
Abstract
Microalgae biomass is a promising source of plant protein that has potential applications in the food, cosmetics and pharmaceutical industries. However, the presence of a strong cell wall in microalgae cells creates difficulties in obtaining these proteins. This study aims to establish an efficient protocol for disruption of Chlorella sorokiniana microalgae cells to improve the extraction of intracellular water-soluble proteins. C. sorokiniana microalgae cells were disintegrated using ultrasound, microwave radiation, and enzymatic lysis with lysozyme. Ultrasound was the most effective of the cell disruption methods applied separately, increasing the yield of water-soluble proteins by up to 11.4% compared to the control sample. When ultrasound and enzyme were used consecutively, a synergetic effect was observed, as the yield of intracellular water-soluble proteins significantly exceeded (up to 35%) the result achieved by using a single disruption method. The combined effect of physical and biochemical cell disruption methods on the extraction of intracellular water-soluble proteins was also investigated. It was established that several methods applied sequentially to disrupt cells produced a combined, synergetic effect. A mechanism of sequential action of two cell disruption methods, explaining the synergy and increased yield of water-soluble intracellular proteins, is proposed.
Introduction
Microalgae cells are a promising source of dietary plant protein as they have greater photosynthetic efficiency than higher plants and a more flexible metabolism. 1 Protein hydrolysates of microalgae have good emulsifying properties and high antioxidant activity, which leads to their potential in nutraceutical and cosmetic industries. 2 In addition, some polypeptides extracted from Chlorella cells have anti-tumor activity and therefore have potential applications in the food and pharmaceutical industries. 3
In the study, 4 it was determined that the molecular weight of microalgae protein isolate was mainly distributed in the areas of 0.18–0.50 kDa, 0.50–1.50 kDa, and 1.50–5.00 kDa, and the contents of essential amino acids in such isolates even exceed the values recommended by FAO/WHO standards. 5 The high value of the essential amino acid index (1.49) of microalgae isolates is an important indicator that suggests that the resulting protein hydrolysate is suitable as a nutritional source for humans. 6 Depending on the microalgae strain, the cellular protein can be equal or superior in protein amino acid composition to traditional plant sources. 7 Microalgae cell proteins can account for up to 42–58% of the dry cell biomass, of which about 20% are part of the cell wall, 50% are intracellular enzymes, and the remaining 30% are secreted into the extracellular medium. 8
The difficulty in extracting proteins from microalgae cells is that most microalgae species possess rigid cell walls, 9 which prevent protein extraction. Microalgae cell walls can vary greatly in structure, but most commonly contain the following polymers: cellulose, hemicellulose (xyloglucan, mannan, glucuronan, (1 → 3)-β-glucan), a chitin-like peptidoglycan and ulvan. 9 This fact poses a serious problem when using microalgae biomass as a source of dietary nutrients, as it is not digested by humans and non-ruminant animals.
To break down the cell wall and make the intracellular protein available to digestive enzymes, efficient disintegration methods are needed. It is important to select cell disruption methods that will produce a high-quality target product that can be used as a food nutrient. Finding ways to improve the energy efficiency of the process of extracting valuable components from cells has received much attention in research, as it accounts for up to 40% to 60% of the total cost of the final product. 10 Cell disruption methods can be divided into three groups: mechanical (physical), chemical and biochemical, and depending on the type of producer used, can be used separately or in combination.
When selecting disintegration methods, the structure of the cell membranes, their mechanical and biochemical characteristics (resistance to load, deformation, destruction; shaping, etc.) must be taken into account. 11
With physical disintegration methods, cells are exposed to a physical field (hydrodynamic, acoustic, etc.), which leads to the disruption of the cell wall. In ball mills, the destruction of the cell membrane occurs through direct mechanical contact between the grinding media and the cells. A disadvantage of this method is the low productivity and high energy consumption rate of the equipment. When exposed to microwave radiation, there is an interaction between the polar molecules inside the cell that creates a significant intermolecular friction as the molecules rotate about their axis, causing intracellular water boiling and rupture of the cell membrane. 12
In the extrusion method, which uses injector nozzles and homogenizers, a cell suspension is pressed through narrow holes under high pressure into low-pressure chambers, resulting in shear stresses that lead to cell rupture. When the cells are dried, the resulting dehydration shock causes tears and cracks in the cell, and the cells themselves die. Exposure to ultrasound leads to destruction of the cell wall of the membrane due to the various physical and physico-chemical phenomena, which have a cavitation nature. 13,14
Chemical methods of treating the cell involve exposure to substances of a different nature. In osmotic shock, the cell membrane is ruptured by water pressure, which penetrates into the cell with a sharp change in osmotic pressure due to an increase (decrease) in the concentration of salt or sugar in the environment. Osmotic shock is most effective when disintegrating cells with weak and permeable cell membrane. The application of chemicals involves alkalis, acids, salts, chelating agents, and organic solvents. These reagents, affecting the cell membrane, cause denaturation and coagulation of proteins, oxidation of cell wall components, and destruction of membrane structure. A disadvantage of the method is the impact of the reagents on other cellular structures, which may result in the destruction of the target product. 11
Enzymatic lysis involves treating a cell suspension with hydrolytic enzymes or enzyme complexes that break the bonds between cell wall substances. Different degradation mechanisms are implemented when antibiotics are applied. 11 For example, exposure of cell wall enzymes to antibiotics leads to their binding, causing the cell to lose the ability to form new cell wall components and resulting in the death of the microorganism. Also, when the active center of the antibiotic interacts with the lipids of the cytoplasmic membrane, the membrane is damaged; pores are formed through which cellular components leak into the culture liquid.
Selection of an effective method of microalgae cell disruption increases the contact surface area of the source material (microalgae cells) and the extractant, which increases the efficiency of the extraction process of the target component. When selecting a method for disintegration of microalgae cells in technologies for production of protein compounds, the purity of the resulting compounds is an important factor, so it is important to choose methods of disruption that do not contaminate the target product.
The aim of this study is to compare the most promising methods for disintegration of microalgae cells: lysozyme enzyme, microwave radiation, and a combination of the two.
Materials and Methods
SUBJECT OF RESEARCH
The subject of the study was the strain Chlorella vulgaris Beijer Institute of Plant Physiology Russian Academy of Sciences (IPPAS) C-1 (Chlorella sorokiniana) from the collection of the Timiryazev Institute of Plant Physiology, Russian Academy of Sciences. 15 The microalgae strain was cultured in 2-L photobioreactors for 8 d on Tamiya nutrient medium, which was introduced on the first and fourth days, under autotrophic conditions at the level of photosynthetically active radiation of 150 μmol photons/(m2·s). The amount of inoculum introduced was 10% of the total suspension, the cultivation temperature was 30 ± 3°C, and pH varied from 6.2 to 8.0. Aeration of the suspension (180 L/(L·h)) was performed with an air-gas mixture containing 0.03% carbon dioxide. 12 The number of cells in the suspension was 40 million cells/mL.
BIOMASS CONCENTRATION
Separation of the centrifuge effluent from the microalgae biomass was performed using a centrifuge (Sigma® 2-16 RK/2-16P; SIGMA Laborzentrifugen, Osterode, Germany) at 4,000 rpm for 10 min.
CELL DISRUPTION
Experiment 1
The resulting biomass with a moisture content of 98–99% was divided into 25-mL samples. Microalgae cells were disrupted using the following methods: Cell disruption using lysozyme enzyme taken in an amount of с = 5–15 mg/g biomass at 37°C for τf = 4–16 h;
8
Cell disruption using a microwave irradiator (LG® MB-40R42DS; LG Electronics, Tianjin, China) (ultra-high frequency radiation) of PMW = 280–560 W for τMW = 10–30 s (1, 2 or 3 treatment cycles (nMW); the break between treatment cycles was 300 s, during which the samples were cooled to 40–45°C);
16
Ultrasound treatment of cells (Scientz® IID disintegrator, Scientz, Zhengzhou, China, at 25 kHz, PUS = 50–150 W for τUS = 60–300 s. Regimes 1–4 (nUS): 10 or 50 treatment cycles at 5 s (ultrasound treatment)/1 s (break); regimes 5–8 (nUS): 30 or 150 treatment cycles at 1 s (ultrasound treatment)/1 s (break);
16
regime 9: 100 W ultrasound treatment for 180 s (45 treatment cycles at 3 s (ultrasound treatment)/1 s (break).
The negative control was the microalgae biomass used for the extraction of water-soluble intracellular proteins, without prior disintegration.
Experiment 2
The resulting biomass with a moisture content of 98–99% was divided into 25-mL samples. Microalgae cells were disrupted using the following methods (Table 1):
Parameters of Complex Cell Disruption
When treated with 150 W ultrasound, the exposure time was 5 min (treatment regime 5 s (ultrasound treatment)/1 s (break).
Exposure to microwave radiation for 30 s, radiation power 560 W, 3 treatment cycles.
Enzyme treatment time at concentration of 15 mg/g was 4 h, at 37°C.
DETERMINATION OF THE DRY MATTER OF MICROALGAE CELLS IN A CONCENTRATED SUSPENSION
A preweighed tube with concentrated microalgae cell biomass was placed in a desiccator at 80°C and dried to a constant mass (Δ = 0.01 g), then weighed. The dry biomass was determined according to the formula:
where M is the dry biomass, g/mL; a is the mass of tube with concentrated microalgae cell biomass, g; b is the mass of tube (filter) without sediment, g; V is the volume of culture liquid, mL.
EXTRACTION OF WATER-SOLUBLE INTRACELLULAR PROTEINS FROM MICROALGAE BIOMASS
Extraction of proteins from microalgae biomass was carried out for 20 h at 4°C using 25 mL of phosphate buffer (Na2HPO4, Lenreactive, Saint Petersburg, Russia), KH2PO4, Lenreactive) (pH 7.2–7.4) as solvent. 17
EXTRACT SEPARATION FROM BIOMASS
After extraction, the cell biomass was separated from the extract using a centrifuge (IKA-Werke; IKA® mini G centrifuge, Staufen im Breisgau, Germany) for 10 min at 6,000 rpm).
DETERMINATION OF WATER-SOLUBLE PROTEIN CONCENTRATION IN THE EXTRACT
Protein content in the extract was determined using spectrophotometric method (Spectrophotometer PE-5400 UV; EKROSCHEM® LLC, Saint Petersburg, Russia) 18 and using Maxlife® Personal Gene Analyzer H 100 fluorometer for the quantitation of DNA, RNA, and protein. The average value of the concentrations obtained using the two techniques was used for the calculations.
CALCULATION OF THE ENERGY COSTS FOR CELL DISRUPTION IN A 25-ML SUSPENSION
Total energy consumption for cell disruption Zf (J) in 25 mL suspension when treated with lysozyme involves the following components:
where Z1 is the energy consumption for heating a microalgae suspension from t1 = 24°C (temperature in laboratory) to t2 = 37°C (optimal temperature for lysozyme activation determined by formula Z1 = cs·m·(t2–t1), where cs is the heat capacity of microalgae suspension, J/(g·K), m is the mass of microalgae suspension; Z2 – the cost of maintaining the temperature at 37°C for the whole treatment time (taken as 40 J/h), J.
Energy costs ZMW (J) for treatment of microalgae biomass (25 mL) with microwave radiation were determined by the formula:
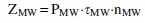
where PMW is the power of microwave radiation, W; τMW is exposure time, s; nMW is the number of treatment cycles.
Energy consumption ZUS (J) for treatment of microalgae biomass (25 mL) by ultrasound was determined by the formula:
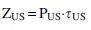
where PUS is the power of ultrasound, W; τUS is the time of exposure to ultrasound, s.
DETERMINATION OF CELL CONCENTRATION IN SUSPENSION AND AVERAGE SURFACE AREA OF CELLS
The diameter of 50 randomly selected cells was determined using an ocular micrometer. The formula was used to calculate the average cell surface area:
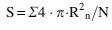
where R is a cell radius, μm.
Results and Discussion
Microalgae biomass used in the experiments contained cells at different stages of development, which can be conventionally classified into young cells (cell diameter 1–2.5 μm) and mature cells (cell diameter over 2.5 μm). The main difference is the presence of a formed cell wall layer of chitin-like glycan in mature cells and different intracellular osmotic pressure. 19
As a result of the enzyme action, the chitin-like glycan layer of the cell wall is disrupted, which reduces the strength of the cell wall. 20 At the same time, the enzyme molecules will be incorporated into the phospholipid membrane, resulting in the formation of pores in the membrane, through which water from the external environment will enter the cell. Cells will increase in size and some of will be disintegrated, while the rest will increase the intensity of mass exchange between the internal volume and the external medium. At low dosages of the enzyme, more intensive disruption of mature cells is observed, resulting in a decrease in the average cell surface area (Table 2, regimes 2 and 4).
Experimental Results on Cell Disruption
By increasing the initial concentration of the enzyme by the 4th hour of treatment (regime 3), an increment in the average cell surface area by 1.2 times (up to 47.8 μm2) is observed compared to the control sample. This is apparently due to the formation of pores in the phospholipid membrane of young cells, which leads to a change in osmotic pressure and water entry into the cells. These cells are then disrupted by the 16th hour of treatment (regime 1), as evidenced by a 53% decrease in average cell surface area compared to the control sample.
When the enzyme dosage is increased, the protein yield increases 1.6-fold; at the same time, an increase in the enzyme treatment time from 4 to 16 h decreases the protein yield by 17%, which seems to be related to degradation of a part of protein molecules during a long stay in the extracellular space. Enzyme treatment is a process limited by slow diffusion, which can be intensified by pre-treating the cells with another disintegration method.
The maximum protein yield during disintegration of microalgae cells by microwave irradiation (regime 1) was higher than after enzymatic treatment and amounted to 5.5% (Table 2). Microwave radiation causes disruption of microalgae cells due to the appearance of 'local overheating' in the intracellular space, arising from the rapid rotation of polar molecules in the generated alternating electromagnetic field; the intracellular water boils, resulting in excess intracellular pressure and subsequent cell disintegration. 12 In this case, most of the cells will be completely disrupted, another part will receive local damage to the cell wall and the phospholipid membrane, through which water will enter the cell.
A batch input of energy (regimes 1–4) into the suspension is more efficient in terms of protein yield compared to a single energy input (regimes 5–8). This is probably due to the fact that each treatment stage allows new cell aggregates of polar molecules, which were formed at the previous treatment stage as a result of destruction of large cellular formations—cell wall components, organelles, multiprotein complexes—to be involved in the disintegration process.
Exposure of cells to ultrasound results in the detachment of macromolecules and molecular complexes from the outer surface of the cytoplasmic membrane and cell wall, which is caused by cavitation effects occurring at the 'cell-external medium' interface. This leads to either the complete destruction of the cell or the formation of pores in the phospholipid membrane, allowing water to enter the intracellular space, which in turn causes an increase in cell surface area.
Exposure of the C. sorokiniana microalgae cells to ultrasound (frequency 25 kHz, power 50–150 , 1–5 min) resulted in the highest yield of water-soluble proteins among the three experiments—1.2–10.4 times higher, depending on the treatment regime (Table 2). In cells exposed to ultrasound, macromolecules and molecular complexes are detached from the outer surface of the cytoplasmic membrane and cell wall, which is the result of cavitation effects occurring at the cell-external medium interface. This leads to either the complete destruction of the cell or the formation of pores in the phospholipid membrane, allowing water to enter the intracellular space, which in turn causes an increase in cell surface area.
The results of the experimental data analysis allow for the assumption that the study of consecutive influence of the considered disruption methods on intensity and completeness of protein extraction is a effective approach to ensure increased yield of target product.
Analysis of the results of Experiment 2 (Table 3) showed that consecutive use of different disruption methods significantly increases the yield of intracellular water-soluble proteins (up to 35%) in comparison with the negative control sample.
Results of Experiments on Complex Cell Disruption
Pre-exposure to physical disintegration methods significantly increases the efficiency of the enzyme (regimes 1–2). Introducing the enzyme in the first step followed, without delay, by treatment with ultrasound or microwave radiation (regimes 3 and 4) led to a lower protein yield. This seems to be due to the fact that ultrasound and microwave radiation destroy both the cell walls of microalgae and the native structure of some of the lysozyme molecules, which reduces the efficiency of its effect on cells.
The combination of ultrasound and enzyme is more effective (regime 1) than that of microwave radiation and enzyme (regime 2). The reason is probably the occurrence of local microscopic overheating during exposure to microwave radiation, leading to suboptimal conditions for the enzyme (T = 37°C) and higher efficiency of ultrasound on microalgae cells compared with that of microwave radiation. This is also confirmed by the results of Experiment 1. Pre-treatment of cells with ultrasound significantly increased the efficiency, and apparently shortened the enzymatic treatment time, as new enzyme targets (partially disrupted cell wall microfibrils, openings in the cell wall and phospholipid membrane) appeared after the exposure to ultrasound.
Conclusion
When using only one method of microalgae cell disruption, ultrasound was most effective and increased the yield of water-soluble proteins by 10.4 times (up to 11.4%) compared to the control sample (1.1%). When ultrasound and enzyme are used consecutively, a synergetic effect is observed, as the yield of intracellular water-soluble proteins significantly exceeds (up to 35%) the results achieved by using a single disruption method. The synergetic effect of several cell disruption methods used sequentially to disrupt cells for extraction of intracellular water-soluble protein has been established. A mechanism of the complex effect of various disintegration methods combinations on the process of cell disruption in the microalgae Chlorella sorokiniana has been proposed.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Research was funded by Ministry of Education and Science of the Russian Federation MK-4348.2022.4.