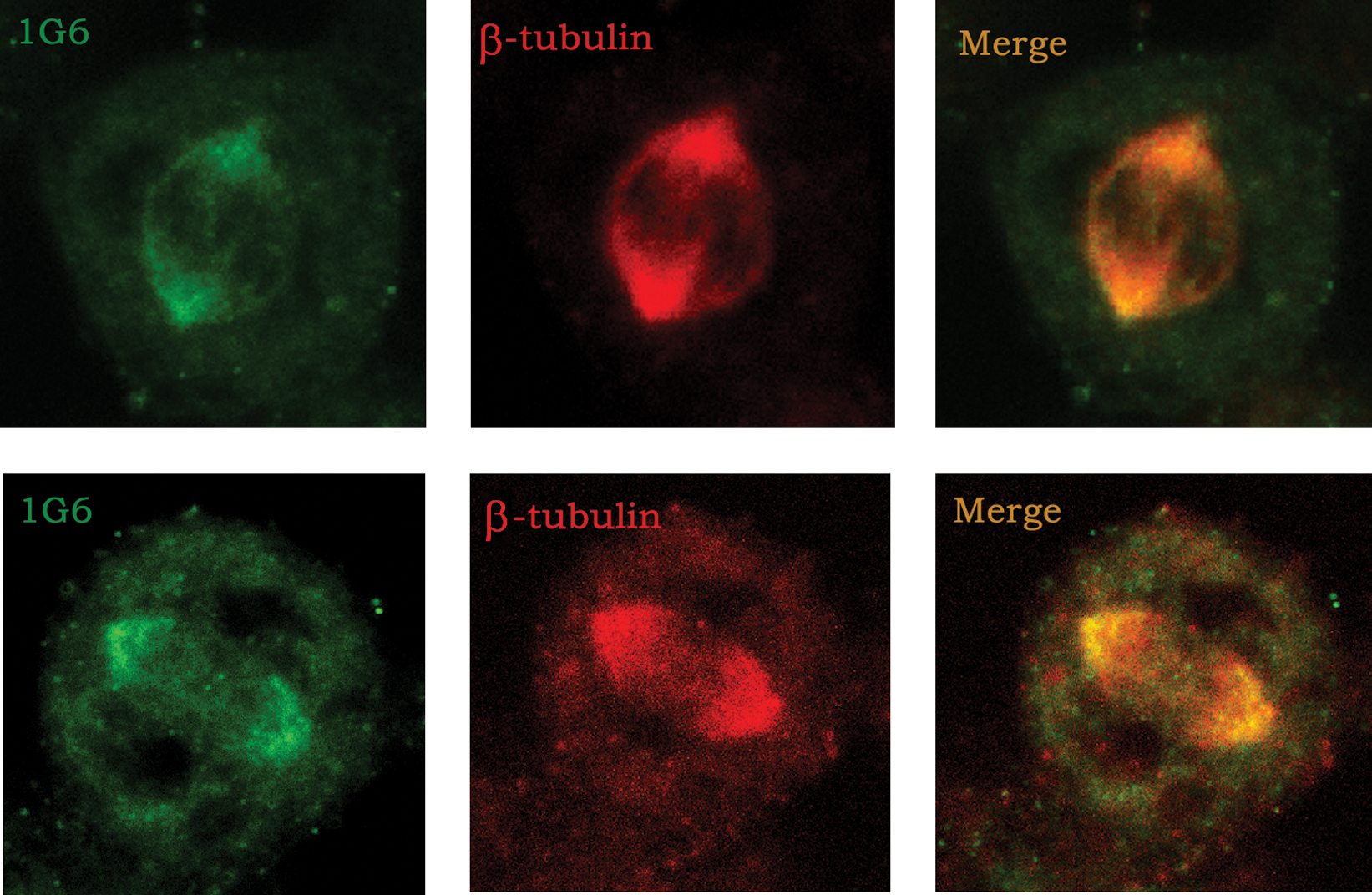
Comparison of tubulin and 1G6 localization on the mitotic spindle by immunocytochemistry. HeLa cells were stained with 1G6 (green, Alexa 488) and anti-β tubulin antibodies (red, Alexa-647). The panel at far right shows the overlay of both signals (yellow).
According to the results, 1G6 showed the expected P60-katanin interaction with microtubules and recognized P60-katanin of two species, human and mouse. These results are also consistent with the information that P60-katanin was available on microtubules to control the length of microtubules, which is necessary for the phases of cell division.
1G6 staining of rat/human P60-katanin coupled GFP protein in HeLa cells
To observe the recognition pattern of overexpressed P60-katanin with 1G6 on extrinsic katanin expression, HeLa cells were transfected with human-originated P60-katanin. The forced overexpressed P60-katanin seemed to specifically concentrate on the spindle poles (Fig. 4) preferentially at first, which is expected with P60-katanin's known function. At further time points, the overexpressed P60-katanin caused break-down of the microtubules, as shown in previous studies (data not shown).(9)
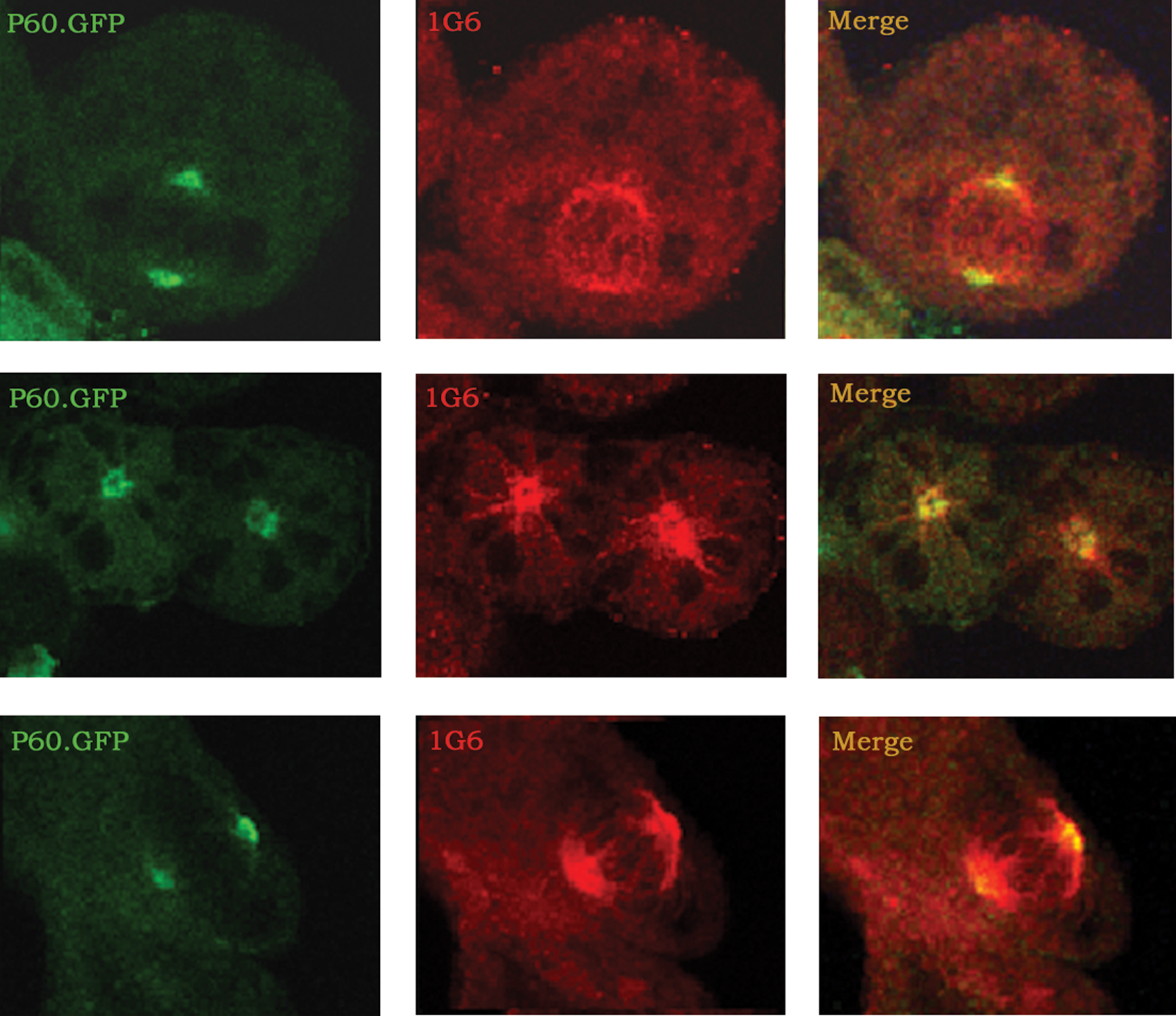
Human P60 expressed in dividing HeLa cells were visualized. P60-GFP expression (green) became dense at the poles of mitotic apparatus and merged with 1G6 staining (red).
Western blotting with anti-RecP60 monoclonal antibodies
Western blotting studies were done to test whether obtained MAbs would recognize native P60-katanin in tissues. Embryonic brain and liver tissues were used to test for 1G6, 3H4, 3H6, 7F11, and 9H3 MAbs. These five antibodies recognized RecP60 at 35 kDa. Yet, four of them (3H4, 3H6, 7F11, and 9H3) did not recognize a protein band at 55–65 kDa where native P60-katanin migrates (data not shown). Only 1G6 recognized both RecP60 at 35 kDa and a protein band at 55–65 kDa, which indicates the intrinsic katanin (Fig. 5).
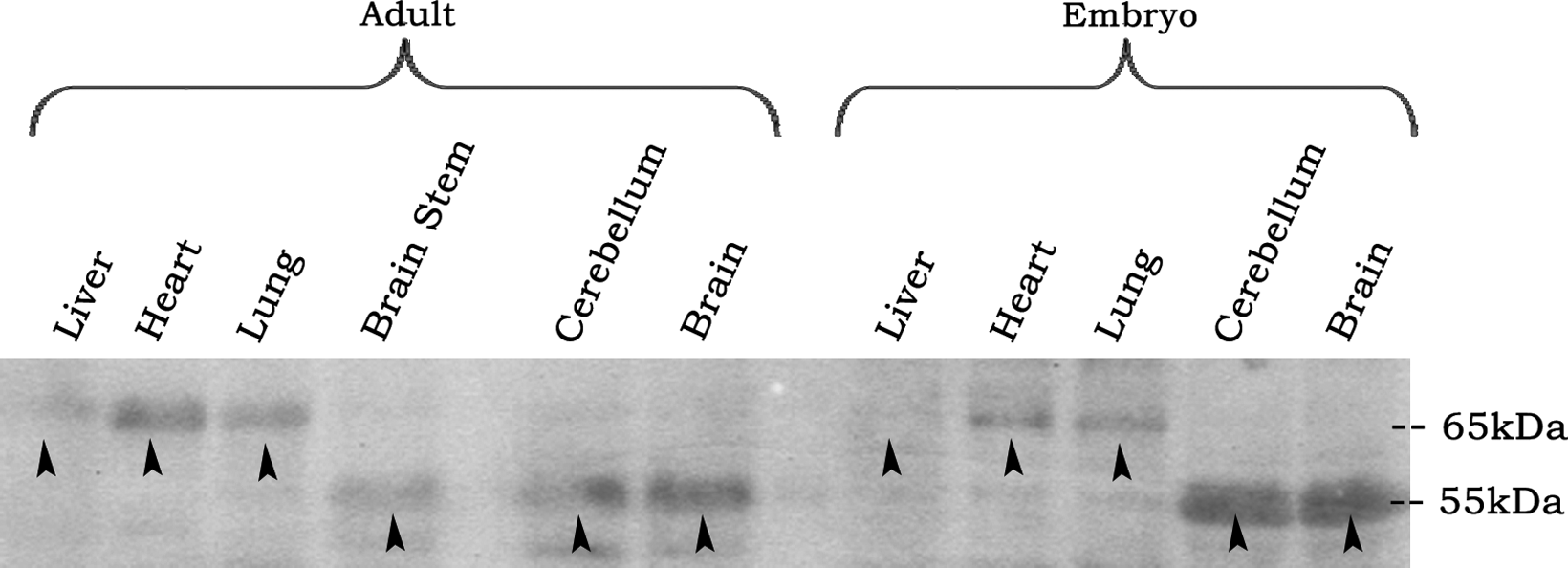
Western blot analysis of liver, heart, lung, brain stem, cerebellum, brain, and lysates of adult and embryonic rat tissues. 1G6 recognized a band at 55 kDa on brain, cerebellum, and brain stem, and at 65 kDa on liver, lung, and heart of adult and embryo (arrows).
1G6 MAb was examined with brain, brain stem, cerebellum, liver, lung, heart lysates of embryo, and adult rat tissues. 1G6 recognized a protein band at 55 kDa on brain, cerebellum, and brain stem, and at 65 kDa on liver, lung, and heart of adult and embryonic rats (Fig. 5). 1G6 also recognized 65 kDa band on HeLa, SHSY-5Y neuroblastoma, and NIH/3T3 fibroblast cells (data not shown), the same as in liver, lung, and heart tissues (Fig. 5).
Discussion
In this study, we used RecP60 as antigen and produced five MAbs against RecP60 by hybridoma technology. MAb 1G6 was the only antibody that recognized intrinsic P60-katanin on mitotic apparatus in dividing cells, which was shown by polyclonal antibodies in previous studies.(5,9) Hence, we could show that 1G6 would successfully work in immunocytochemistry applications. 1G6 is the first monoclonal antibody showing the specific cellular localization of P60-katanin and can be used to further understand the role of P60-katanin both structurally and functionally. This antibody would extend the range of applications and would especially enable multiple stainings.
The results of immunocytochemistry studies with 1G6 also provided evidence that P60-katanin was available on microtubules for its regulation to control the length of microtubules, which is necessary for the phases of cell division.(6)
The immunocytochemistry studies also indicate that 1G6 MAb is specific to P60-katanin of two species, mouse and human, because it recognized native P60-katanin of human-origin HeLa, SHSY-5Y neuroblastoma cells, and mouse-origin NIH/3T3 fibroblast cells. 1G6 was obtained by immunizing mouse with RecP60, which was expressed based on the sequence of rat P60-katanin. Amino acid sequences of human P60-katanin and mouse P60-katanin were blasted with rat P60-katanin and gave a high homology of 93% with human and 95% with mouse. This indicated that it is possible for 1G6 to recognize P60-katanin of three types of organisms, and this observation was confirmed with immunocytochemistry and Western blot studies.
Western blot analysis of P60-katanin with 1G6 on cell lines and tissues yielded protein bands at 65 kDa and 55 kDa, indicating that there may be alternative spliced forms of P60-katanin depending on tissue types. A 65 kDa band was obtained on liver, heart, and lung of adult and embryonic rat tissues and in HeLa, SHSY-5Y, and NIH/3T3 cell lines, which were all mitotic. A 55 kDa band was shown to be present in non-mitotic cerebellum and brain tissues of adult and embryonic rat tissues and also in the brain stem of adult rat tissue. From these results, it is possible to conclude that 1G6 MAb interacts with the common epitope of alternative-spliced forms of P60-katanin of both tissues.
As a result, we generated the first monoclonal antibody against P60-katanin, named 1G6, which recognizes P60-katanin of multiple species and is suitable for immunofluorescence stainings and Western blotting, making it a valuable tool to investigate P60-katanin function.
Author Disclosure Statement
The authors have no financial conflicts to declare.
Footnotes
Acknowledgments
We thank the other members of our lab for valuable discussions and help, especially our previous master's degree student Agne Stonkute for her work in preparing the RecP60 katanin antigen. This work was supported by research grants from TUBITAK (104T221TBAG), TUBA-GEBIP, and TWAS 04-375 RG/BIO/AS (AK).