
Abstract
Fpr3 is the third member of the FKBP (FK506 binding protein) family in yeast. In this study, the fpr3 gene from Saccharomyces cerevisiae was overexpressed and the protein product was purified using different methods. The recombinant Fpr3 fusion protein (rFpr3) was then used as antigen to immunize BALB/c mice for the production of monoclonal antibodies (MAb). Western blot and ELISA results indicated that rFpr3 had specific binding ability to the MAbs, and isotyping results classified the MAb as the subclass IgG1 by antibody. The MAbs obtained in this study will be used as a molecular chaperone to obtain Fpr3 crystals.
Introduction
In structural analysis, there are several X-ray and NMR (nuclear magnetic resonance) structures of FKBPs, with FKBP12 being the prototype of the family and FKBP73 being extensively studied free or in complex with small ligands.(6,7) However, there are no solved crystal structures compatible with the important roles in the biochemical processes of Fpr3. The main reason for this is that the crystals of Fpr3 are difficult to obtain due to the protein containing poorly ordered regions. To this point, the MAbs of the protein were usually used as a molecular chaperone to obtain the ordered structures of the protein. Therefore, we overexpressed the Fpr3 in E. coli and purified the rFpr3 with a high purity. The purified rFpr3 protein was then used in ELISA as antigens to immunize BALB/c mice for the production of monoclonal antibodies with a high titer. Finally, two hybridoma cell lines screening MAbs against Fpr3 were generated.
Materials and Methods
Cell culture and preparation of Fpr3 protein
SP2/0 cells were maintained in RPMI-1640 with 10% fetal calf serum. The cell line and the later prepared hybridomas were maintained in a humidified chamber with 5% CO2 at 37°C. The fpr3 gene was amplified by standard PCR method from yeast and constructed into a pEGX-5X-1 vector to give a recombinant expression plasmid as described previously.(8) E. coli BL21(DE3) carrying the recombinant expression plasmid was grown in LB media supplemented with appropriate antibiotics at 37°C to A600=0.6, and induced with 0.8 mmol/L IPTG (isopropyl-1-thio-β-D–galactopyranoside) at 30°C for 6 h. Recombinant Fpr3 fusion protein with GST-tag was expressed as a soluble protein after induction. The protein was purified by GST affinity chromatography and Superdex 200 gel filtration chromatography, and then verified by SDS-PAGE.
ELISA screening
To select for positive hybridomas, 96-well plates were coated with 5 μg/mL rFpr3 at 100 μL/well and incubated at 4°C overnight. The plates were then washed three times with 0.05% Tween-20/PBS and blocked for 2 h with blocking solution. The blocking solution was removed and 100 μL of culture supernatant was added and incubated for 1 h at 37°C. Next, plates were washed with 0.05% Tween-20/PBS, and goat-anti mouse IgG-HRP conjugate diluted 1:10,000 in blocking solution was added for 1 h at 37°C. After washing three times in Tween-20/PBS, ELISAs were developed using 100 μL of Zymed TMB substrate (Invitrogen, Carlsbad, CA), stopped with 100 μL of 2 M H2SO4 and read at 492 nm.
Characterization of MAbs and Western blot and ELISA analyses of purified MAbs
Isotyping of MAbs was carried out with ELISA utilizing specific antibody class/subclass (Sigma-Aldrich, St. Louis, MO). The MAb was then purified with Protein G Sepharose 4B resin (GE Healthcare, Bellefonte, PA) using an established method according to the immunoglobulin isotype. The specificity of the MAbs were evaluated by Western blot and ELISA methods described above with rFpr3, while the same molar concentration of BL21(DE3) cell lysate containing pEGX-5X-1 was used as negative control. Western blot analysis of the MAbs was performed using the protocol of TE 22 Mini Tank Transfer Unit (GE Healthcare Life Sciences, Little Chalfont, United Kingdom).
Results
Preparation of rFpr3
After induction, the crude proteins of E. coli were separated by SDS-PAGE to examine whether the rFpr3 were expressed. As shown in Figure 1A, the rFpr3 were expressed as an expected molecular weight of 90 kDa (the molecular mass of rFpr3 is 70 kDa and the molecular mass of GST is 20 kDa). After being purified with GST affinity chromatography and Superdex 200 gel filtration chromatography, the protein was found with a high purity (Fig. 1A, lane 2).
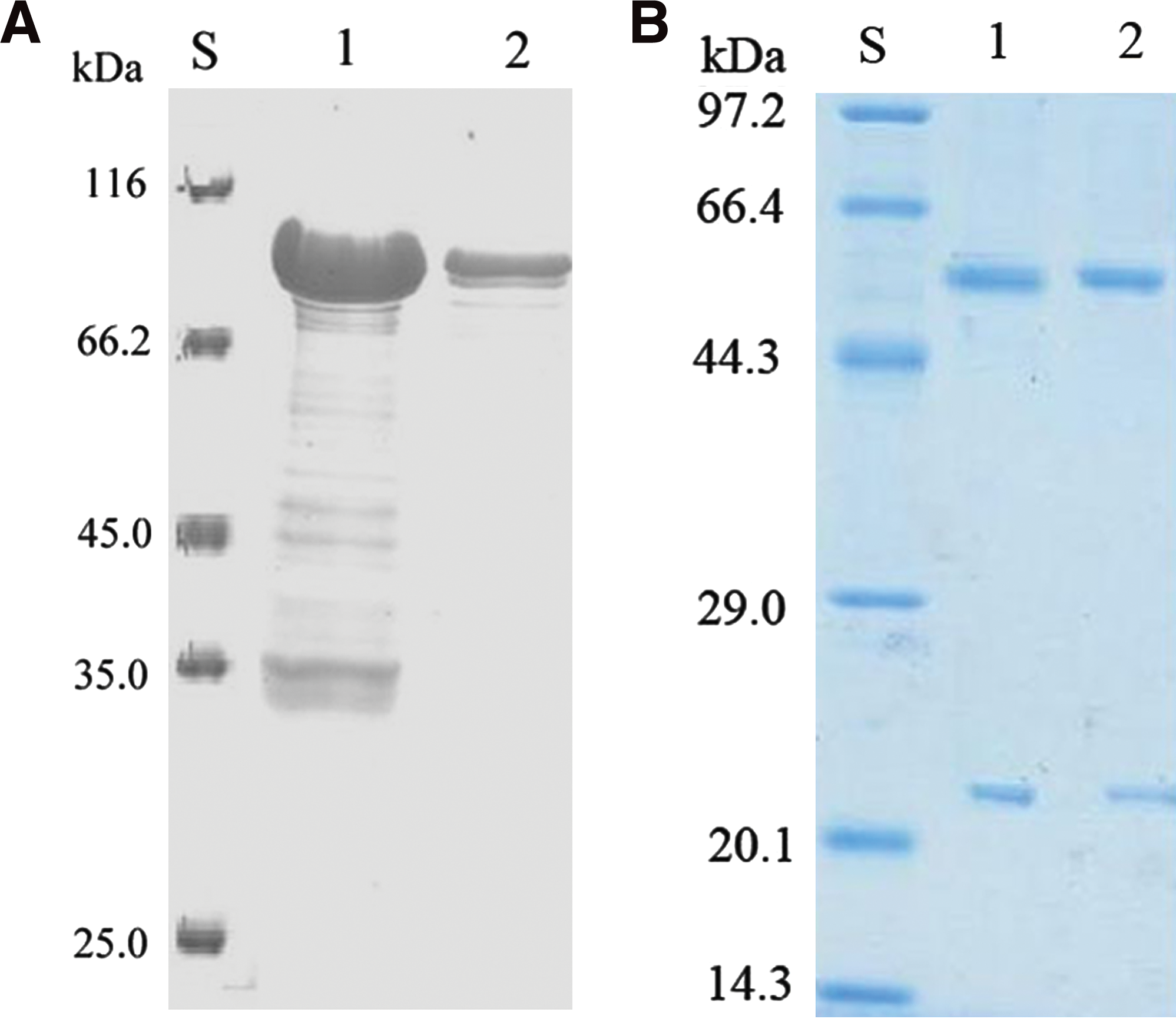
(
Preparation of hybridoma cell lines, screening, and characterization of MAbs
At the end of the immunization period, the spleen of the mouse that gave the highest antibody response was selected as a fusion partner in conventional hybridoma technology. After 14 days, supernatants were screened using an indirect ELISA to detect hybridomas secreting specific antibodies against rFpr3. The screening procedure resulted in two hybrid clones reactive against rFpr3. Finally, one hybridoma was selected for the production of MAbs following subcloning by limiting dilution method. After purification, the concentration of the MAbs was 1.7 mg/mL (Fig. 2, lanes 1, 2). Isotyping results classified the MAb as the subclass IgG1 by antibody.

Western blot analysis of the purified MAbs. S, size standard (pre-stained; lanes 1, 2, the recombinant of GST-Fpr3; lanes 3, 4, BL21(DE3) cell lysate containing pEGX-5X-1.
The specificity of the MAbs obtained was further investigated by Western blot and ELISA. In Western blot, signals at approximately 90 kDa were observed, consistent with the expected molecular weight of the recombinant proteins (Fig. 2, lanes 1, 2). The results showed that the MAbs were specifically recognized against rFpr3 but had no cross-reactivity with GST (Fig. 2, lanes 3, 4). The affinity of the MAbs to rFpr3 was measured by the ELISA described using serial dilutions of MAbs. The results showed that the purified MAbs had a high titer with the proteins.
Discussion
Fpr3 as an FK520-binding protein is the first proline isomerase,(9) which has two domains: a highly charged NH2 terminus that is suggested to be involved in the nuclear import of the accumulated hybrid proteins(9) required for ribosome biogenesis, and a COOH terminus with homology to yeast proline isomerases.(9,10) In recent studies, Fpr3 was also found to play a critical role in protein folding and assembly,(11) cell growth, viability, and other cellular functions.(12) However, there were no reports of the crystal structures of Fpr3.
For elucidating the mechanism of the Fpr3 in diverse cellular processes, we used the purified rFpr3 as antigen to immunize BALB/c mice for the production of MAbs. Western blot and ELISA results showed that the MAbs obtained in this study have a high affinity, stability, and specificity. Therefore, the MAbs obtained in this study will provide another useful tool in the functional study of Fpr3. Furthermore, the studies also gave isotyping results and classified the MAb as the subclass IgG1. We can construct the structural probe against the rFpr3 from these results, and the probe can be used in the study of other immunophilins.
In the structural study of Fpr3, we found that the crystals of the protein were difficult to obtain. On the other hand, we can use NMR to analyze the structure of the protein, while the molecular mass of Fpr3 (70 kDa) is too large to get the three-dimensional structural data. Mosbacher(13) also found that it was difficult to get the crystals of the FK domain in the study of a wheat FKBP73, which is homologous to Fpr3 and hypothesized that the domains contained poorly ordered regions. At the same time, other studies have shown that combining protein with substrate, nucleotides, or other small molecules are useful in stabilizing the protein conformations and obtaining its crystal, without changing the native structure of the protein itself.(14) Therefore, the MAbs in this study can be used as a molecular chaperone to co-crystallization and obtaining the crystal of rFpr3 as well. In comparison with other ligands, this MAb is inexpensive, stable, and specific. Relevant works are now underway.
Footnotes
Acknowledgment
This research was supported by NSFC (no. 20702017), Natural Science Foundation of Hubei Province of China (no. 2010CDB01206).
Author Disclosure Statement
The authors have no financial interests to disclose.