
Abstract
The use of lignocellulosic biomass is critical for the economic production of transportation fuels and chemicals in renewable bioprocesses. While biomass is an abundant resource, necessary pretreatment to yield fermentable monosaccharides produces toxic compounds that dramatically affect fermentation performance. Weak acids such as acetic acid play an important role in the toxicity of lignocellulosic hydrolysate to Saccharomyces cerevisiae, a commonly used industrial organism. In order to explore the ramifications of weak acid inhibition on cellular metabolism, we adapted a genome-scale metabolic model of S. cerevisiae to describe toxicity of acetic acid by a decoupling mechanism. We evaluated the performance of the model in predicting growth rates and ethanol production characteristics under aerobic and anaerobic cultivations. We found that the model was able to capture the decreased growth during aerobic cultivations in the presence of acetic acid, but was unable to capture the increase in ethanol yield observed. The model was able to predict anaerobic growth rates and ethanol yields; however, at conditions of higher toxicity levels, discrepancies arose. We expect that a model such as this may find application in the optimization of lignocellulose-based bioprocesses in which there exists a critical economic trade-off between neutralization costs and product yields.
Introduction
In recent years, renewable means of producing transportation fuel and chemicals have been heavily investigated in order to reduce reliance on natural oil resources. It has been previously noted that as a result of increasing global rate of oil consumption and decreasing rate at which new fossil fuel resources are discovered, alternative sources of transportation fuels and chemicals are highly sought after. 1 Bioprocesses offer an attractive alternative to petroleum-based processes by potentially converting renewable biomass feedstock to drop-in replacements for petroleum-derived chemicals. In particular, bioprocesses that use lignocellulosic biomass, an abundant and renewable resource, as feedstock are among the current areas of focus in the field of biotechnology.
Lignocellulose is composed of cellulose, hemicellulose, and lignin. Cellulose and hemicellulose are long-chain polymers of saccharides that must undergo enzymatic treatment to release fermentable monomers. This process is complicated by the presence of lignin—a complex aromatic polymer—that makes it necessary for the biomass to undergo a pretreatment such as weak acid hydrolysis prior to enzymatic treatment. 2 A significant limitation of pretreatment processes is that inhibitory compounds are produced that significantly impact fermentation characteristics. Inhibitory compounds can be classified as phenolics, furan derivatives, or weak acids, and mechanisms of these inhibitor classes have been reviewed extensively. 2 –4
Weak acid toxicity is a particularly interesting physical phenomenon arising from the interaction of internal and external pH and cellular response mechanisms. The toxicity of weak acids has largely been attributed to the acidification of the cytosol by decoupling and anion accumulation. 3,5 The decoupling mechanism of weak acid toxicity is a result of the ability of non-polar undissociated weak acid species to diffuse across the cellular membrane to the near-neutral pH cytosol. Upon entering the cytosol, the acid dissociates, yielding a proton and the conjugate base. To maintain neutral cytosolic pH, the cell uses the reverse action of adenosine triphosphate (ATP) synthase, exporting protons at the expense of ATP. In addition, the anion accumulation theory proposed by Russell states that the anionic form of the acid species may accumulate in the cytosol and drive the diffusion of undissociated acid across the cell membrane towards equilibrium. 3,5
A significant weak acid inhibitor is acetic acid, which is formed largely from the degradation of hemicellulose and is present in hydrolysates in concentrations up to 10 g/L, depending on the biomass source. 6 While many studies have focused on strain engineering for tolerance to furan derivatives and phenolics, there are significantly fewer that focus on improving tolerance to weak acids. 6 –8 Notably, Hasunuma et al. demonstrated that a recombinant xylose-utilizing strain of Saccharomyces cerevisiae over-expressing the TAL1 gene had improved ethanol yield in the presence of acetic acid. 6 Recently, an acetic-acid-tolerant strain of S. cerevisiae was generated using a genome shuffling method. 9 However, hypothesis discovery remains a bottleneck in rational strain engineering for tolerance to weak acids.
Mathematical modeling is a powerful tool for examining the metabolism of microorganisms. Previous studies have modeled the impact of weak acids on microorganisms using kinetic and thermodynamic approaches; however, such studies have largely been for the purpose of food preservation. 10 –12 In the past decade, constraint-based metabolic models have been demonstrated to be powerful tools for describing cellular phenotype while not requiring enzyme kinetic parameters. 13 A particularly useful tool in constraint-based modeling is Flux Balance Analysis (FBA), wherein realistic rates of nutrient uptake based on the growth environment are defined; a pseudo-steady state assumption is applied; and metabolic fluxes are determined by solving the system as a linear programming problem. In the past, these models have successfully been used to predict experimental phenotypes. 14,15
In this study, we attempt to describe the toxic effects of acetic acid on the metabolism of S. cerevisiae using the iMM904 genome-scale metabolic model. 16 A workflow of the study is presented in Fig. 1. We demonstrate that the model's predictive power may be enhanced with the inclusion of additional constraints describing the uptake of acetic acid by simple diffusion and export of protons via ATP synthase. We anticipate that this work could be built upon to further improve the predictive quality of metabolic models and improve their use in industrial settings. To the best of our knowledge, this is the first instance of incorporating concentrations of inhibitory compounds present in lignocellulosic hydrolysates into an FBA framework. The model could find further use in the optimization of lignocellulosic-based bioprocesses, in which there exists a trade-off between fermentation characteristics and cost of neutralization.
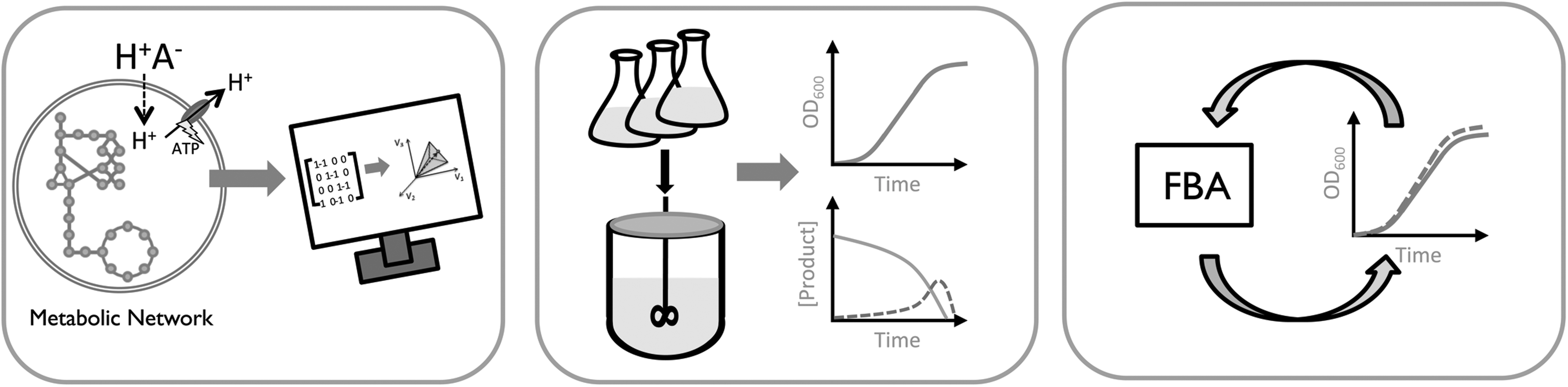
Workflow of the current study. The iMM904 metabolic model of S. cerevisiae was amended with reactions liquid chromatography to describe weak acid toxicity by the decoupling mechanism
Materials and Methods
Metabolic Modeling
The genome-scale compartmentalized iMM904 S. cerevisiae metabolic model was adapted to describe toxicity of acetic acid via the uncoupling mechanism. Figure 2 shows a schematic of reactions added to the model. The concentration of undissociated acetic acid in the extracellular environment was modeled as a function of pH and concentration of acid species using the Henderson-Hasselbalch equation:
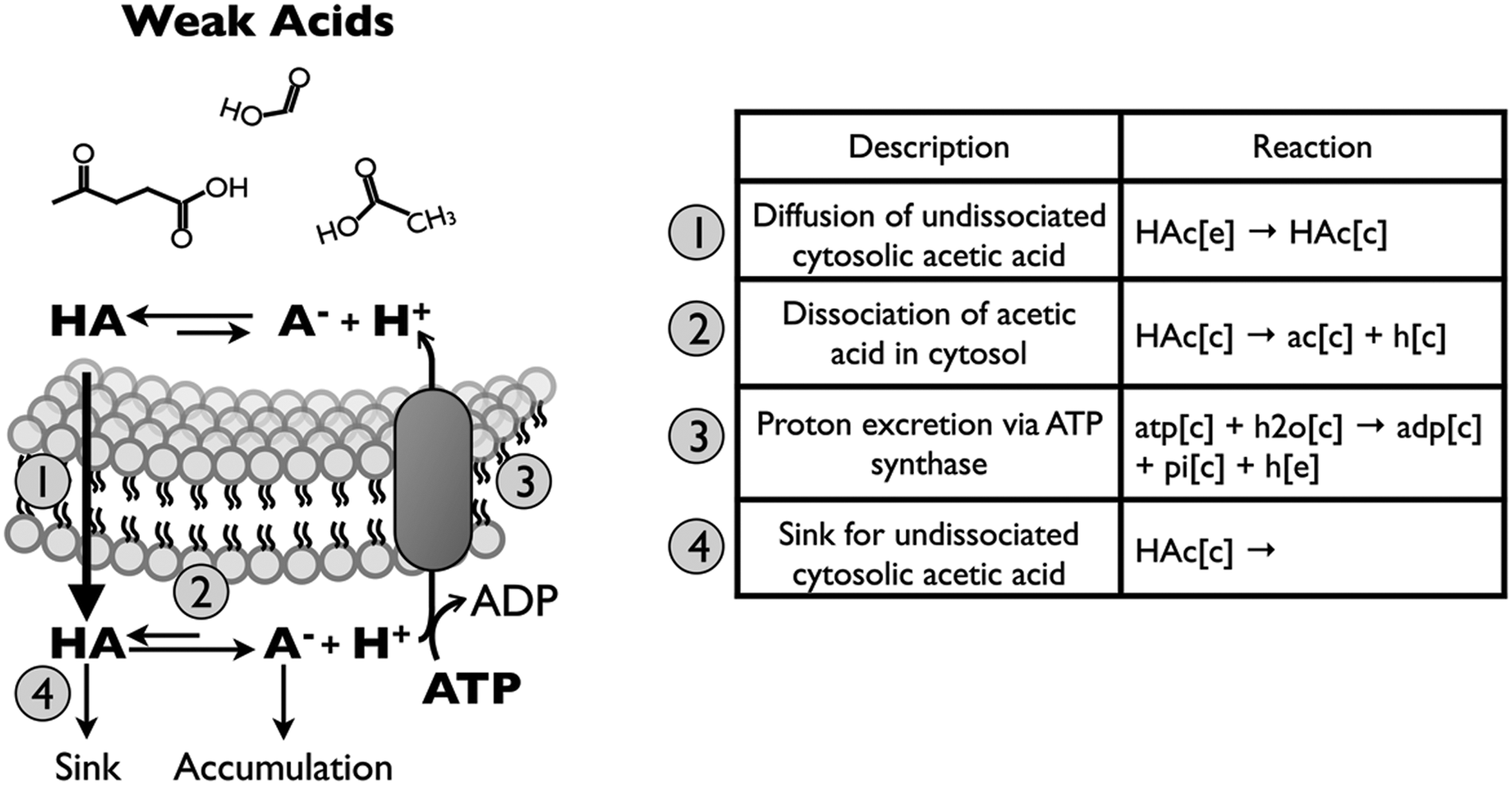
Reactions describing weak acid toxicity by decoupling mechanism. Non-polar undissociated weak acids may diffuse across the plasma membrane by simple diffusion and dissociate due to near-neutral pH of the cytosol. To maintain cytosolic pH neutrality, protons are excreted at the expense of ATP by the reverse action of ATP synthase.
where
Simulations of aerobic cultivations were carried out using a glucose uptake rate of 15.9 mmol/gDW/hr based on measured experimental rates and an oxygen uptake rate of 8.8 mmol/gDW/hr as measured by van Hoek et al. 18 For anaerobic simulations, oxygen uptake rate was set to 0 mmol/gDW/hr. FBA was conducted using the COBRA toolbox. 19,20 Cultivations that were conducted with no exogenous acetic acid added to the growth media were used to fit the first data point of the model by altering the ATP maintenance requirement and glucose uptake rate.
Strains, Media, and Culture Conditions
The prototrophic strain S. cerevisiae CEN.PK 122 was used in all experiments. Cells were maintained on Yeast Extract Peptone Dextrose (YPD) medium. Preculture conditions were as follows: cells from frozen stock were cultured overnight in YPD at 30°C and washed three times with sterile, filtered water before being transferred to baffled flasks containing mineral medium (as described in a previous study) supplemented with 20 g/L glucose and 3 g/L glacial acetic acid. 21 The cells were then cultured overnight at 30°C and 250 rpm. This procedure was used for inoculating both flask- and batch-cultivation experiments. In flask experiments, exogenous acetic acid concentration was varied between 0–5 g/L and pH was not controlled. Aerobic batch-cultivation experiments were conducted in Infors (Basel, Switzerland) 5-L Mini Reactors (3-L working volume), and pH was controlled with the addition of 4 M KOH.
Analysis
Metabolites were measured using high-performance liquid chromatography (HPLC; Aminex HPX-87H column, Bio-Rad Laboratories, Hercules, CA); 5 mM H2SO4, 50°C, 0.4 mL/min. Growth was monitored through optical density measurements at 600nm, and cell concentration was calculated using a correlation of 1 OD600=0.4 gDW/L.
Results and Discussion
Model Characteristics
The iMM904 metabolic model of S. cerevisiae was amended to include reactions that describe the uncoupling mechanism of acetic acid toxicity. Using an FBA approach, we found that model predictions more closely matched experimental data of S. cerevisiae grown in the presence of acetic acid, as compared to the original model, which does not include reactions that account for weak acid toxicity. The fraction of acetic acid species existing in an undissociated form is a function of external pH and the pKa of the compound, related by the Henderson-Hasselbalch equation (Fig. 3A). In optimal growth conditions—with an external pH of 5.5—approximately 15% of external acetic acid exists in its undissociated form and is therefore able to diffuse across the cellular membrane (Fig. 3A). We used the literature values reported in Casal et al. that describe the rate of simple diffusion of undissociated acetic acid across the membrane of S. cerevisiae as a function of concentration and external pH. 17
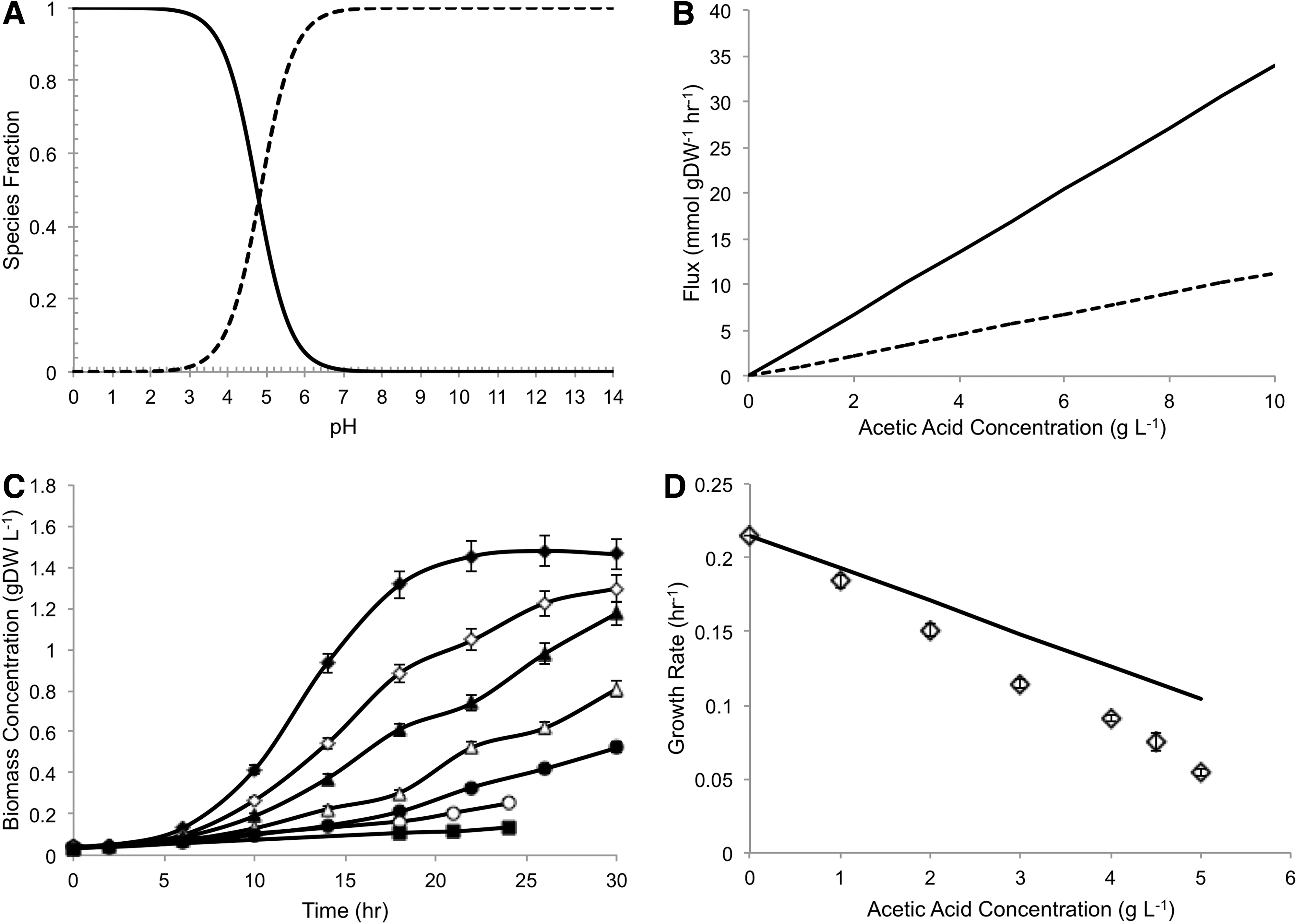
Model characteristics and prediction of aerobic cultivation in baffled flasks. Henderson-Hasselbalch equation relating dissociated acetic acid (dashed black line) and undissociated acetic acid (solid black line). Calculated using pKa of acetic acid=4.75, pH=5.5
Upon entering the cytosol, which is nearly neutral in pH, less than 1% of acetic acid species exist in the undissociated form (Fig. 3A). Thus, from the Henderson-Hasselbalch equation, it is expected that the majority of acetic acid entering the cytosol will contribute to the acidification of the cytosol and ATP depletion. Under the assumption of steady state, the rate of acetic acid dissociation in the cytosol must be equal to the diffusion rate of acetic acid from the extracellular environment to the cytosol. At steady state, the rate of ATP hydrolysis due to weak acid toxicity is therefore equivalent to one-third the rate of cytosolic acetic acid dissociation, reflective of one molecule of ATP transporting three protons out of the cytosol (Fig. 3B).
As a result of diffusion of undissociated acetic acid into the cytosol, we observed changes in the flux distribution surrounding cytosolic acetate node (Fig. 4). Specifically, flux through the cytosolic NADP+-dependent aldehyde dehydrogenase reaction (ALDD2y), converting acetaldehyde and NADP+ to acetate and NADPH, was eliminated in the presence of small concentrations of exogenous acetic acid. We reasoned that this is likely due to cytosolic acetate being supplied by the diffusion of acetic acid into the cytosol and its subsequent dissociation. This insight is interesting in that the refined model predicts that exogenous acetic acid is to some extent directed towards production of biomass, likely through the synthesis of acetyl-CoA, as was previously suggested by Taherzadeh et al. 22 The balance of acetate was predicted to be exported from the cytosol through the action of an acetate transporter (ACtr).
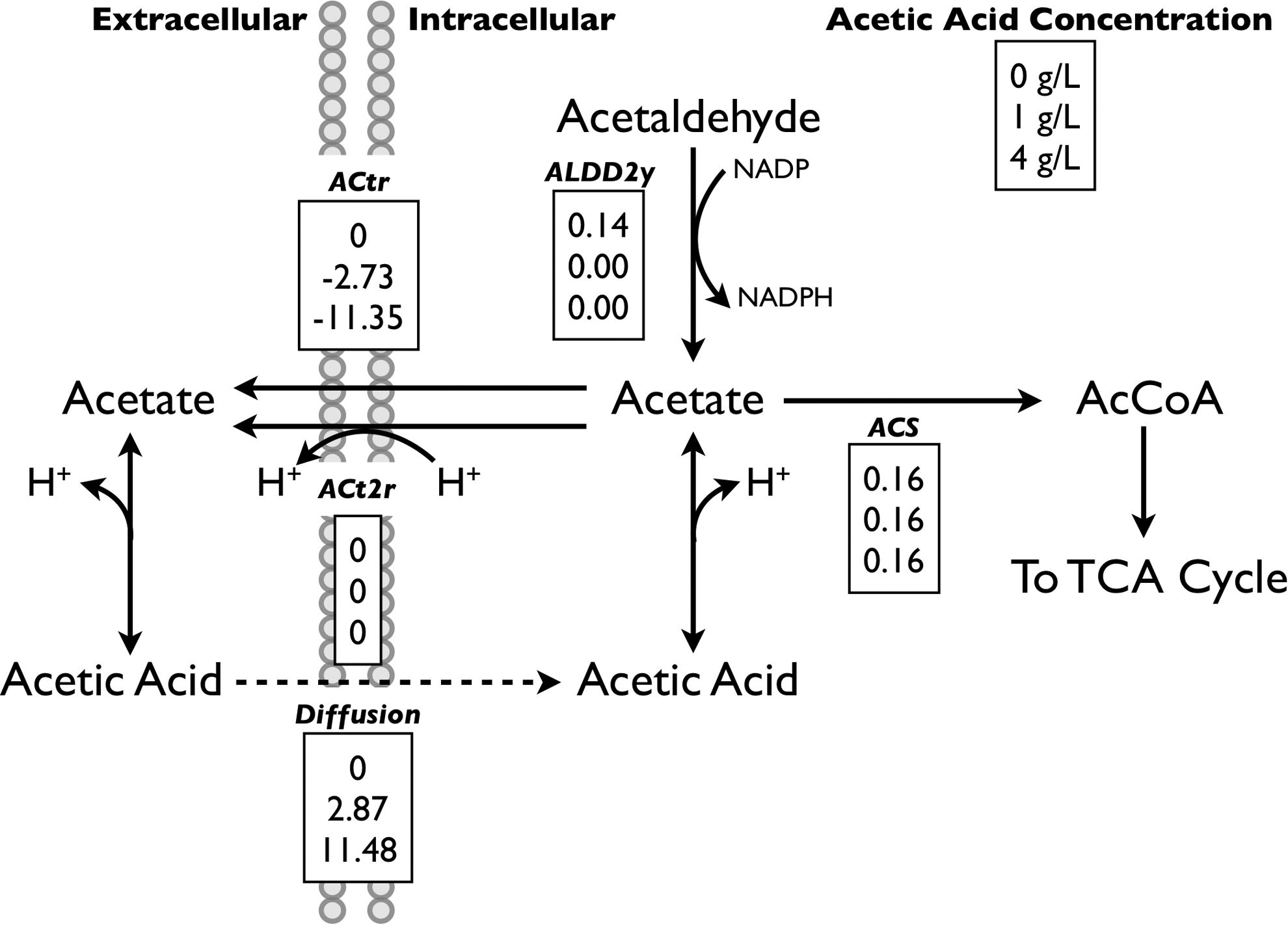
Predicted flux distribution around the cytosolic acetate node in the presence of exogenous acetic acid. Predicted flux distribution around the cytosolic acetate node in the presence of acetic acid. Values reported in mmol/gDW/hr.
Prediction of Growth Under Aerobic Conditions
Initial aerobic cultivations were conducted in baffled shake flasks with mineral media supplemented with 20 g/L glucose and varying concentrations of acetic acid without buffering the change in pH (Fig. 3C). In these cultivations, we found a linear correlation between growth rate decrease and increase in acetic acid concentration (Fig. 3D). Experimental growth rates were predicted using an external pH of 5 to approximate the starting pH of the media used. Predictions were good for low concentrations of acetic acid (up to 1 g/L), after which point the model was unable to capture fully the toxic effects of the compound. We reasoned that the discrepancies between the model and experimental results could be partially attributed to lack of pH control, particularly at higher concentrations of acetic acid. The external pH of growth medium plays a critical role in the toxicity of acetic acid, as lower external pH increases the fraction of total acetate species that remains undissociated and therefore may transverse the cytoplasmic membrane. Under conditions that lack pH control, thereby allowing the pH to decrease as the cultivations continue, it is expected that the toxic effects would be magnified.
To account for variations in external pH, further cultivation studies were conducted in fermenters with pH controlled to 5.5 using 4M KOH. In these studies, we found a nonlinear relationship between the concentration of acetic acid and decrease in growth rate (Fig. 5A). Low concentrations of acetic acid were found to have less impact on growth rate as compared to the higher concentrations, as expected based on the relative concentrations of extracellular. undissociated acetic acid species. Although the predictions of growth rate at low concentrations of acetic acid matched experimental data closely, we found that the model was unable to capture toxicity at higher concentrations. We qualitatively assessed the efficacy of the model by both increasing and decreasing the acetic acid diffusion rate by 50%, corresponding to an increase and decrease in toxic effects, respectively. We found that despite a 1.5-fold increase in acetic acid diffusion, the model was still unable to capture the reduction in growth rate experimentally determined. This result suggests that significant toxic effects arise from secondary mechanisms of toxicity, such as anion accumulation, osmotic stress, and increased maintenance requirement, and demonstrates that the decoupling mechanism alone is unable to account for the decrease in growth rate observed experimentally. 5
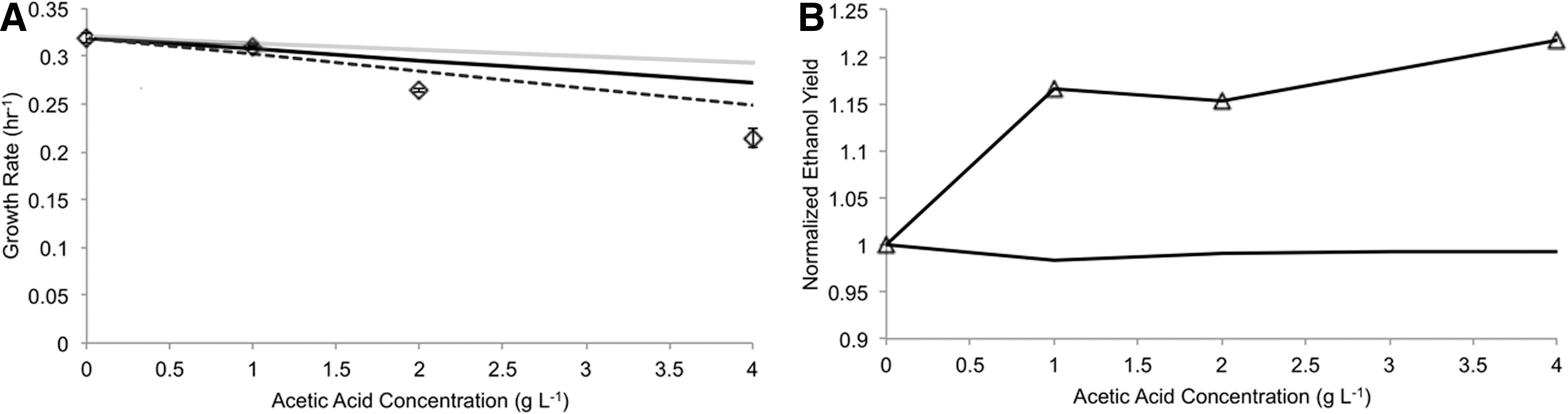
Prediction of aerobic batch cultivation. Experimental (open diamonds) and predicted aerobic growth rates (solid black line) of S. cerevisiae at pH 5.5 in the presence of acetic acid. Error bars indicate standard deviation between two or more experiments. The diffusion rate of acetic acid was increased and decreased by 50% (dashed and solid gray line, respectively)
Moreover, we found that the model was unable to capture increases in ethanol yield due to the presence of acetic acid (Fig. 5B). A previous study by Taherzadeh et al., found that the presence of acetic acid increased the yield of ethanol in fermentations. 22 Model predictions indicated a minor increase in ethanol yield with increasing concentration of external acetic acid, but did not match experimental results. A small portion of this discrepancy may be attributed to the assimilation of acetate by conversion to acetyl-CoA by acetyl coenzyme-A synthetase, although it has previously been suggested that this consumption is small compared to the rate of acetic acid uptake. 22 Moreover, varying the oxygen uptake did not improve the prediction of ethanol yield in the anaerobic case (data not shown). It is likely that the addition of constraints that accurately reflect the physiology of S. cerevisiae may be included to improve predictions and will be the subject of future studies.
Prediction of Growth in Anaerobic Conditions
To evaluate the accuracy of the model, we used anaerobic experimental data reported by Taherzadeh et al. 22 In that study, the authors evaluated the impact of acetic acid on the batch fermentation of glucose to ethanol by S. cerevisiae CBS 8066 and reported the fermentation characteristics at pH values of 3.5 and 5.0. 22 Using the same model as the aerobic case, and a glucose uptake rate reflective of the fermentations conducted by Taherzadeh et al., we predicted the growth rates and ethanol yields at pH values 3.5 and 5.0, as reported in the study (Fig. 6). Model predictions were in good agreement with experimental values reported for the growth rates and ethanol yields in the presence of acetic acid in cultivations at pH 5.0 and 3.5 (Fig. 6). However, the refined model was unable to predict the growth rate at high concentration of acetic acid at an external pH of 3.5. Moreover, in the pH 3.5 cultivations, a given increase in acetic acid concentration resulted in a larger increase in ethanol yield than was predicted by the refined model. In addition to secondary mechanisms of weak acid toxicity that may contribute to discrepancies observed between the experimental and model predicted growth rates and ethanol yields, it is possible that additional impacts of extracellular pH on S. cerevisiae also contributed to prediction error. The impacts of the pH of cultivation media are unable to be captured by metabolic models and could have contributed to the reduction in growth rate as well as the increase in ethanol yields reported by Taherzadeh et al. 22 Nonetheless, we found that the refined model was able to predict growth rates and ethanol yields for anaerobic conditions.
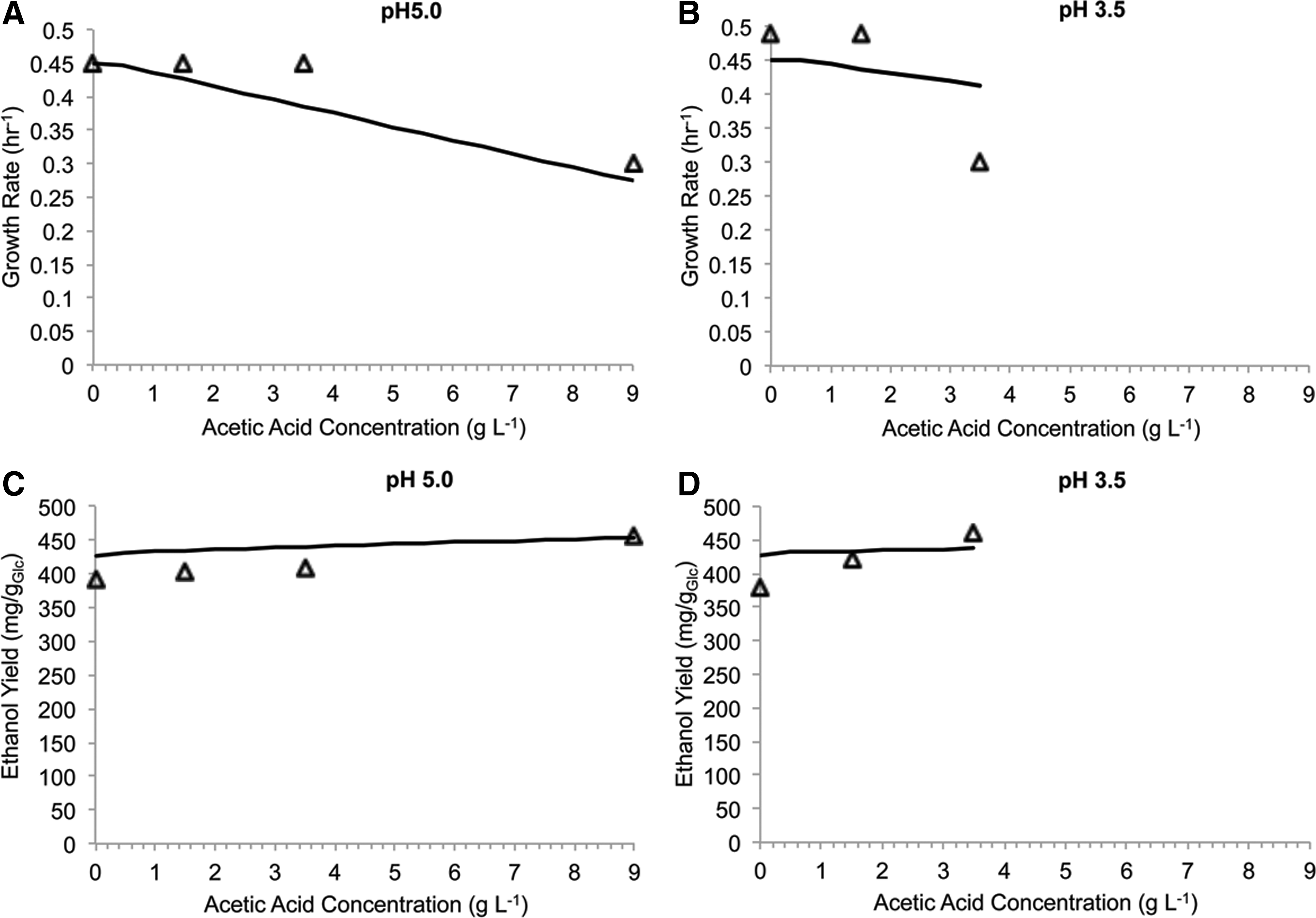
Prediction of anaerobic cultivation. Prediction of batch fermentation of glucose to ethanol by S. cerevisiae CBS 8066. Experimental data (open triangles) from Taherzadeh et al.22 Growth rate (solid black line) was predicted for pH 5.0
Conclusions
We used constraint-based metabolic models to predict the toxicity of acetic acid in S. cerevisiae. We amended the iMM904 model of S. cerevisiae to include reactions that describe the uncoupling mechanism of weak acid toxicity and showed that the model could describe the trends of growth rate and ethanol yield observed in aerobic and anaerobic cultivation experiments using FBA.
The refined model was able to capture, to a certain extent, the toxicity of acetic acid under aerobic cultivation; however, we found that at higher concentrations of the inhibitor, the model could not capture the full toxic effect. We believe that the discrepancy between experimental and model results is due to the secondary inhibitory mechanisms of weak acids that the model did not account for, such as anion accumulation or an increase in maintenance requirement. Additionally, inconsistencies observed between experimental and predicted results could be a consequence of differences between the strains cultivated in experiments and the S. cerevisiae S288C strain upon which the metabolic model is based. We found that the refined model was unable to predict the trends in ethanol production in the presence of acetic acid under aerobic conditions, suggesting that the addition of further constraints representing the physiology of S. cerevisiae may be required.
The refined metabolic model was capable of predicting anaerobic growth rates and ethanol yields for a pH of 5.0; however, at higher levels of toxic effect—ie, lower external pH and higher concentration of acetic acid—discrepancies arose between the model and experimental data. This result shows conclusively that dissociation cannot fully account for the growth defect. We believe this model could find application in the optimization of lignocellulosic bioprocesses, in which there exists a delicate trade-off between cost of neutralization and process productivity. It is possible that constraint-based modeling alone cannot capture other mechanisms of toxicity such as phenolics and furan derivatives, as they have inhibitory impacts on enzymatic activity rather than affecting steady state yields. In these situations, it is likely that coupling a constraint-based model with a kinetic model could improve model predictions.
Footnotes
Acknowledgments
The prototrophic strain S. cerevisiae CEN.PK 122 used in all experiments was a gift from Dr. Vince Martin at Concordia University (Montreal, Canada).
Author Disclosure Statement
The authors declare no competing financial interests exist.