
Abstract
PEX5 is a key protein of the peroxisomal protein import machinery. This cycling receptor binds newly synthesized proteins with a peroxisomal targeting signal type 1 in the cytosol and directs them to the peroxisomal membrane. There, PEX5, together with its docking protein, forms a transient membrane-spanning channel that enables cargo-transport across the membrane. Through interaction with other multimeric membrane complexes, the receptor is released from the membrane back to the cytosol. Very little is known about the various conformational states of the receptor during its cycling. Here we report the generation and characterization of a mouse monoclonal antibody that recognizes in vivo primarily the membrane-associated form of the human PTS1 receptor.
Introduction
The cycling PTS1 receptor PEX5 is the central player within the cascade of intracellular events facilitating matrix protein import into peroxisomes.(3–5) In human cells, two isoforms––PEX5L (long isoform) and PEX5S (short isoform), which differ in length by 37 amino acids, are expressed in similar amounts. In their cytosolic states, both soluble isoforms recognize newly synthesized peroxisomal proteins via their carboxy-terminal targeting sequence PTS1 and target them to the peroxisomal membrane.(6) After docking at the peroxisomal membrane protein PEX14, the receptor becomes an integral constituent of a flexible and highly dynamic pore that allows translocation of the cargo protein.(7) A recycling process that involves ubiquitination and ATPase-dependent action of two AAA-peroxins, PEX1 and PEX6, results in release of the receptor from the membrane into the cytosol to enable another round of protein import.(8,9)
The unique mechanism underlying peroxisomal matrix protein import is not understood in detail. Several of the unsolved central questions are related to topology of the membrane-associated form of the receptor. It will be of high interest to gain insight into the architecture of the transient translocation pore, which accommodates fully folded, even oligomerized proteins. Another question concerns the dynamic properties of the PTS1 receptor enabling a switch between a soluble cytosolic and an integral membrane conformation during the cycle. So far, antisera raised against human PEX5 were not applicable to detect the peroxisome-bound form of the receptor in normal cells by immunofluorescence microscopy due to the strong fluorescence background caused by the primarily cytosolic distribution of the PTS1 receptor.(10) For immunodetection of peroxisome-associated PEX5, either removal of cytosol or genetic or biochemical manipulations, which lead to arrest of the receptor cycle,(11) had to be carried out.
In the present study, we report on the generation and characterization of a monoclonal antibody raised against human PTS1 receptor PEX5, referred to as MAb 9D12. In contrast to other available PEX5-specific antisera and monoclonal antibodies, MAb 9D12 marks primarily the membrane-embedded form of the human receptor and therefore provides a tool with great potential for analyzing the receptor cycle in normal and Zellweger patient cell lines.
Materials and Methods
Expression and purification of recombinant PTS1 receptor
Human His6-tagged PEX5L was expressed from a modified pET24d vector (G. Stier, EMBL-Heidelberg, Germany) in Escherichia coli strain BL21(DE3). Cells were induced in the mid-log phase with 0.4 mM isopropyl-β-D-thiogalactopyranoside and grown for another 4 h at a temperature of 30°C. Cells were broken with a French pressure cell (Thermo Fisher Scientific, Waltham, MA) in one or two passages in buffer containing 20 mM Tris-HCl [pH 8.0], 150 mM NaCl, and 50 mM imidazole. Cleared lysates were loaded to a HisTrap™ HP-Column (GE Healthcare, Freiburg, Germany) and eluted with 500 mM imidazole. After subsequent loading to a ResourceQ™ Anion exchange column (GE Healthcare) and washing with buffer containing 20 mM Tris-HCl [pH 8.0], PEX5 was finally eluted with a 20 mL linear gradient between 0 and 500 mM NaCl. All purification steps were carried out at 4°C and monitored by SDS-PAGE analysis.
Immunization of mice and production of monoclonal antibodies and purification
A female Balb/c mouse was subcutaneously injected with 0.1 mg of PEX5 antigen in PBS and Freund's complete adjuvant. Animals were boosted four times with 0.1 mg antigen each and serum-antibody-titer was measured by ELISA. Three days after the fourth boost, mouse spleen cells were PEG-fused with an equivalent number of vital cells of the HGPRT-deficient mouse-myeloma-line P3-X63-Ag8.653 (ATCC no. CRL-1580), suspended in HAT-selection medium, supplemented with 20% FCS (PAN Biotech, Aidenbach, Germany), and seeded in 96-well cell culture plates.
Aminopterine was added for 14 days (HAT supplement), and hypoxanthine and thymidine for a further 7 days (HT supplement) to the selection medium. After selection, the remaining hybridoma cells were cultivated in RPMI1640/glutamine (PAA Laboratories, Pasching, Austria) supplemented with 10% FCS. For MAb enrichment, cell culture supernatant was subjected to ammonium sulfate precipitation ranging from 20 to 50% (w/v).
ELISA
ELISA tests were performed on Costar High Binding 96-well microtiter plates. Fifty μL of PEX5 antigen (5 μg/mL PBS/sodiumazide) were adsorbed onto the surface of each well by overnight incubation at 4°C. Remaining free binding capacities on the well surface were blocked with bovine serum albumin (BSA) solution (3% w/v in PBS/ sodiumazide, 200 μL/well) for 12 h at 4°C. After washing with H2O, 50 μL cell culture supernatants were transferred into the wells and incubated for 1 h at 37°C. After subsequent washing with H2O, wells were incubated with 50 μL alkaline phosphatase-conjugated goat anti-mouse IgG antibody (Jackson ImmunoResearch Labs, Suffolk, United Kingdom), diluted 1:5000 with PBS/1% BSA, for 45 min at 37°C. After subsequent washing with H2O, p-nitrophenyl-phosphate substrate (1 mg/mL in DEA buffer [0.8 M diethanolamine, 0.5 mM MgCl2 [pH 9.8]), 200 μL/well) was added and incubated for 30 min at room temperature. The optical density (OD) was measured in a 2-beam plate-photometer (Tecan SpectraFluor, Männedorf, Switzerland) at 405 nm.
Epitope mapping
Peptides comprising 15 amino acids and representing the full peptide sequence of human PEX5L were directly synthesized on a cellulose membrane as described previously.(12) The membrane was blocked for 1 h at room temperature with 5% skim milk in PBS-ST (10 mM Na2HPO4, 1.8 mM KH2PO4, 140 mM NaCl, 2.7 mM KCl, 0.1% [v/v] Triton, 0.02% [v/v] SDS [pH 7.3]). After overnight incubation at 4°C with MAb 9D12 and subsequent washing, epitopes were immunochemically detected with horseradish peroxidase–coupled secondary antibodies in PBS-ST and ECL Immunoblotting Detection Reagent (GE Healthcare).
Immunoblotting
Purified His-tagged HsPEX5L and cell lysates of GM5756T fibroblasts were separated by 12.5% SDS-PAGE and subsequently transferred to nitrocellulose membranes. Blocking of membranes was achieved by 1 h incubation in 5% skim milk in PBS-ST. Membranes were incubated with monoclonal antibodies overnight in PBS-ST at room temperature. After washing with PBS-ST, membranes were incubated for 1 h at room temperature with anti-mouse IgG-conjugated IR-Dye (Li-Cor Biosciences, Lincoln, NE) followed by two times washing and subsequent detection and processing by Odyssey Imaging System (Li-Cor).
Immunofluorescence microscopy
Human fibroblast cells were cultured at 37°C in Dulbecco's modified Eagle's medium supplemented with 10% fetal calf serum, 2 mM L-glutamine, 100 U/L penicillin, and 100 mg/L streptomycin at 8% CO2. Patient-derived PEX6-deficient skin fibroblasts were kindly donated by R. Wanders (AMC, Amsterdam, The Netherlands) and immortalized by transfecting with SV-40 large T antigen. The resulting cell line ΔPEX6T, PEX19-deficient ΔPEX19T cells,(13) and normal fibroblasts (GM5756T) were grown for 1 day on cover slides in 60 mm tissue culture dishes, fixed with 3% formaldehyde in PBS, permeabilized with 1% Triton X-100 in PBS, and subjected to immunofluorescence microscopy. Whereas different monoclonal mouse antibodies were used for labeling of different conformational states of PEX5, polyclonal rabbit antiserum against HsPEX14(14) was used as peroxisomal marker. Secondary antibodies were conjugated with Alexa Fluor-594 or 488 (Invitrogen, Carlsbad, CA). All micrographs were recorded on a Zeiss Axioplan 2 microscope with a Zeiss Plan-Apochromat 633/1.4 oil objective and an Axiocam MR digital camera and were processed with AxioVision 4.2 software (Zeiss, Jena, Germany).
Results and Discussion
The PTS1 receptor in human fibroblasts has a dual intracellular localization in human fibroblasts. While the majority are distributed within the cytosol, only a small portion, less than 5%, is associated with the peroxisomal membrane.(10) Due to the high cytosolic background, the membrane-embedded state is normally not detectable by standard immunocytochemical techniques and polyclonal antibodies against PEX5.(10,11)
We have raised a set of monoclonal antibodies against human PEX5 and characterized them by immunoblotting and in vivo localization studies. All MAbs including the ones produced from clone 9D12 (MAb 9D12) specifically detected both isoforms of the PTS1 receptor, PEX5S and PEX5L, in human fibroblast lysates under denaturing conditions (Fig. 1). To test the suitability of MAbs for localization studies, we performed immunofluorescence microscopy in human fibroblasts. Among all tested antibodies, only MAb 9D12 labeled dot-like structures (Fig. 2). Such punctate cell pattern is typical for a peroxisomal staining. Peroxisomal co-localization was verified by the congruent fluorescent pattern obtained with antibodies against the peroxisomal membrane protein PEX14 (Fig. 2).
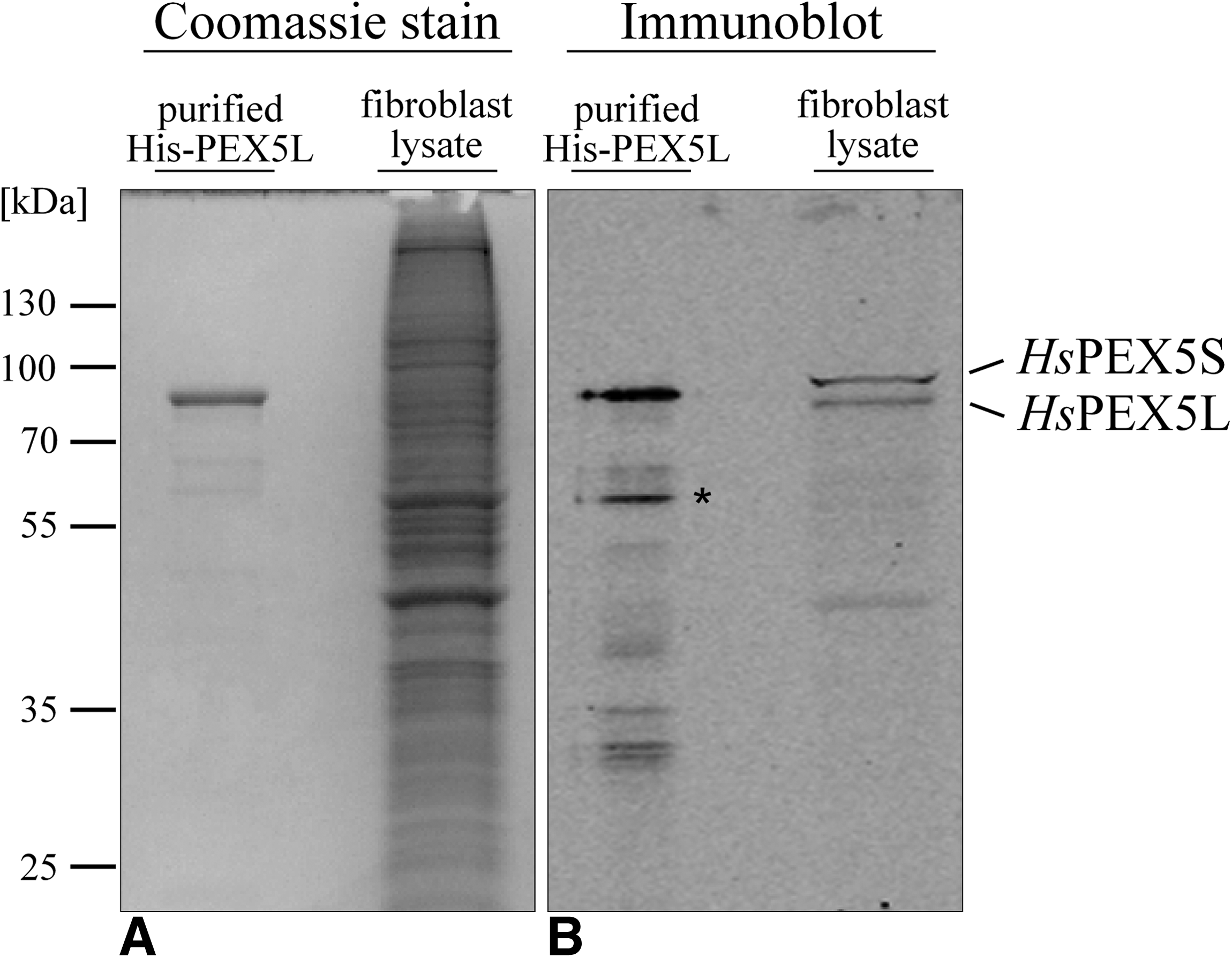
Specificity of monoclonal antibody (clone 9D12) under denaturing conditions. 600 ng of purified recombinant His-tagged HsPEX5L (His-PEX5L) and 4 μg total protein of human fibroblast lysate were separated by 12.5% polyacrylamide SDS gel electrophoresis and stained with Coomassie blue (
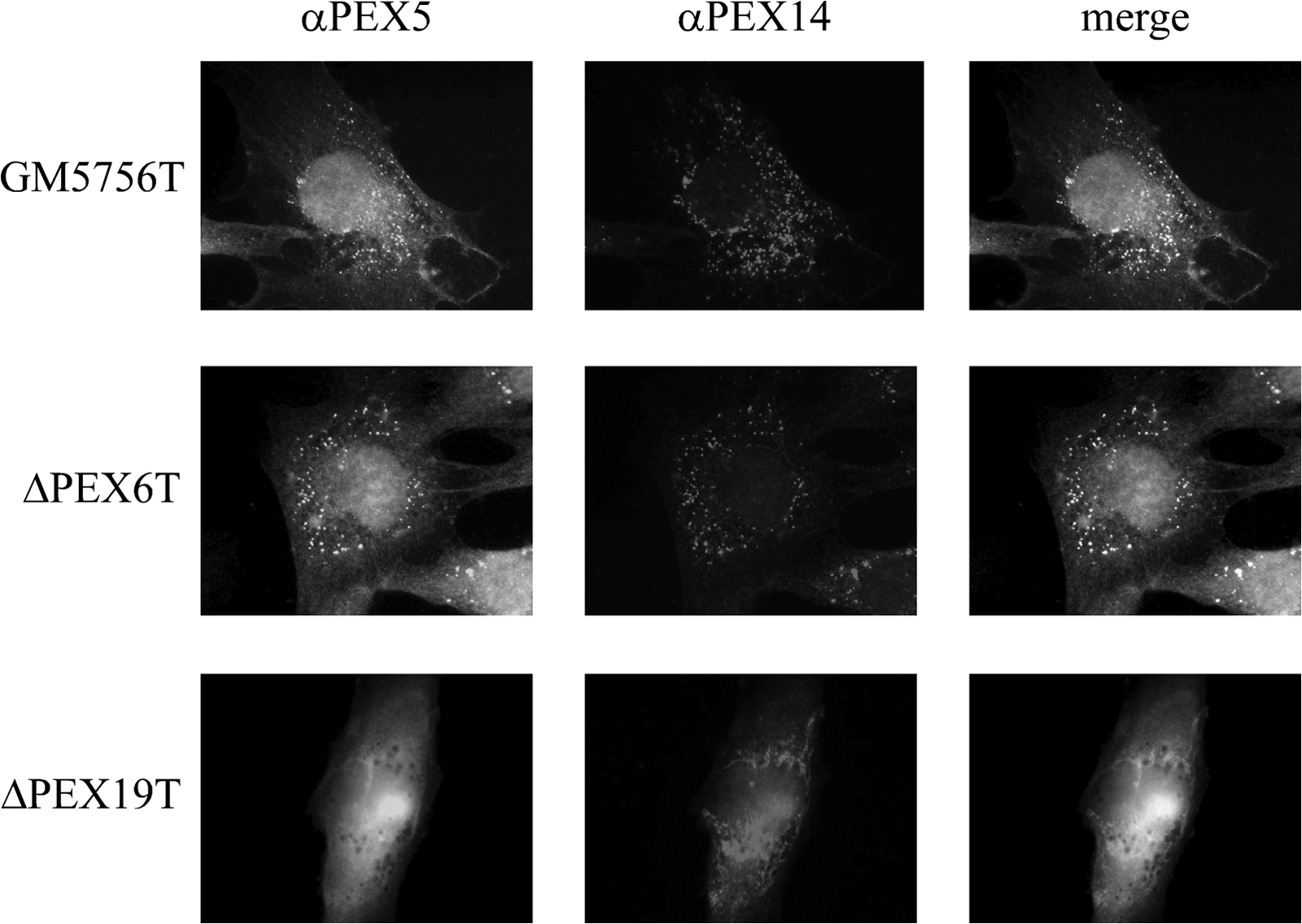
Immunofluorescence microscopy with monoclonal antibody (clone 9D12). In human fibroblasts (GM5756T) PEX5 was stained indirectly with Alexa Fluor-488 labeled second antibody via MAb 9D12 (αPEX5). Peroxisomal localization was confirmed with PEX14 antiserum (αPEX14) and Alexa Fluor-594 labeled second antibody. PEX6 and PEX19 deficient strains (ΔPEX6T and ΔPEX19T) were used as controls.
To further verify that the antigenic determinant at the peroxisome is indeed the membrane-bound state of the PTS1 receptor, immunofluorescence studies were performed in fibroblasts of Zellweger patient cell lines. PEX19 is the receptor for peroxisomal membrane proteins, and therefore deficiency results in a complete absence of intact peroxisomal membranes.(15) Accordingly, PEX5 remains in the cytosol in PEX19-deficient cells as indicated by the lack of punctate staining obtained with MAb 9D12 (Fig. 2). In contrast, a more pronounced immunofluorescence signal at peroxisomes could be observed in PEX6-deficient cells. PEX6 is one of the two AAA-peroxins that is required for displacement of PEX5 from the peroxisomal membrane. As shown for yeast and mammalian cells,(8,16) the PTS1 receptor accumulates at the peroxisomal membrane in such cells.
Membrane insertion of PEX5 as shown for human and yeast counterparts(17,18) is an enigmatic process that might resemble the action of the so-called pore-forming toxins.(19) It seems clear that the N-terminal half of the receptor is involved in this process whereas the C-terminal half is required to bind the PTS1 (Fig. 3A). It has been shown that the N-terminal half of the soluble form is mostly unstructured.(20) No structural information is available on the membrane-embedded form but, in analogy to the action of pore-forming toxins, it has been suggested that regions within the N-terminal half are able to form amphipathic helices that could traverse the membrane to build the transient pore.(19) We performed peptide scan analysis to map the dominant epitopic region to amino acids QDYNET (amino acids 133-138) (Fig. 3B). Our results strongly suggest that the epitope located within the N-terminal half (Fig. 3A) is even more accessible in the membrane-embedded than in the cytosolic form. This result supports the notion that the receptor undergoes major conformational changes after docking at the peroxisomal membrane.
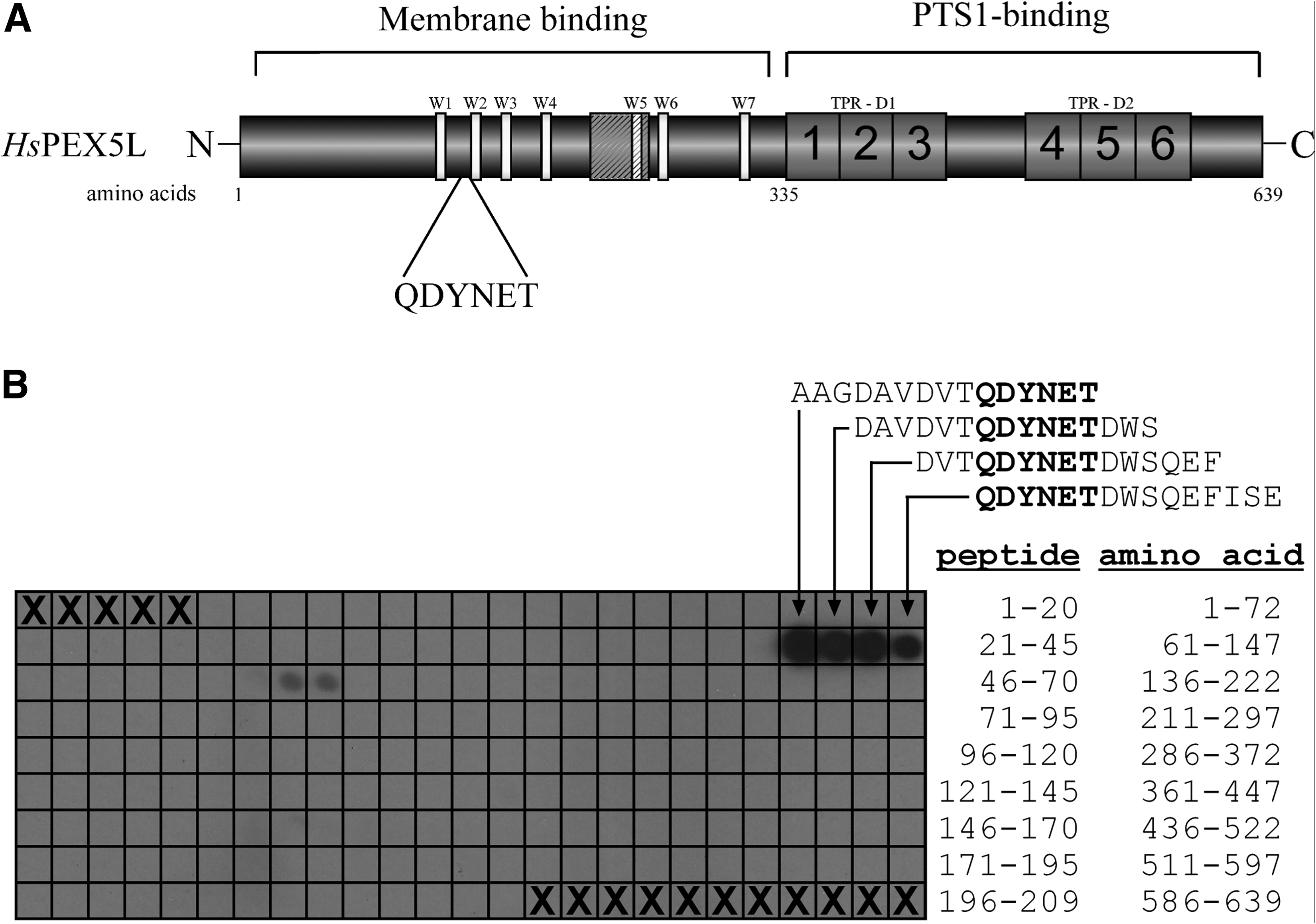
PEX5 epitope mapping of monoclonal antibody (clone 9D12). (
In future studies, MAb 9D12 will be a valuable tool to elucidate the localization and topology of membrane-bound states of PEX5 during receptor cycling. Such states could be arrested in Zellweger patient cell lines or can be provoked by site-directed mutagenesis of the PTS1 receptor.
Footnotes
Acknowledgments
We thank Elisabeth Becker for technical assistance. This work was supported by grants from the European Commission (LSHG-CT-2004-512018) and the Deutsche Forschungsgemeinschaft (SFB642).
Author Disclosure Statement
The authors have no financial interests to disclose.