
Abstract
Glucosyltransferases GtfB, GtfC, and GtfD of Streptococcus mutans are virulent factors involved in dental caries. Consequently, they are considered to be target molecules in the development of vaccines against dental caries. Among them, GtfD plays a significant role in the sucrose-dependent cellular adhesion of S. mutans, and a number of studies have suggested that the N-terminus of GtfD is an important part of its role in enzymatic activity. In this study, we generated monoclonal antibodies against the N-terminus of GtfD (anti-GtfDN antibody) in an initial attempt to investigate its preventive efficacy against dental caries. To obtain anti-GtfDN monoclonal antibodies, the gene for the N-terminus of gtfD (2 kb) was cloned into an Escherichia coli expression vector, pQE30; then the expressed protein (about 75 kDa) was purified. The purified GtfDN protein was injected into BALB/c mice, and hybridoma clones were established. We obtained three hybridoma clones (HDN9, HDN11, and HDN28) capable of producing anti-GtfDN antibodies. Their binding specificity was characterized by ELISA, dot blot, and Western blot analysis after purification using affinity column chromatography. The isotype of the monoclonal antibodies was confirmed to be IgG2a.
Introduction
To prevent dental plaque, a number of studies have suggested the possibility of controlling dental caries through vaccination, including passive immunization with a previously produced antibody. Because the Gtfs of S. mutans are considered to be essential virulence factors in the formation of dental plaque, they have become target candidates in the development of an effective vaccine. Several studies on Gtfs of Streptococcus mutans have identified the amino acids in the N-terminus of Gtfs of S. mutans to be responsible for the enzymatic activities of sucrose splitting and glucan synthesis,(12–15) while amino acids in the C-terminus have been found to be responsible for glucan binding.(16) A report has also shown that the soluble glucan synthesized by GtfD plays a significant part in the sucrose-dependent cellular adhesion of S. mutans.(17)
We are interested in preventing dental plaque with passive immunization techniques using anti-Gtf antibodies. The purpose of the present study was to generate hybridoma cell lines producing monoclonal antibodies (MAbs) against the N-terminus of GtfD (Gtf-S, 155 kDa).
Materials and Methods
Amplification of gtfD gene
To produce a monoclonal antibody against glucosyltransferase D of S. mutans, the N-terminus of the gene was amplified, sequenced, and inserted into a T vector (Promega Benelux, Leiden, The Netherlands). Polymerase chain reaction (PCR) was performed with GtfD F (5′-GAT CGG ATC CGA AAC AGA ACA GCA GAC CTC AG-3′) and GtfD R (5′-ATC CCA ATC CAT ATG ACT TTT ATC-3′) in a 50 μL mixture containing 1 μL template DNA (about 20 ng), 5 μL of 10 x Tag buffer, 0.5 μL of Tag polymerase (5 U/μL, GeNet Bio, Nonsan, Korea), 3 μL of 2.5 mM dNTP mixture, and autoclaved distilled water. The reaction included initial denaturation at 95°C for 3 min and 30 cycles of denaturation at 95°C for 1 min, annealing at 55°C for 1 min and primer extension at 72°C for 1 min, then final extension at 72°C for 5 min. The amplified DNA fragment was purified and subcloned to a T vector (Real Biotech, Taipei, Taiwan). The sequenced and confirmed N-terminus of the gtfD gene was then cut with SphI and SalI and inserted into a pQE30 protein expression vector.
Bacterial strains and growth conditions
The pQE30 vector that contained the N-terminus of gtfD gene (pGtfDN) was transformed into E. coli M15, and the transformed bacteria were cultured at 37°C overnight in 5 mL of lysogeny broth (LB) broth (1% of trypton, 0.5% of yeast extract, and 1% of NaCl) containing ampicillin (100 μg/mL).
Expression and purification of GtfDN
The following day, 5 mL of LB broth containing M15 was transferred to 500 mL of LB broth containing ampicillin. Expression and purification of the GtfDN protein were preformed according to the protocol in the manufacturer's instructions (Qiagen, Valencia, CA) with some modification. When the culture reached optical density (OD) 0.6–0.8, protein expression was induced by adding isopropylthio-β-D-galactoside (IPTG, 1 mM). Four hours later, the mixture was centrifuged at 5,000 g for 10 min, and the pellet was resuspended in His-binding buffer (20 mM Tris [pH 8.0], 0.5 mM imidazole, and 0.5 M NaCl), and sonicated. It was then centrifuged at 10,000 g for 15 min, and the cleared lysate was loaded into an equilibrated Ni-NTA column with His-binding buffer. It was washed once with wash buffer I (20 mM Tris [pH 8.0], 5 mM imidazole, and 0.5 M NaCl), again with wash buffer II (20 mM Tris [pH 8.0], 5 mM imidazole, and 0.1 M NaCl), and then eluted with elution buffer (20 mM Tris [pH 8.0], 0.2 M imidazole, and 0.1 M NaCl). The size of the eluted protein, GtfDN (about 75 kDa), was confirmed by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). The protein was dialyzed in a dialysis tube in distilled water for 48 h and freeze-dried, then resuspended in phosphate buffered saline (PBS). It was stored at −20°C until use.
Immunization
Four-week-old female BALB/c mice (Damul, Daejeon, Korea) were purchased and raised for 2 weeks before they were injected and immunized three times. The homogeneous emulsion of Freund's complete adjuvant (Sigma Chemical, St. Louis, MO)–GtfDN protein (about 80 μg in PBS) was injected in a 1:1 volume ratio subcutaneously. Two weeks after the first injection, a booster of Freund's incomplete adjuvant was performed, and blood was taken from each mouse 1 week after the second immunization. Serum obtained from the blood was screened at a 1:1000 dilution by Western blot against the GtfDN protein (50 μg/mL in PBS) and stored at −20°C until use. The mouse that showed the highest antibody titer was immunized a third time subcutaneously (80 μg in PBS) with Freund's incomplete adjuvant.
The procedures for the experimental use of animals were approved by the Institutional Animal Care and Use Committee of the Chonbuk National University (approval no. CBU 2009-0014) and followed the guidelines suggested by the committee.
Generation of hybridomas and monoclonal antibodies expressing anti-GtfDN antibodies
On the fourth day after the third injection, the spleen cells of the mouse were harvested and fused with SP2/0 myeloma cells in polyethylene glycol 1500 (PEG 1500, Roche Diagnostics, Mannheim, Germany) in a cell ratio 4:1. After a series of fusion steps, the pellet of the fused cells was diluted in Dulbecco modified Eagle medium (DMEM, Hyclone Laboratories, Logan, UT) containing histone acetyltransferase (HAT) supplemented with 18% fetal bovine serum and then plated onto 96-well microtiter plates (SPL, Pocheon, Korea). The cells were cultured for 10–15 days, and the culture supernatant was assayed for the presence of GtfDN-specific antibodies by ELISA and confirmed by dot blot and Western blot. GtfDN-specific hybridomas were cloned 10 cells/well and then cloned 1 cell/well three more times. The antibody specificity was tested by ELISA, dot blot, and Western blot.
ELISA
The levels of specific anti-GtfDN antibody activity were determined by an enzyme-linked immunosorbent assay (ELISA). Ninety-six-well, flat-bottomed polystyrene microtiter plates (Nunc, Roskilde, Denmark) were used. The wells were coated with GtfDN protein (50 ng/well in coating buffer—0.05 M sodium carbonate and 0.05 M sodium bicarbonate) and incubated overnight at 4°C. Two hundred μL of 5% skim milk were then added, and the wells were incubated for 1 h at room temperature (RT). The wells were washed three times for 5 min each with TBST (100 mM Tris [pH 7.5], 0.1% NaCl, and 0.1% Tween-20) between reactions. The media supernatant, which had the primary antibody (anti-GtfDN antibody) to be tested, was added, and the wells were incubated for 2 h at 37°C followed by the addition of a secondary antibody, alkaline phosphatase-labeled goat anti-mouse immunoglobulin G (Sigma Chemical), and then incubated at 37°C for 1 h. The generation of anti-GtfDN antibody was detected with the substrate p-nitrophenyl phosphate (Sigma Chemical), and the wells were read at 405 nm using an ELISA reader (Packard Instrument, Downers Grove, IL).
Dot blot analysis
The presence of anti-GtfDN antibody can be confirmed by dot blot without performing gel electrophoresis. One or 2 μL (100 ng) of GtfDN was placed on a nitrocellulose membrane and dried. The remaining steps are the same as the protocol of Western blot with some modification. Briefly, the membrane was soaked in 5% skim milk for 1 h at RT and washed three times with TBST between reactions. Culture supernatant was added, and the membrane was then incubated at 37°C for 2 h. A secondary antibody was added, and the membrane remained at 37°C for 1 h. After washing, the presence of anti-GtfDN antibody was detected with 10 mL of TMN (100 mM Tris [pH 9.5], 100 mM NaCl, and 5 mM MgCl2) containing 66 μL of nitro blue tetrazolium (NBT) and 33 μL of 5-bromo-4-chloro-3-indoyl phosphate (BCIP).
Western blot analysis
GtfDN was separated using SDS-PAGE and transferred onto a nitrocellulose membrane for Western blot. The membrane was blocked in 5% skim milk for 1 h at RT and washed three times for 5 min each with TBST. It was soaked in media supernatant for 90 min at RT and washed three times. The positive control was the serum at a 1:1000 dilution (100 μg/mL in PBS), which was treated with secondary antibody for 1 h at RT, then washed three times with TBST for 5 min each. Anti-GtfDN antibody was detected with TMN.
Production of ascitic fluid
The selected final hybridoma cell lines (HDN 9, HDN 11, and HDN 28) were injected into BALB/c mice. Four six-week-old female BALB/c mice were primed with Freund's incomplete adjuvant (300 μL/mouse) and 10 days later, hybridomas of HDN 9, HDN 11, and HDN 28 were harvested, centrifuged, and resuspended in phosphate- buffered saline, and 0.5 mL (2 × 106 cells/mL) was injected into the abdomen of each mouse. Ascitic fluid was collected between 2 and 4 weeks after the injection. Antibody titers were expressed as the reciprocal log2 of the highest sample dilutions that gave an OD405 of 0.09, which was the value of the PBS blank.
Monoclonal antibody purification and determination of antibody isotypes
Monoclonal antibodies were purified using a protein A sepharose column from the culture supernatants of HDN 9, HDN 11, and HDN 28 hybridoma clones, according to the manufacturer's instructions (BioVision, Mountain View, CA) with a slight modification. The column was washed with distilled water to remove air bubbles and filled with protein A beads. The column was then washed with 5x volume of binding buffer (0.05 M sodium borate, 1.5 M glycine, and 1.5 M sodium chloride [pH 8.9]), and the sample was loaded. The column was washed with 5x volume of binding buffer, and IgG was eluted with elution buffer (0.1 M citric acid [pH 2.75]). The isotype of antibodies produced from the stabilized hybridoma cell lines was determined using an isotyping kit (Sigma Chemical) according to the manufacturer's instructions.
Statistical analysis
The mean values and standard deviations were statistically analyzed by Sigmaplot 10.0 (Systat Software, Chicago, IL).
Results
GtfDN protein produced from 2 Kb of the N-terminal half fragment of gtfD gene (Fig. 1) was used as an antigen to immunize mice. We constructed and selected 16 hybridoma clones producing monoclonal antibodies against GtfDN. The specificity of the clones was confirmed through ELISA, dot blot, and Western blot, and three of them, HDN9, HDN11, and HDN 28, showed high affinity to GtfDN and were subjected to further characterization (Fig. 2). Titers of MAbs produced by the selected hybridoma cells in a supernatant of culture media (Fig. 3) and ascitic fluid (Fig. 4) were tested, and the results showed that the binding activity of HDN28 is greater than those of HDN9 and HDN11, which suggested that HDN 28 produced more specific MAbs to GtfDN than HDN9 and HDN11.
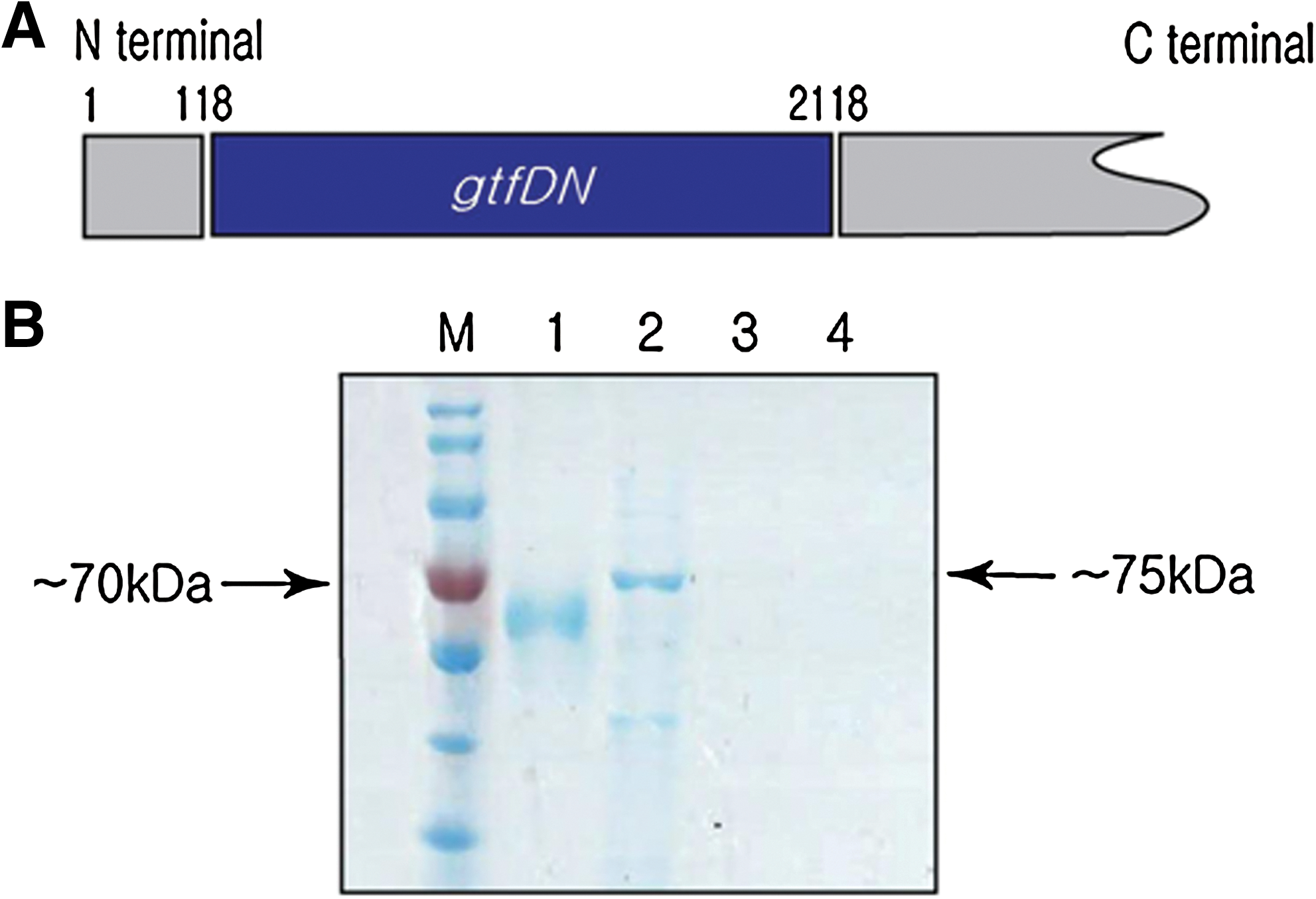
Purification of GtfDN protein. (
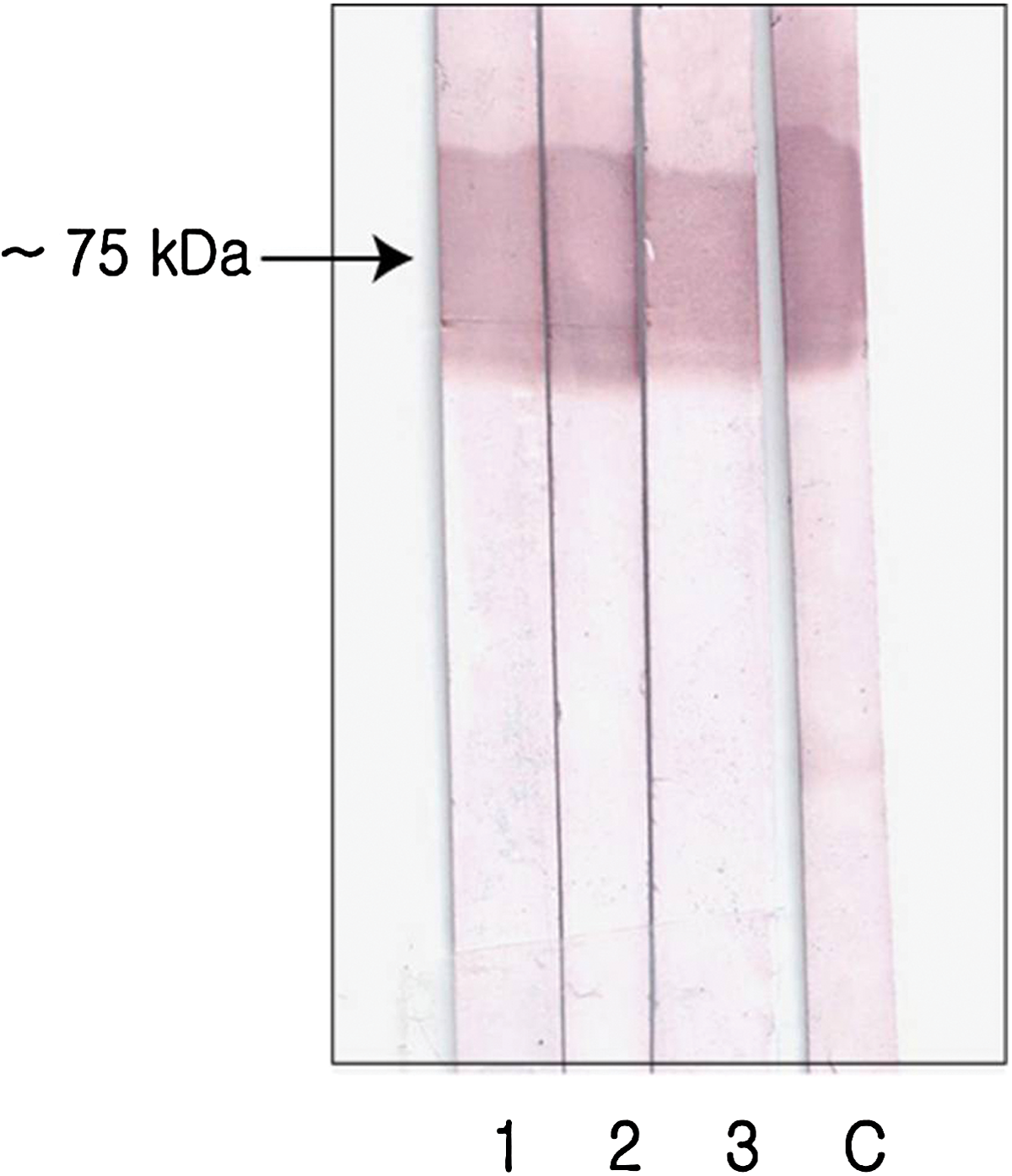
Western blot for fourth selection. Six wells were selected and three showed a strong positive band. Membrane 1, HDN9; membrane 2, HDN11; membrane 3, HDN28 C: control (1:1000 dilution of GtfDN positive serum in PBS).
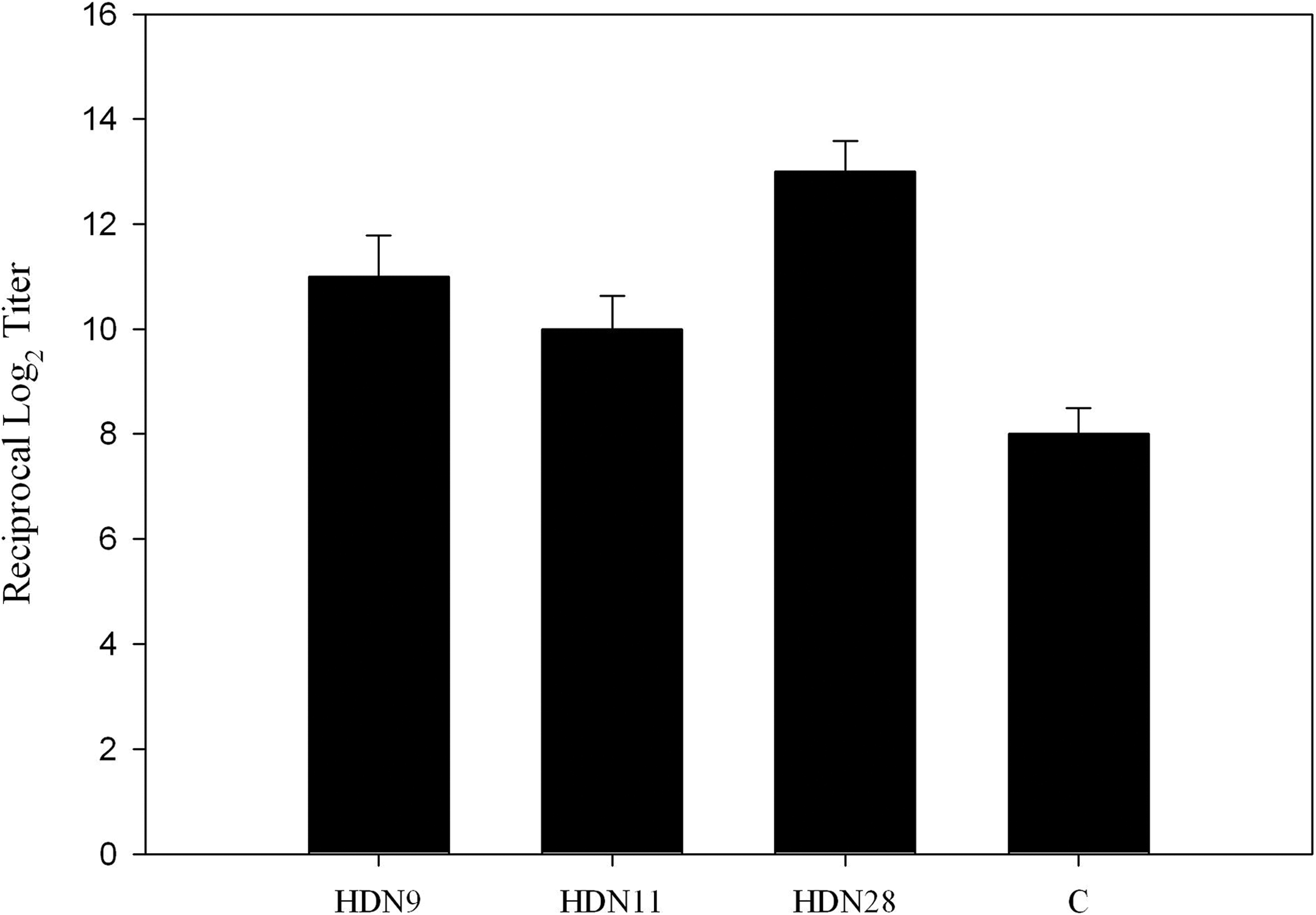
Levels of anti-GtfDN antibodies in culture supernatant of HDN9, HDN11, and HDN28. The control was 1:1000 dilution of GtfDN positive serum in PBS. Data are the mean + SD of triplicate assays.
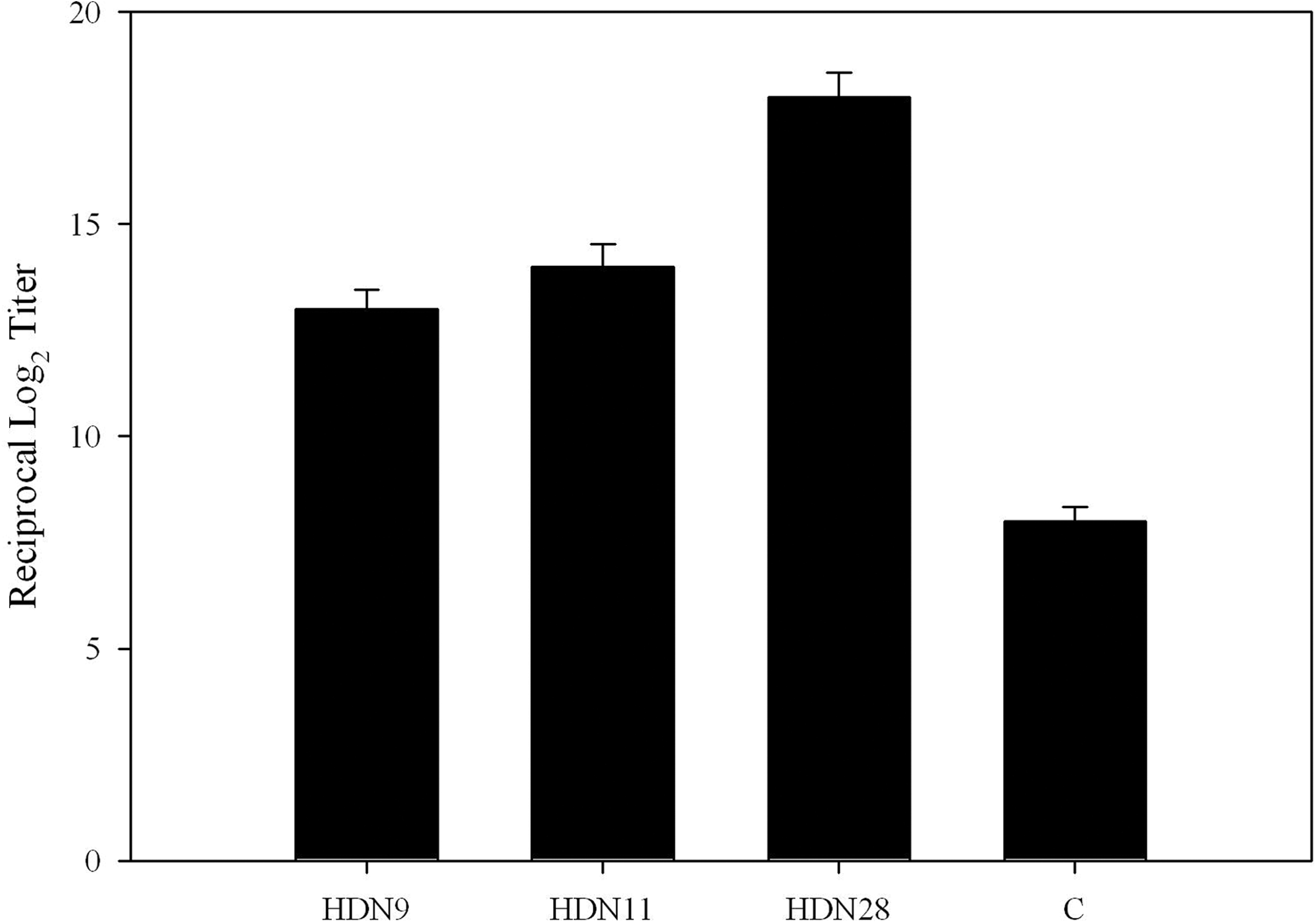
Levels of anti-GtfDN antibodies in ascites fluid of HDN9, HDN11 and HDN28. The control was 1:1000 dilution of GtfDN positive serum in PBS. Data are the mean ± SD of triplicate assays.
Although pristane is normally used to prime mice prior to the injection of hybridoma cells, we used Freund's incomplete adjuvant in this study, because several researchers have shown that Freund's incomplete adjuvant has a better stimulation effect.(18,19) Monoclonal antibody in the culture supernatant was purified using protein A sepharose, but the purification protocol in the manufacturer's instruction manual was modified because we could not get purified IgG suggested by the protocol. We added 1.5 M glycine to the binding buffer and raised the concentration of sodium chloride up to 1.5 M to end up with purified IgG (Fig. 5).
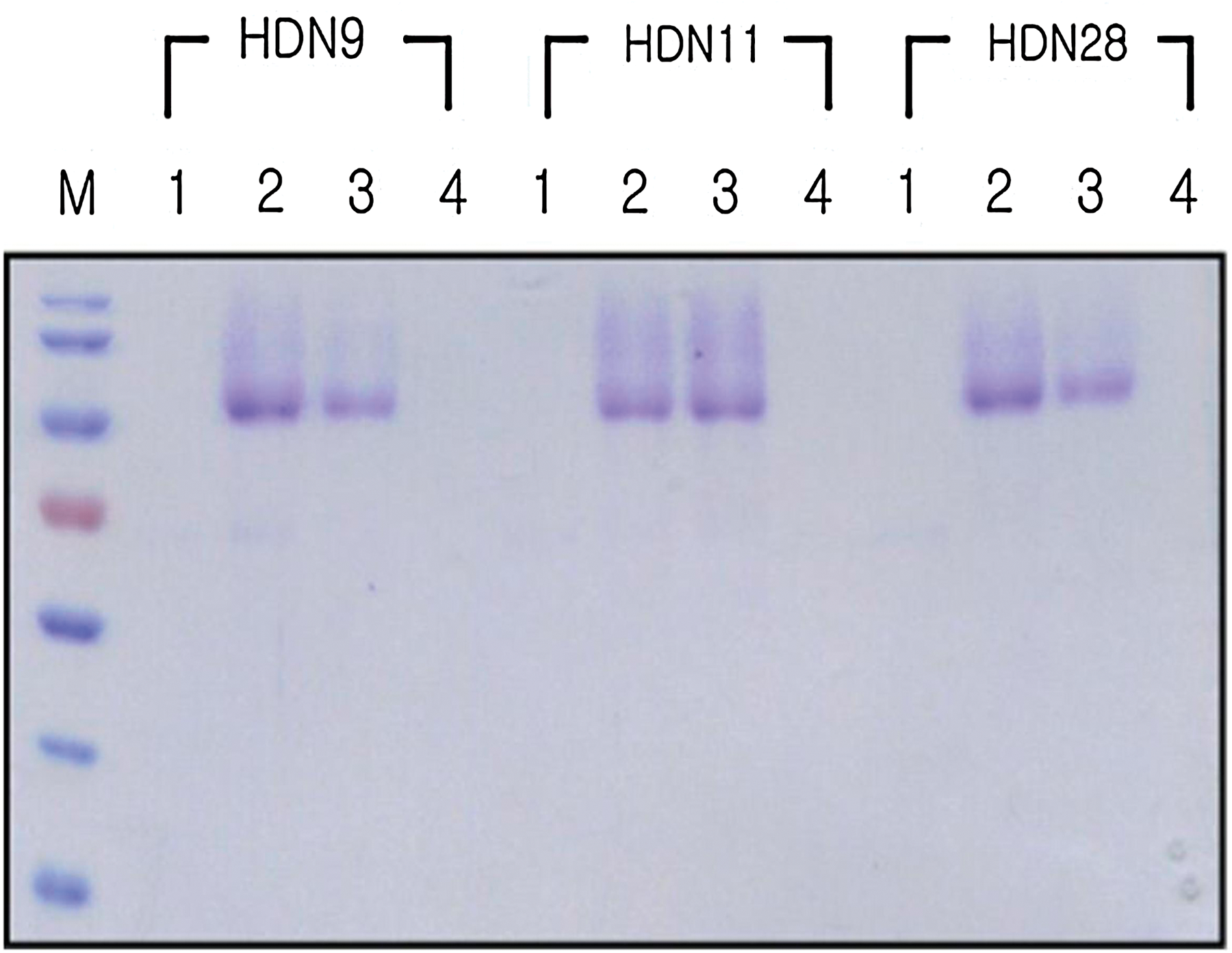
Purification of monoclonal antibody against GtfDN. Monoclonal antibody IgG against GtfDN in culture supernatant was purified using protein A sepharose. M, protein marker; lane 1, elute 1; lane 2, elute 2; lane 3, elute 3; lane 4, elute 4.
During the test of determining the purified IgG specificity to GtfDN protein, the MAbs from each hybridoma clone reacted to GtfDN but not to bovine serum albumin (BSA) (data for HDN11 and HDB28 are not shown) (Fig. 6).
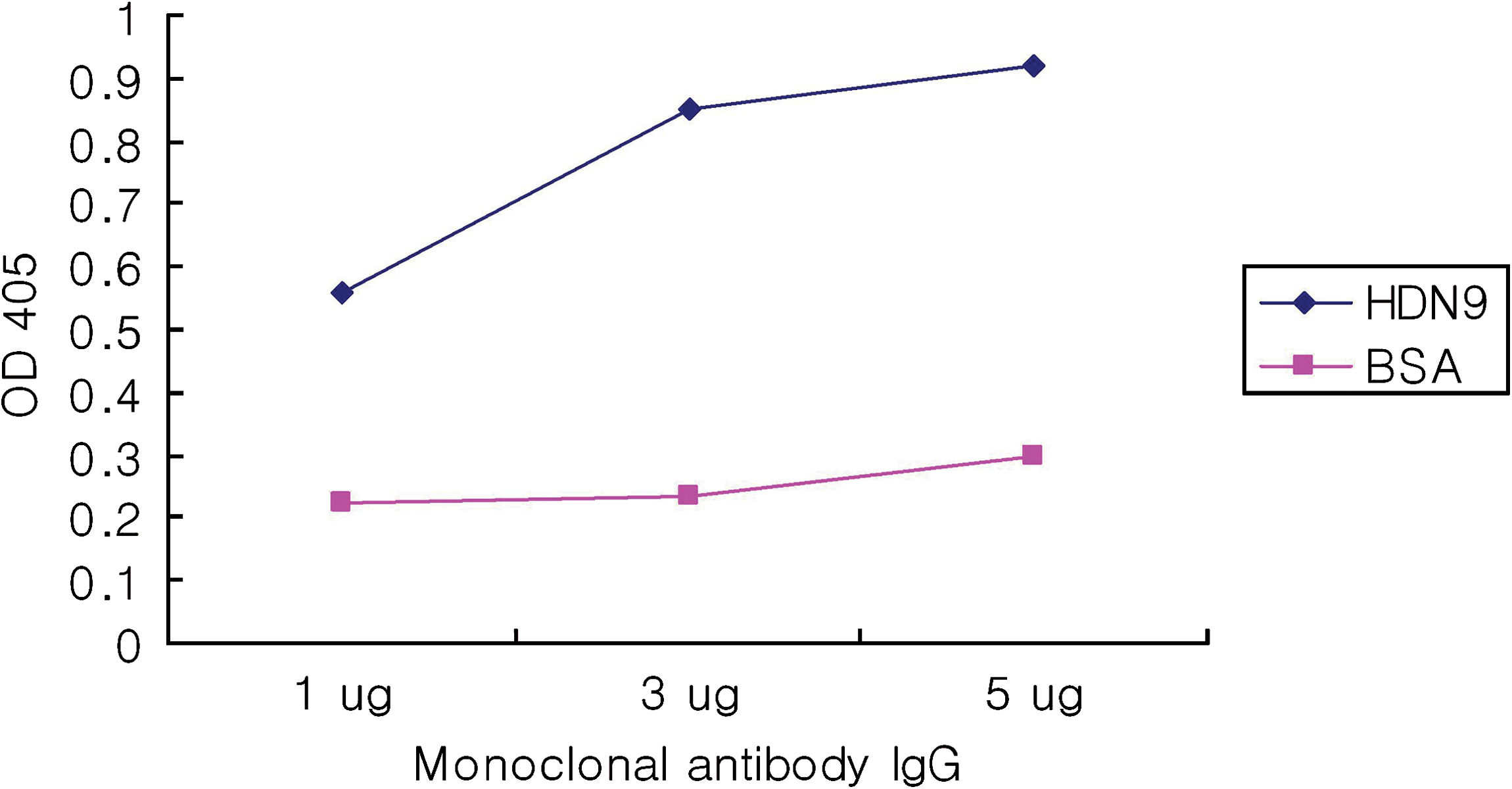
Specificity of monoclonal antibody IgG of HDN9. Antibody at 1 μg, 3 μg, and 5 μg were examined for reactivity with GtfDN and BSA. Data are the mean +SD of triplicate assays.
Discussion
Streptococcus mutans is considered to be a main cause of dental caries, and the pathogenesis of bacteria depends on its ability to accumulate on dental surfaces.(2,10) This process results from the extracellular synthesis of glucans from sucrose by the glucosyltransferases of Streptococcus mutans; these glucosyltransferases have thus been regarded as major virulence factors of S. mutans.(20,21) A study has suggested that an effective way to prevent dental caries is active immunization against S. mutans using the virulence factors of that bacteria.(22) This method has some side effects, however.(23,24) As a result, an alternative approach has been investigated, and several studies have shown that passive immunization using antibodies against S. mutans can be safer than active immunization, since passive immunization works very quickly, lasts for a short time, and has no effect on the systemic immune system.(25–27) Motivated by our interest in passive immunization, we intended to make antibodies against Gtfs, and, as an initial step, we focused on GtfD, which is thought to be indispensable for S. mutans colonization on tooth surfaces.(21) In the present study we made hybridoma cells producing anti-GtfDN monoclonal antibodies.
In the process of generating hybridoma cells, we found that the fusion of spleen cells and myeloma cells on the fourth or sixth day after the third immunization provides better results than the usual fusion on the third day (data not shown). We assume that this is because the state of boosting is more stable and stronger on the fourth or sixth day than on the third day. The GtfDN protein that was made in this study was from a recombinant plasmid, and its glucosyltransferase enzyme activity should be investigated in our next study. Also, further research will be needed to assay whether anti-GtfDN antibodies are an effective tool to control dental caries.
Footnotes
Acknowledgment
This study was supported by a grant from the National Research Foundation of Korea.
Author Disclosure Statement
The authors have no financial interests to disclose.