
Abstract
Double homeobox 4 (DUX4) is a candidate disease gene for facioscapulohumeral dystrophy (FSHD), one of the most common muscular dystrophies characterized by progressive skeletal muscle degeneration. Despite great strides in understanding precise genetics of FSHD, the molecular pathophysiology of the disease remains unclear. One of the major limitations has been the availability of appropriate molecular tools to study DUX4 protein. In the present study, we report the development of five new monoclonal antibodies targeted against the N- and C-termini of human DUX4, and characterize their reactivity using Western blot and immunofluorescence staining. Additionally, we show that expression of the canonical full coding DUX4 induces cell death in human primary muscle cells, whereas the expression of a shorter splice form of DUX4 results in no such toxicity. Immunostaining with these new antibodies reveals a differential effect of two DUX4 isoforms on human muscle cells. These antibodies will provide an excellent tool for investigating the role of DUX4 in FSHD pathogenesis.
Introduction
Overexpression of DUX4 in cultured muscle cells has been shown to induce apoptosis(10) and increase oxidative stress,(11) possibly reflecting mechanisms of muscle deterioration seen in FSHD. Currently, to our knowledge, there is only one anti-DUX4 monoclonal antibody reported in the literature, called 9A12.(9) While it has been useful, the mouse monoclonal 9A12 antibody does not distinguish DUX4 from DUX4c, another double homeobox that shares more than two thirds of its sequence with DUX4.(12,13) It is important to distinguish the two species as DUX4c has also been proposed as a candidate gene for FSHD. Thus, we raised our antibodies against the unique C-terminus region of DUX4 in addition to the shared N-terminus. In this study, we generated three mouse monoclonal antibodies, P4H2, P2G4, and P2B1, and two rabbit monoclonal antibodies, E5-5 and E14-3, and we report the characterization of these novel monoclonal antibodies to human DUX4.
Materials and Methods
Expression and purification of DUX4 fusion protein
The sequence encoding the last C-terminal 76 amino acids of DUX4 was amplified by PCR using forward primer 5′-CGCGGATCCCCATGCAAGGCATCCCGGCGC-3′ and reverse primer 5′-CCGGAATTCCTAAAGCTCCTCCAGCAGAGCCCG-3′ and cloned in frame after glutathione-s-transferase in the bacterial expression vector pGEX-3x (Glutagene, Amrad, Kew, Australia). The sequence encoding the first N-terminal 159 amino acids of DUX4 was amplified by PCR using forward primer 5′-CGCGGATCCCCATGGCCCTCCCGACACCCTC-3′ and reverse primer 5′-CCGGAATTCCTGCGCGGGCGCCCTG-3′ and cloned in frame after glutathione-s-transferase in pGEX-3x. The template used for both N- and C-termini cloning was the previously described pCS2+mkgDUX4.(8) Expression of the fusion protein in E. coli strain BL21DE3pLysS was induced by 1 mM isopropyl-β-
Antibody production and screening
Purified and concentrated fusion protein was used as immunogen. Antibody production was done in collaboration with Epitomics (Burlingame, CA) for the rabbit monoclonal antibodies (MAbs) and by the Antibody Development Laboratory at the Fred Hutchinson Cancer Research Center (Seattle, WA) for the mouse MAbs. The mouse MAbs will be commercially available. The rabbit MAbs will be available through Epitomics. The antisera from all animals were screened for reactivity by ELISA against the immunogen, and Western blots and immunofluorescence for against transfected DUX4. The best rabbit and mouse were chosen for fusion, and the subsequent hybridoma clones were screened by ELISA for positive reactivity for the fusion protein and negative reactivity for GST protein alone. To narrow the selection of clones, they were further screened by Western blots and immunofluorescence on transfected DUX4. The best clones were subcloned, expanded, and isotyped by the production facilities.
Expression plasmids and ectopic expression
The pCS2+mkgDUX4(8) and pClneo-DUX4c(10) expression vectors used were previously described. The murine myoblast line C2C12 was maintained in Dulbecco's modified Eagle's medium supplemented with 10% (v/v) fetal bovine serum and 1% penicillin/streptomycin (Gibco, Carlsbad, CA) at 37°C in an atmosphere containing 5% CO2. Transient transfections were performed using SuperFect reagent (Qiagen, Valencia, CA) according to the manufacturer's specifications, and cells were harvested 24 h post-transfection for lysate or direct fixation. pclBABE+DUX4 was constructed by inserting the mkgDUX4 from pCS2 construct into blunted BamHI and EcoRI sites of a pclBABE vector (similar to Addgene Plasmid 20917, in place of MyoD) and transfected into C2C12 myoblasts using SuperFect as described above, and then collected for staining at 42 h post-transfection. Lentiviral constructs were made by inserting the coding sequences of full length and short DUX4 in place of GFP in the plasmid pRRLSIN.cPPT.PGK-GFP.WPRE (Addgene, Cambridge, MA). Full length and short DUX4 are referred to as DUX4-fl and DUX4-s, respectively, in the report by Snider and colleagues.(15)
Western blot analysis
Cells were directly lysed in 2x Laemmli sample buffer and sonicated to shear genomic DNA. Equal amounts of untransfected C2C12, transfected DUX4, and transfected DUX4c lysates were loaded onto 10% bis-tris polyacrylamide gels and separated by electrophoresis. Proteins were transferred onto nitrocellulose membranes (Invitrogen, Carlsbad, CA) and blocked with 5% non-fat dry milk in phosphate-buffered saline (PBS) and 0.1% Tween-20 (PBST). The membranes were then separately incubated in appropriate dilutions of E5-5, E14-3, and P4H2 hybridoma supernatants in PBST overnight at 4°C. Blots were washed in PBST and incubated with appropriate secondary antibodies, HRP-conjugated goat-anti-rabbit IgG, or HRP-conjugated goat-anti-mouse IgG (Jackson ImmunoResearch Laboratories, West Grove, PA), and subsequently washed again. Pierce ECL Western Blotting Substrate (Thermo Scientific Pierce, Rockford, IL) was added to membranes, and then blots were exposed to film and developed. Blots were stripped and reprobed with anti-α tubulin antibody (Sigma, St. Louis, MO) as a loading control.
Immunofluorescence
Cells were gently washed in PBS and fixed in 2% paraformaldehyde for 7 min at room temperature and then washed twice with PBS. Cells were permeabilized with 1% Triton X-100 (Sigma) in PBS for 10 min at room temperature with gentle rocking. Primary antibodies E5-5, E14-3, and P4H2 were added at appropriate dilutions in PBS overnight at 4°C and then washed with PBS. TRITC- or FITC-conjugated goat anti-rabbit or mouse secondary antibodies (Jackson ImmunoResearch Laboratories) were added for incubation for 1 h at room temperature and then washed with PBS and counterstained for DNA with 4′,6′-diamidino-2-phenylindole (DAPI). Cells were examined under a fluorescence microscope.
Results
Production of antibodies against human DUX4 N- and C-termini
The first 159 amino acid sequence of DUX4 was used as the N-terminus antigen and the last 76 amino acid sequence of DUX4 was used as the C-terminus antigen, which is not shared by DUX4c (Fig. 1A). To generate the recombinant C-terminus antigen for immunization, the last 228 base pairs of DUX4 open reading frame was cloned in frame after a GST tag and expressed in Escherichia coli. Induction of E. coli using 1 mM IPTG resulted in a high level of expression of a protein of the expected size at 36 kDa (Fig. 1B). The expressed product was purified on a glutathione-coupled agarose column and eluted by competition with free glutathione. Peak A280 fractions eluted from the column were pooled and showed a high level of purity by SDS-PAGE (Fig. 1B, lanes 3, 4). The purified GST-DUX4 C-terminus was concentrated and injected into two rabbits and two mice for production of antibodies. Production of the N-terminal antigen was performed in the same manner and yielded similar purity (data not shown), and was injected into two rabbits and two mice for development of antibodies.
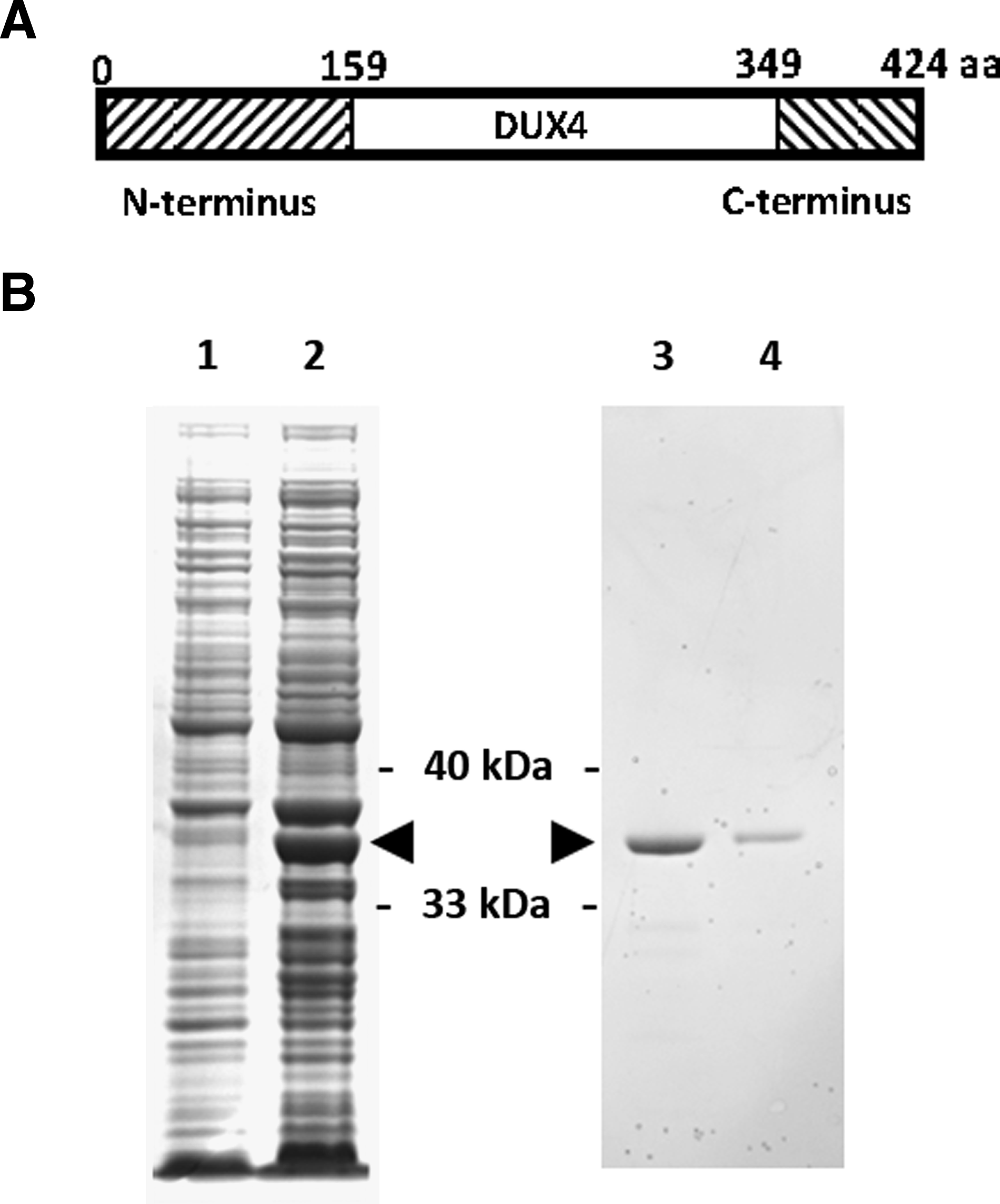
Production of recombinant DUX4 antigen. (
The antisera from all animals were screened for reactivity by Western blots and immunofluorescence against transfected DUX4 (data not shown). After fusion, the subsequent hybridoma clones were screened by ELISA for positive reactivity for the fusion protein and negative reactivity for GST protein alone. To narrow the selection of clones, they were further screened by Western blots and immunofluorescence on transfected DUX4. The clones with the best reactivity from screening were mouse MAbs P4H2, P2G4, and P2B1 and rabbit MAbs E5-5 and E14-3 (Table 1).
IF, immunofluorescence.
Less effective.
P4H2, P2G4, P2B1, E5-5, and E14-3 specifically recognize DUX4 protein
C2C12 mouse myoblast cells were transfected with plasmid expressing human DUX4. The sequence contained in the expression plasmid includes an upstream alternative initiation site about 60 amino acids away, thus producing two protein species differing by approximately 10 kDa in molecular weight. P4H2, E5-5, and E14-3 were all reactive to transfected DUX4 and none showed reactivity against untransfected C2C12 lysate by Western blot analysis (Fig. 2A). Mouse MAbs P2G4 and P2B1 were less effective on the Western blot method (data not shown). To assess for cross-reactivity against DUX4c, we transfected C2C12 cells with a plasmid expressing human DUX4c. Mouse monoclonal P4H2 and rabbit monoclonal E5-5 displayed no reactivity against DUX4c, whereas E14-3 also recognized DUX4c, which shares its N-terminus antigen sequence (Fig. 2A). Western blot using the previously characterized 9A12 monoclonal antibody confirmed the identity of DUX4 (Fig. 2B). Blots were stripped and reprobed with anti-α-tubulin antibody and showed similar loading of all samples per blot.
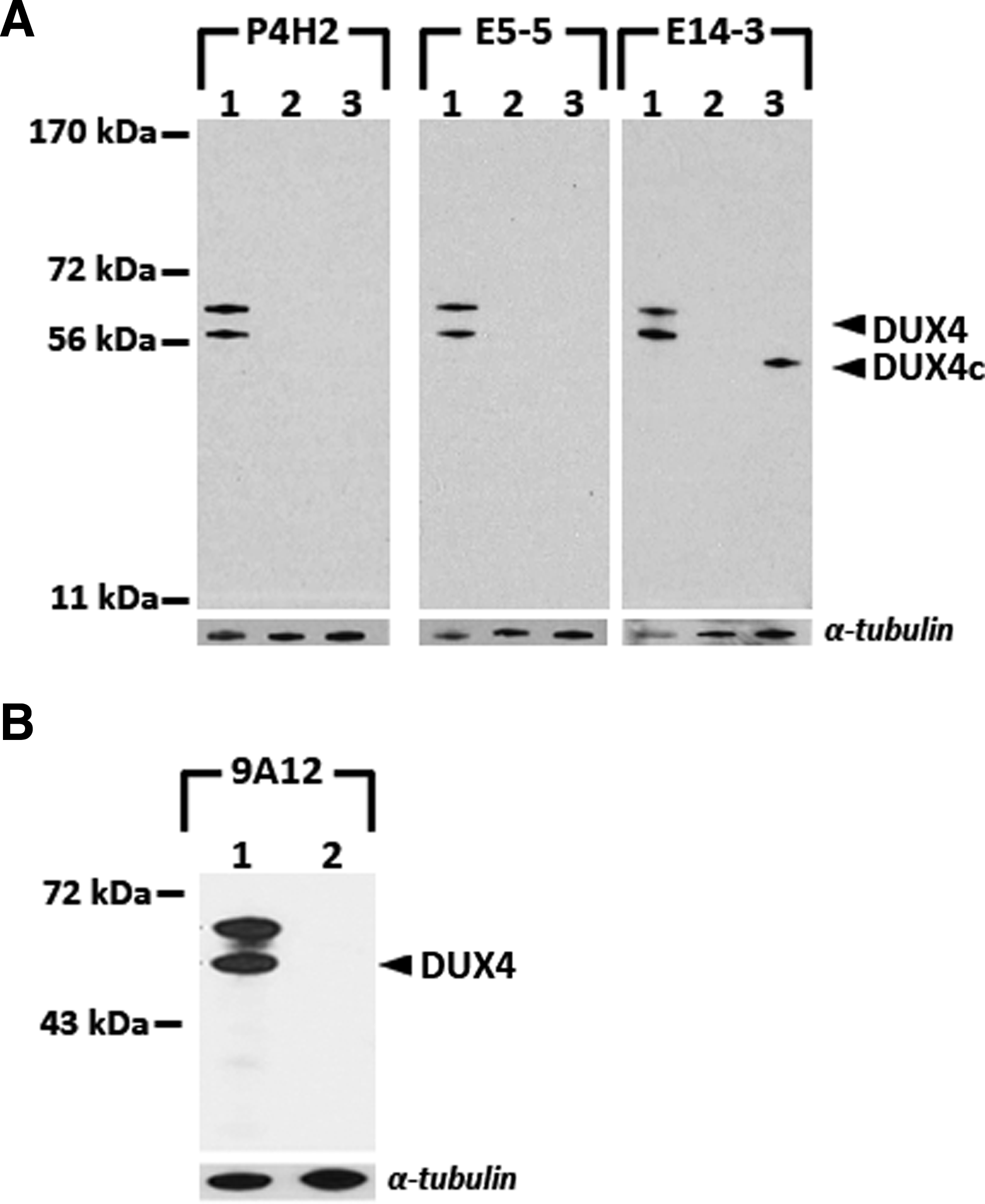
Reactivity of monoclonal antibodies on Western blot analysis. Sample 1, C2C12 cells transfected with pCS2+mkgDUX4(8); sample 2, untransfected C2C12 cells; sample 3, C2C12 cells transfected with pClneo-DUX4c.(10) Blots were probed with the appropriate antibodies and then stripped and reprobed with anti-α-tubulin antibody for loading control. (
C2C12 cells transfected with DUX4 were also fixed and stained with the antibodies. The immunofluorescence analysis using all five monoclonal antibodies demonstrated strong nuclear staining of cells expressing DUX4 that is not seen in neighboring cells, which were not expressing DUX4 (Fig. 3). The immunostaining indicates that these antibodies are able to detect DUX4 protein expressed in the nuclei of mammalian cells, which agrees with previous studies on DUX4's cellular distribution using epitope tag-based experiments.(14) The nuclear localization of DUX4 is consistent with its function as a transcription factor that is able to bind to DNA sequences via its homeodomains and regulate target gene expression.(9,10)
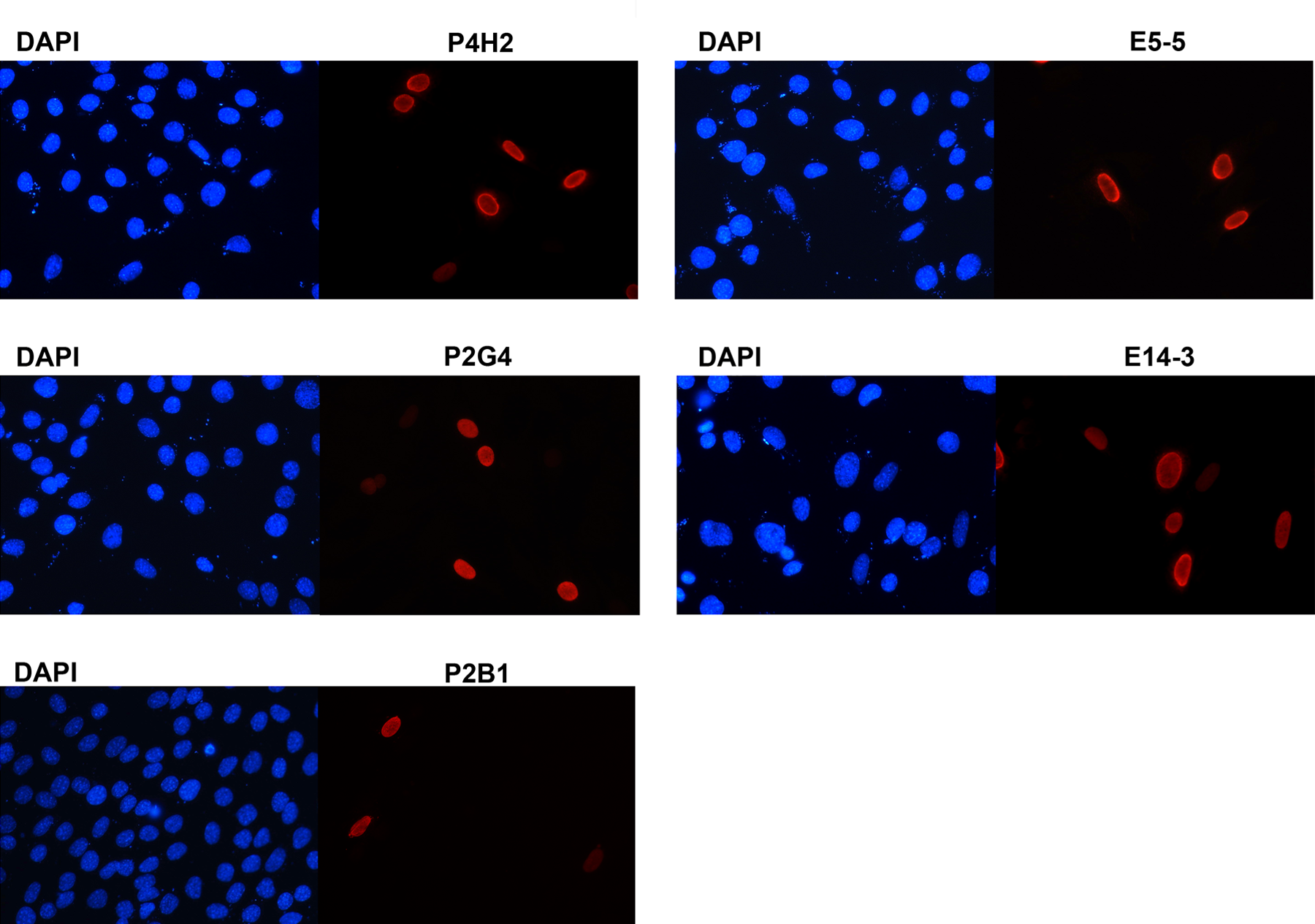
Reactivity of monoclonal antibodies on immunofluorescence. C2C12 myoblasts were transfected with human DUX4 and cells were stained with mouse monoclonal anti-DUX4 antibodies P4H2, P2G4, and P2B1 (left), and rabbit monoclonal anti-DUX4 antibodies E5-5 and E14-3 (right). Cells were counterstained with DAPI for nuclei.
Full length DUX4 induces cell death whereas the shorter splice form does not
To examine the effects of DUX4 when expressed at a lower level, a retroviral vector carrying DUX4 was used to introduce DUX4 into C2C12 myoblasts. At 42 h post-infection, cells were fixed, permeabilized, and co-labeled with mouse monoclonal P4H2 and rabbit monoclonal E14-3. Double-positive DUX4 nuclei on immunofluorescence exhibited a punctate, granular pattern suggestive of chromatin condensation and possibly a pre-apoptotic state (Fig. 4A). Kowaljow and colleagues demonstrated that ectopic overexpression of DUX4 in HEp-2 cells, an epidermoid carcinoma line, results in apoptosis via caspase 3/7 activation just 24 h post-transfection, though dramatically more evident at 48 h.(10) The expression of DUX4 from our retroviral construct is likely closer to endogenous levels, and thus the effects on the murine myoblasts are more subtle, but the results are still indicative of cellular changes associated with the early stages of cell death.
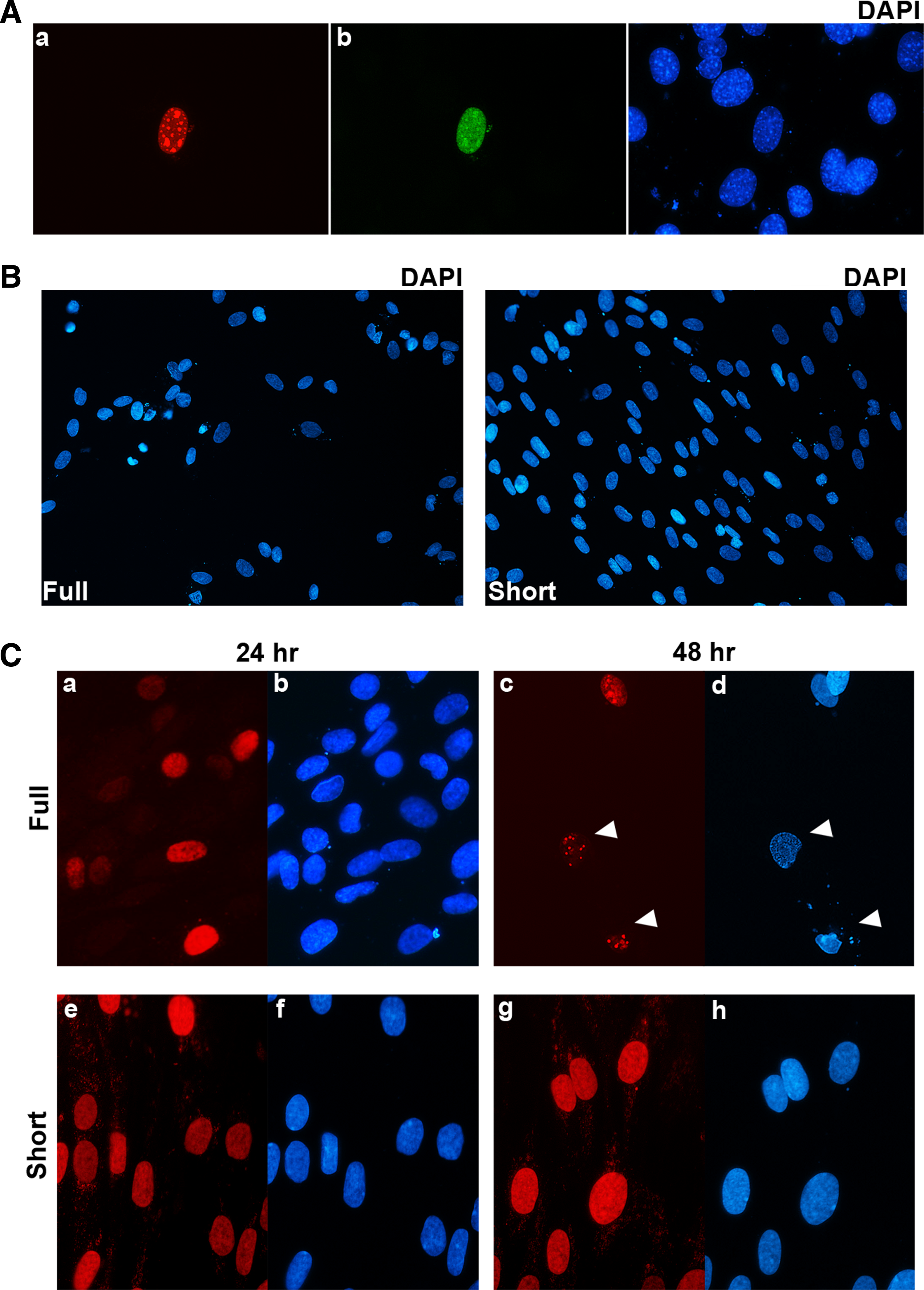
Effects of DUX4 in muscle cells. (
To examine the effects of the different DUX4 splice forms in the context of human muscle cells that would be highly relevant to FSHD, we used a lentiviral delivery system to transduce human primary myoblasts with full length and short DUX4. An equivalent number of myoblasts were infected with either splice form, but 48 h post-infection, more than half the cells were dead on the plates with full length DUX4, whereas there was minimal, if any, cell toxicity was seen on the plates with the short form of DUX4 (Fig. 4B). At 24 h post-infection, both full length and short DUX4 show little effect on cell viability and morphology. However, at 48 h post-transfection, full length DUX4-infected cells exhibited condensation of nuclei and cytoplasm, blebbing of membranes, and fragmentation of cells, features characteristic of apoptosis (Fig. 4B, c,d). The cells infected with short DUX4 remain intact, round and full, and show homogenous staining (Fig. 4B, panels g,h).
Discussion
Few antibodies are currently available against DUX4. Only one polyclonal antibody is commercially available (Santa Cruz Biotechnology, Santa Cruz, CA), and it has not been characterized in the literature to our knowledge. The 9A12 mouse monoclonal antibody developed by Dixit and co-workers has been useful,(10) but it cross-reacts with another highly related protein and candidate disease gene DUX4c, which shares over two thirds of its N-terminal sequence with DUX4.(12,13) We have generated the monoclonal antibodies P4H2, P2B1, and E5-5 specific to the C-terminus sequence of DUX4 that is not present in DUX4c. We also generated monoclonal antibodies P2G4 and E14-3 against the N-terminus of DUX4. Three antibodies, P4H2, E5-5, and E14-3, can detect both denatured DUX4 protein on a Western and fixed native DUX4 protein by immunofluorescence. Antibodies P2G4 and P2B1 work well to detect fixed native DUX4 protein by immunofluorescence, but are less effective for denatured DUX4 on Western blot. These new monoclonal antibodies can be used in combination with other monoclonal and polyclonal antibodies to assess the protein expression of DUX4 in muscle cells affected by FSHD. Additionally, multiple antibody labeling experiments and combination methods such as immunoprecipitation followed by Western blot can be performed with these anti-DUX4 antibodies produced from different species. Development of tools for studying DUX4 protein is critical to advancing the field of FSHD research and for elucidating the molecular pathogenesis of this debilitating disease.
We previously identified alternatively spliced transcripts from DUX4 in FSHD and wild-type muscle cells, including the canonical full length DUX4 that contains the entire coding region of DUX4 as well as a shorter splice form that removes more than two thirds of the coding region.(8) While studies have shown the full length DUX4 to be toxic to cultured cells as mentioned above, there have been no reports on the effects of the shorter splice form on human muscle cells. Here we show that while full length DUX4 exerts overt toxicity on human myoblasts leading to cell death; the short DUX4 shows no obvious negative effects, suggesting that the critical effector component of DUX4 mediating its pro-apoptotic effects is located within the C-terminal region of the protein that is unique to full length form. This is the first demonstration that the different splice forms of DUX4 result in dramatically different effects on human muscle cells. It remains to be investigated whether the short DUX4 performs a beneficial function in muscle cells.
DUX4 is the leading candidate disease gene for FSHD, but the paucity of appropriate molecular tools has greatly limited the study of DUX4 protein expression and function. We have developed five new antibodies that specifically recognize DUX4 by Western blot and immunofluorescence. Immunostaining with the antibodies reveal a differential toxic effect of DUX4 isoforms on skeletal muscle cells and suggest that perhaps only specific splice forms of DUX4 are responsible for the pathogenesis of FSHD. These new tools will provide the opportunity to dissect the role of DUX4 in FSHD by various molecular methods.
Footnotes
Acknowledgments
This work was supported by NIH R01AR045203, NS069539, the Pacific Northwest Friends of FSH Research, and the Shaw Family Foundation. We thank Lauren Snider for reagents and support, Alexandra Belayew for reagents, and the Fred Hutchinson Cancer Research Center Antibody Development Laboratory staff. Mouse monoclonal antibodies will be licensed and made commercially available, for which authors may receive royalties.