
Abstract
Abstract
Background:
Pulmonary surfactant spreads rapidly over the airway epithelium, a property that could be harnessed to transport drugs into the lungs. For efficient drug delivery, an interaction between pulmonary surfactant and the drug to be administered is likely needed. On the other hand, the interaction should not compromise the activity of surfactant or the drug once delivered in vivo. The antibiotics gentamicin (an aminoglycoside) and polymyxin E represent drugs that could benefit from being delivered directly to the lung, thereby increasing local concentrations and reducing systemic side effects. Our aim was to study how the animal-derived surfactant poractant alfa (Curosurf®) affects the activities of polymyxin E and gentamicin against Pseudomonas aeruginosa.
Methods:
In vitro antimicrobial assays and a neonatal near-term rabbit model were used to evaluate the combinations of antibiotics and surfactant against Pseudomonas aeruginosa.
Results:
The bactericidal activity of polymyxin E, but not of gentamicin, against P. aeruginosa was partly reduced in vitro in the presence of poractant alfa. In contrast, in the rabbit model of P. aeruginosa pneumonia, polymyxin E administrated together with surfactant was superior in lowering the bacterial load in the lungs compared to polymyxin E alone, without affecting plethysmographically recorded lung compliance.
Conclusions:
The results suggest that polymyxin E interacts with poractant alfa, which reduces the antibacterial effect in vitro. However, when polymyxin E mixed with surfactant is used in the in vivo pneumonia model, increased bactericidal effect was observed. This may be due to a more efficient spreading mediated by interactions between polymyxin E and surfactant. These results warrant further studies of surfactant preparations for drug delivery against lung infections.
Introduction
E
Several studies have investigated the effects of pulmonary surfactant combined with antibiotics in vitro(5,10–12) and in animal models.(13–16) The effects of antibiotics in vitro are generally unaffected(11) or slightly reduced(5,10,12) in the presence of pulmonary surfactant, while several studies indicate that the therapeutic effect of surfactant/antibiotic mixtures in animal models is increased.(13–16)
A possible explanation for this discrepancy is that the surfactant may interact with the antibiotics and attenuate the activity in vitro, but at the same time, the interaction results in a more efficient spreading of the antibiotic in vivo. The chemical properties differ between antibiotics and consequently their interaction with surfactant preparations cannot be generalized. Therefore, the activity of different antibiotics in the presence of different surfactants must be experimentally determined.
Pseudomonas aeruginosa is a Gram-negative and opportunistic pathogen that is able to rapidly develop resistance to several types of antibiotics.(17) Ventilator-associated pneumonia in preterm infants is frequently caused by P. aeruginosa(18) and in cystic fibrosis patients, P. aeruginosa often colonizes the respiratory tract and a chronic pneumonia is established as the strain undergoes phenotypic changes, which cause major problems.(19) Although associated with severe systemic side effects, polymyxins are considered the last resort for treatment of infections caused by several multidrug-resistant (MDR) Gram-negative bacteria such as Klebsiella pneumoniae, Acinetobacter baumannii, and P. aeruginosa.(20)
Polymyxin E (colistin) and B are the two polymyxins used clinically. They are both cationic antibiotics that bind to the lipopolysaccharide (LPS) of Gram-negative bacteria, destabilizing and disrupting the outer membrane. Polymyxin E is the active substance, which is administered as an antibiotic either in the form of colistin sulfate or as the inactive prodrug colistin methanesulfonate sodium. Aminoglycosides represent another group of antibiotics that are used to treat lung infections in ventilator-associated pneumonia and cystic fibrosis patients.(21) Their mechanism of action involves interference of the protein synthesis by targeting bacterial ribosomes.(22) As with the polymyxins, aminoglycosides are associated with serious side effects like nephrotoxicity and ototoxicity when administered systemically.(23)
The aim of this study was to investigate the effect of the animal-derived surfactant poractant alfa on the antibacterial activity of polymyxin E and gentamicin in vitro and to determine if exogenous lung surfactant increases the efficacy of intratracheally instilled polymyxin E against a P. aeruginosa lung infection in neonatal near-term rabbits.
Materials and Methods
Bacteria, pulmonary surfactant, and antibiotics
P. aeruginosa (ATCC27853) was obtained from Culture Collection, University of Gothenburg, Sweden. The animal-derived surfactant poractant alfa (Curosurf) was provided by Chiesi Farmaceutici (Parma, Italy). The antibiotics polymyxin E and gentamicin were purchased from Sigma-Aldrich (St. Louis, MO) as colistin sulfate and gentamicin sulfate, respectively.
In vitro studies
P. aeruginosa was cultivated at 37°C in lysogeny broth (LB) for about 2 h until OD600 ≈0.2. The antibiotics polymyxin E sulfate and gentamicin sulfate were dissolved in phosphate-buffered saline (PBS) to 250, 125, 50, and 25 μg/mL. Eight microliter of each of the antibiotic stock solution or PBS only was mixed with 92 μL of the P. aeruginosa culture (OD600 ≈0.2) and with 100 μL of PBS, or poractant alfa. The final antibiotic concentrations were 1–10 μg/mL, which bracket minimum inhibitory concentration (MIC) values of both antibiotics. The mixtures were incubated with gentle shaking for 6 hours at 37°C. Samples were collected, serially diluted in PBS, and plated on blood agar plates after 0, 2, 4, and 6 hours of incubation, for determination of the number of colony-forming units (CFUs).
The logarithm of CFU/mL was plotted against time and the data shown represent the mean ± standard error of the mean of six independent experiments for the mixtures not containing any antibiotic and of three independent experiments for the mixtures containing any of the two antibiotics tested. The statistical analysis was done using two-way analysis of variance with the GraphPad software (Graph Pad Software, Inc., San Diego, CA).
Cultivation of P. aeruginosa for in vivo studies
Bacterial preparations were made by transferring 75 mL of an overnight culture of P. aeruginosa into 100 mL of LB. After incubation at 37°C for 2 h, the bacteria were washed twice with sterile physiological saline (0.9%, w/v) (NaCl) and subsequently diluted to three different concentrations using OD595 measurements. Plating on blood agar plates and CFU counting was used to determine the CFU/mL.
An animal model of P. aeruginosa pneumonia
Pregnant New Zealand White rabbits were anesthetized and near-term newborn rabbits were delivered by caesarean section at a gestational age of 29 days (term 31 days). The animals were anesthetized right after birth with intraperitoneal (ip) injection of 2 mL/kg body weight of 50 mg/mL Ketaminol®/1 mg/mL Domitor®/saline, 4:1:15 (by vol), tracheotomized, and given an ip injection of 25 mg/kg body weight of 3 mg/mL pentobarbital in 9.5% glucose solution. The animals were subsequently placed in plethysmograph boxes at 37°C connected to a respirator through a metal tracheal cannula. The animals were ventilated with 100% oxygen, at a frequency of 40 breaths per minute, at an inspiration-to-expiration (I:E) time duration ratio of 1:2 and without positive end-expiratory pressure.
The lungs of the animals were opened with five breaths at 35 cmH2O and then the ventilator pressure was adjusted to maintain tidal volumes of 6–7 mL per kg body weight, while keeping the pressure under 30 cmH2O. After 15 minutes of ventilation, the animals were shortly disconnected from the respirator and instilled with 5 mL/kg body weight of physiological saline (NaCl) (n = 2) or one of three different concentrations of P. aeruginosa: 1.7 × 108 (corresponding to OD595 ≈0.12) (n = 2), 1.7 × 109 (OD595 ≈1.31) (n = 3), and 3.4 × 109 CFU/mL (OD595 ≈1.8) (n = 2), which corresponds to an infectious dose of 8.25 × 108, 8.25 × 109, and 1.70 × 1010 CFU/kg body weight, respectively. After instillation of bacteria or NaCl, the lungs of the animals were opened with another five breaths at 35 cmH2O.
During the rest of the experiment, the tidal volumes were individually adjusted to 6–7 mL per kg body weight. After approximately 2 hours of ventilation, the animals were given a second dose of pentobarbital and glucose. Four hours postinfection, the animals were sacrificed and the lungs were removed and weighed. The right lungs were immersed in 4% formalin, embedded in paraffin, and sectioned. The sections were stained with hematoxylin and eosin and observed by light microscopy. The left lungs were homogenized, using ULTRA Turrax T10 (IKA, Staufen, Germany), serially diluted with PBS, and plated. The plates were incubated overnight at 37°C, the CFUs were counted, and the CFUs per gram of lung were calculated. Statistical analyses of the CFU counts between the different groups were evaluated by t-test using GraphPad software.
Experiments with polymyxin E and animal-derived surfactant in P. aeruginosa-infected rabbits
The experiments were conducted as described above, with the following exceptions: the near-term rabbits were infected immediately after being connected to the ventilator in the multiplethysmograph system. The bacterial solution was adjusted to an OD595 = 0.09 ± 0.01 corresponding to 8 ± 4 × 107 CFU/mL, and an infectious dose of 4 ± 2 × 108 CFU/kg body weight. All four treatment groups received essentially the same bacterial dose (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/jamp).
After 30 minutes of ventilation, the animals were briefly disconnected from the ventilator and instilled with 2.5 mL/kg body weight of either physiological saline (NaCl) (n = 10), poractant alfa 80 mg/mL (surfactant) (n = 10), 0.8 mg/mL polymyxin E in poractant alfa 80 mg/mL (surfactant + polymyxin E) (n = 10), or 0.8 mg/mL polymyxin E in physiological saline (NaCl + polymyxin E) (n = 10). Four hours postinfection, the animals were sacrificed and the lungs and left kidney were removed and weighed. Lungs and kidneys were separately homogenized, diluted with PBS, plated, and incubated overnight at 37°C, and thereafter, the number of CFUs was counted.
Forty-two fetuses with a body weight equal to or higher than 20 g were primarily included in the study and assigned to the four treatment groups sequentially in the order of delivery. Two animals showed a heart rate lower than 60 beats per minute during the first 3.5 hours of the mechanical ventilation and were excluded.
Ethical approval
The animal experiments were performed according to the ethical permit N174/14 obtained from Stockholms Norra Djurförsöksetiska Nämnd.
Quantification of polymyxin E in the presence of poractant alfa and in kidneys
Mixtures of 0.1 mg/mL polymyxin E in diluted, 10 mg/mL, poractant alfa (Curo + PxE) and 0.1 mg/mL polymyxin E in physiological saline (PxE) were centrifuged at 1000 g or 100,000 g for 1 h. The supernatants and a sample of 0.1 mg/mL polymyxin E in physiological saline that had not been centrifuged were diluted to a concentration of 180 ng/mL polymyxin E, assuming that all polymyxin E remained in the supernatant, and the concentration of polymyxin E was then meausered using an ELISA Kit from Bio Scientific Corporation (Austin, TX).
Kidneys from animals that had been treated with polymyxin E + Curosurf or polymyxin E in saline were homogenized, and polymyxin E was measured with the same ELISA kit as above, according to the manufacturer's instructions. Kidneys from animals that had not received polymyxin E were spiked with polymyxin E and the detection yield was used to correct the values obtained for kidneys from polymyxin E-treated animals.
Results
Effect of poractant alfa on the activity of polymyxin E and gentamicin in vitro
P. aeruginosa showed similar growth rate during 6 hours of incubation in PBS and in the presence of 40 mg/mL of pulmonary surfactant, and both antibiotics tested (polymyxin E and gentamicin) presented concentration-dependent bactericidal effects (Fig. 1). After 4 and 6 hours of incubation in the presence of 2 μg/mL polymyxin E, there was a significantly higher number of CFUs in the surfactant group compared to the PBS group (Fig. 1D). Likewise, with 5 μg/mL polymyxin E, after 2 and 4 hours of incubation, the number of CFUs in the lung surfactant group was higher than in the PBS group (Fig. 1E). At 10 μg/mL polymyxin E, there was no significant difference between the surfactant and PBS groups (Fig. 1F). In contrast to polymyxin E, gentamicin was unaffected by the presence of surfactant at all concentrations tested (Fig. 1H–K).
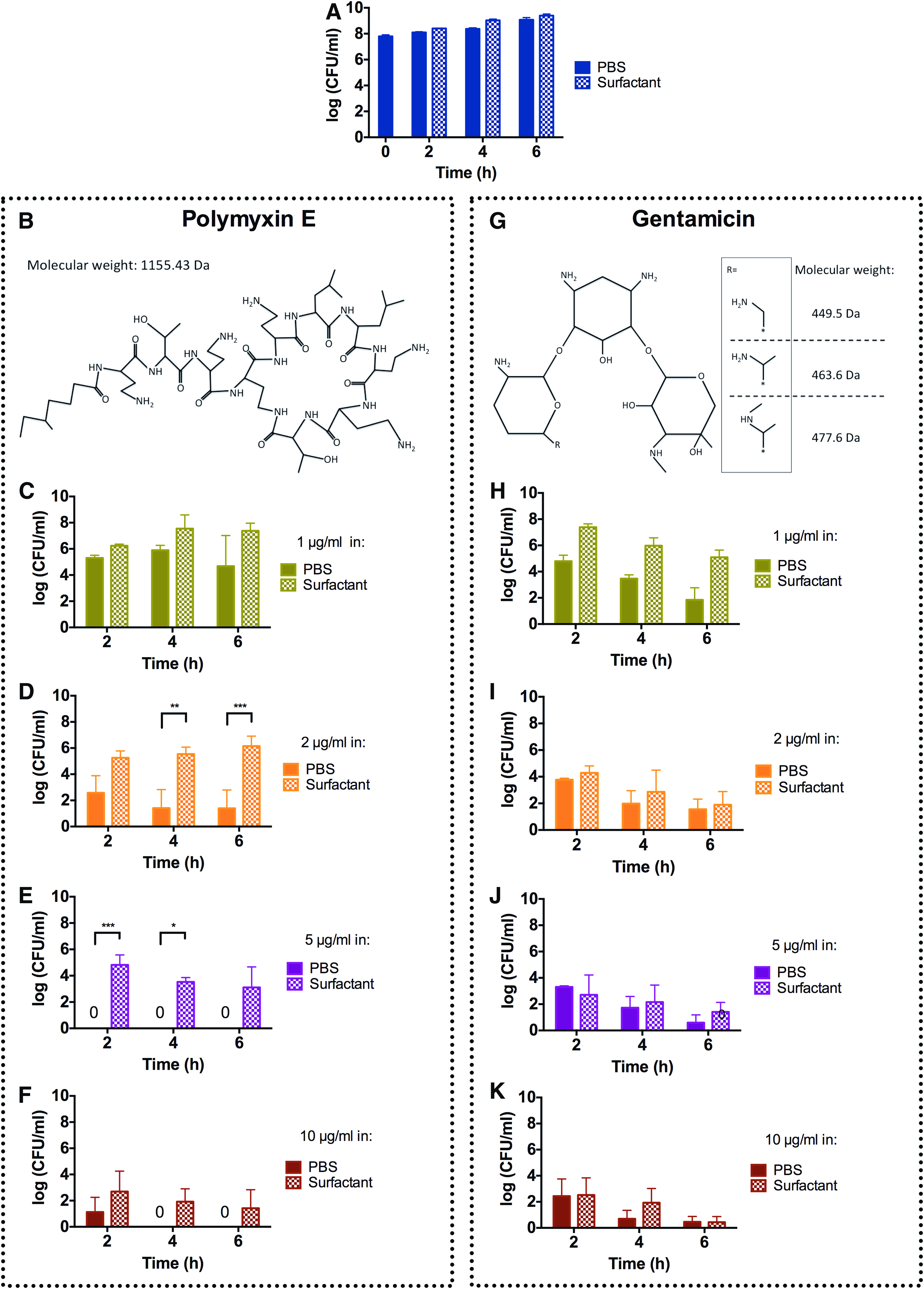
Effects of polymyxin E and gentamicin in the presence or absence of pulmonary surfactant against Pseudomonas aeruginosa in vitro. The logarithmic CFU values are plotted for 0, 2, 4, and 6 hours of incubation. The panels show results for no antibiotic
Evaluation of different infectious doses of P. aeruginosa
The animals were randomly assigned to instillation of saline, or different doses of P. aeruginosa. After 4 hours of ventilation, the animals were sacrificed and one lung was used to determine the bacterial content and the other lung was used for histopathological examination (Fig. 2). The animals instilled with saline showed normal macroscopic and histological appearances of the lungs and a minimal inflammatory reaction, as well as absence of bacteria in the lungs (Fig. 2A, F). The lungs of the animals receiving the highest dose of bacteria showed signs of massive hemorrhage, edema, and infiltration of inflammatory cells (Fig. 2D, I). Even the animals that received the lowest dose (8.25 × 108 CFU/kg) presented hemorrhage and infiltration of inflammatory cells in the lung parenchyma as well as macroscopic signs of lung injury (Fig. 2B, G).

Identification of a suitable infectious dose for the development of an animal model of P. aeruginosa pneumonia. Histological sections of right lungs of premature rabbit fetuses
All the animals that received bacteria showed high amount of bacteria in the lungs. Based on these results, an infectious dose of 4 × 108 CFU/kg was chosen for the treatment study. Notably, we observed that the infection seemed unevenly distributed in the lungs (Fig. 2G–I). Therefore, in the subsequent experiments, we decided to use both lungs for determination of bacterial counts.
Effects of polymyxin E instilled with and without poractant alfa against P. aeruginosa pneumonia
The animals were randomly allocated to one of the four treatment groups and instilled with P. aeruginosa (Fig. 3). The CFU counts of the lung homogenates at the end of the experiment showed no difference between animals in the saline- and the surfactant-treated groups. The animals treated with antibiotic (surfactant + polymyxin E or NaCl + polymyxin E) showed significantly reduced numbers of CFUs in the lungs compared to the animals treated without antibiotics (NaCl and surfactant). The animals treated with surfactant + polymyxin E had a significantly reduced number of CFUs in the lungs at the end of the experiment compared to the animals treated with NaCl + polymyxin E (p-value = 0.0067).
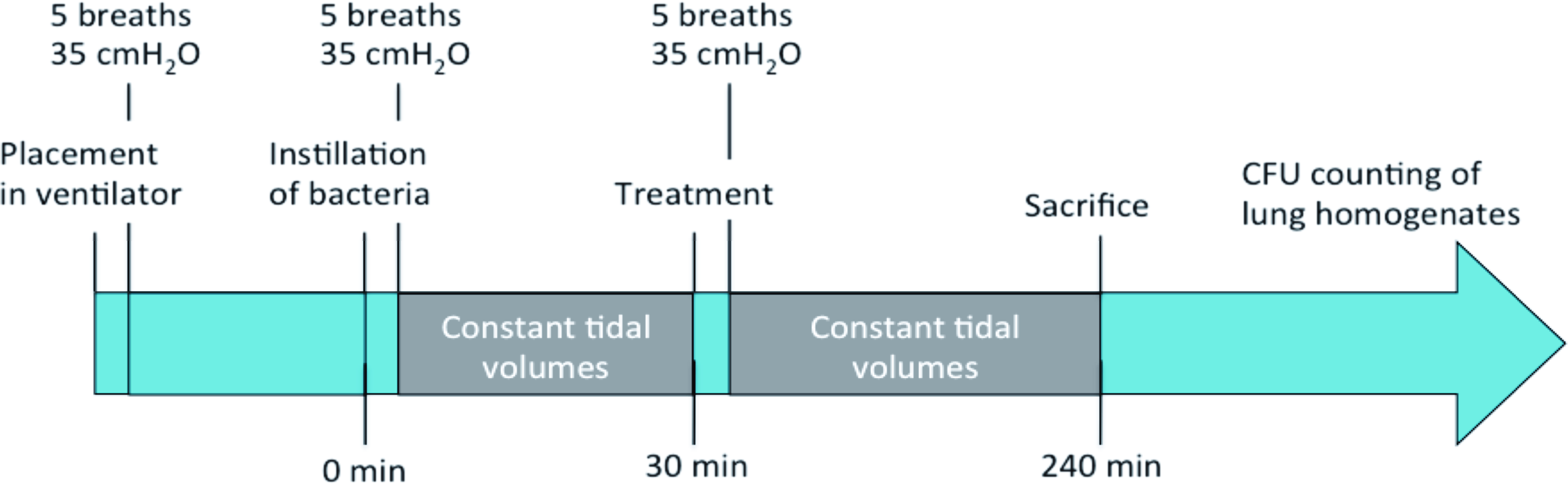
Experimental setup for testing the effects of polymyxin E with and without lung surfactant against P. aeruginosa pneumonia in premature rabbit fetuses. The rabbits were delivered by cesarean section after 29 days of gestation, tracheotomized, placed in the ventilator, and ventilated for 4 hours. Color images available online.
Thus, poractant alfa in combination with polymyxin E was more efficient in reducing the number of bacteria in the lungs compared to NaCl + polymyxin E (Fig. 4). To evaluate whether there was a systemic spreading of bacteria in the infected animals, one kidney per animal was homogenized for bacterial count. Low numbers of bacteria (2–5 × 102 CFU/g of kidney) were present in homogenates from 3 out of the 40 animals in the study. Two of these animals belonged to the saline-treated group and one animal belonged to the group treated with NaCl + polymyxin E. In all groups, the compliance was stable during the experiment (Fig. 5). Finally, the macroscopic appearances of the lungs confirmed the findings from the pilot study, showing uneven distribution of affected areas in the lungs (Supplementary Fig. S2).
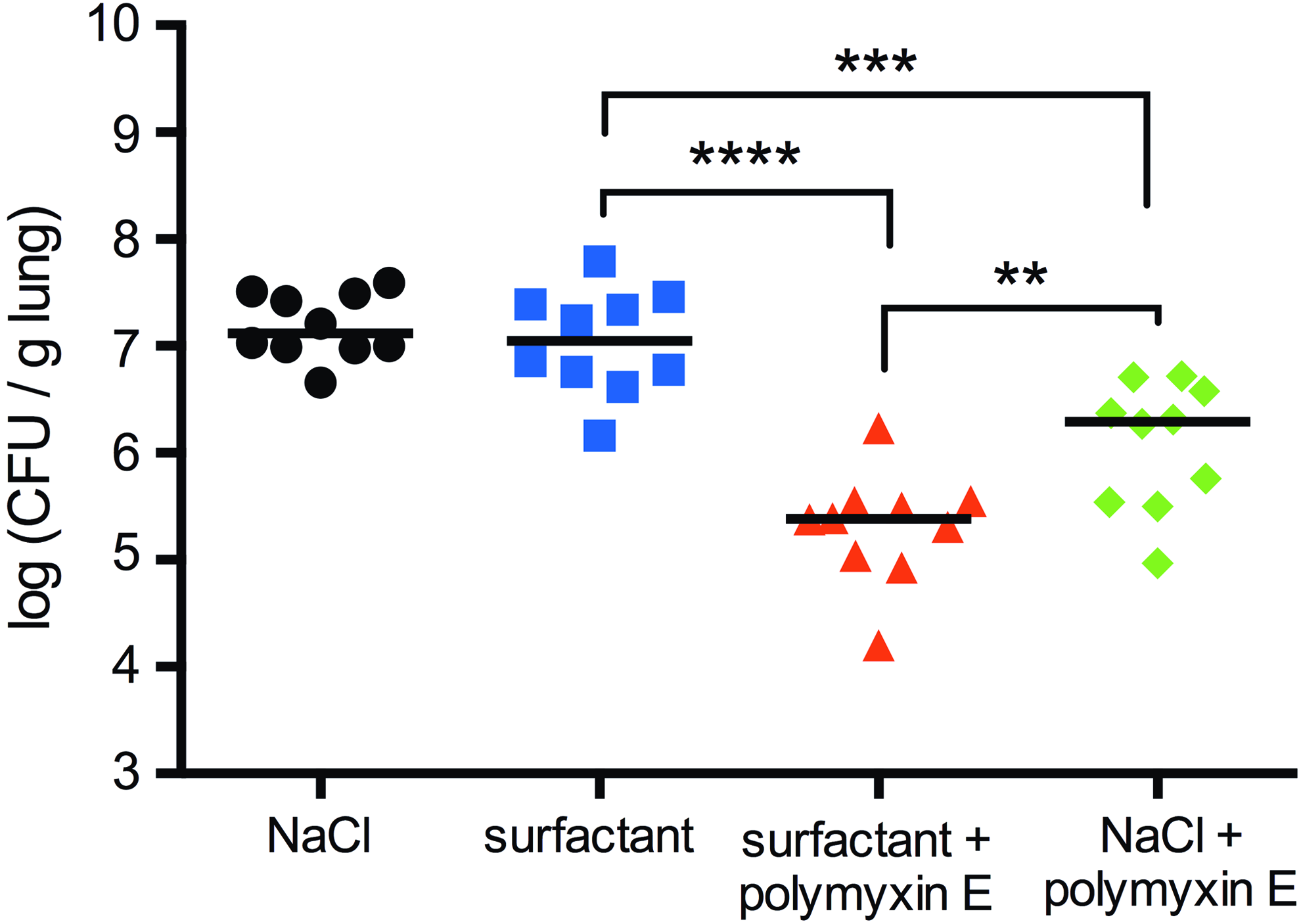
Logarithmic plot of CFU per gram lung tissue after different treatments; 30 minutes postinfection, the animals were instilled with 2.5 mL per kg body weight of NaCl (black circles), surfactant (blue squares), surfactant + polymyxin E (red triangles), or NaCl + polymyxin E (green diamonds) and were ventilated for an additional 210 minutes before homogenization and plating of the samples. N = 10 in each group. The black lines represent the median of each group. **p ≤ 0.01, ***p ≤ 0.001, and ****p ≤ 0.0001. Color images available online.
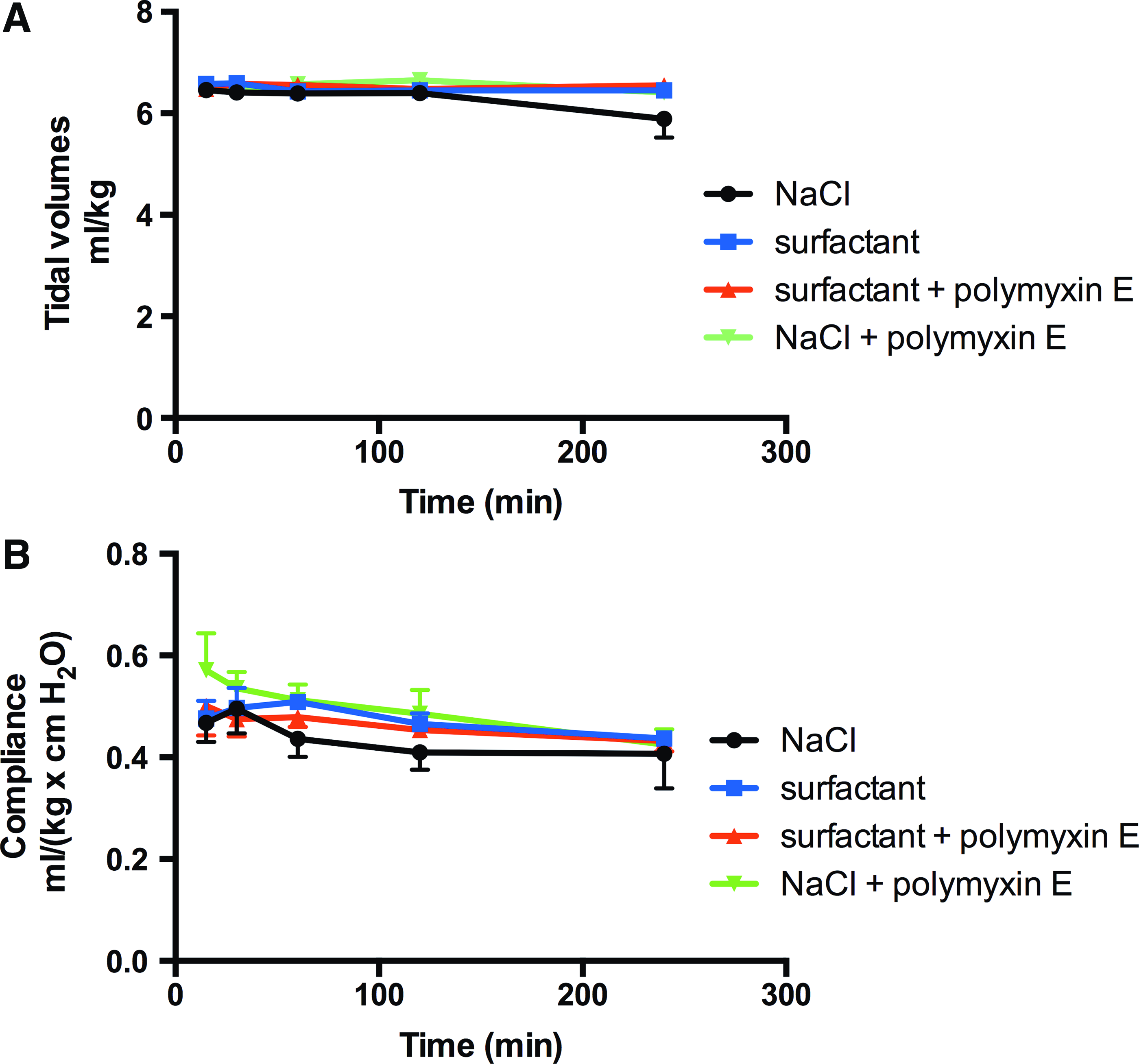
Tidal volumes
Association between poractant alfa and polymyxin E
The amount of polymyxin E measured in the supernatants after centrifugation of mixtures of surfactant and polymyxin E was lower than in mixtures of polymyxin E in saline (Supplementary Fig. S3). Surfactant vesicles are pelleted under the centrifugation procedure used and the results thus indicate that polymyxin E partly associated with poractant alfa.
Discussion
This study was performed to investigate the potential benefit of using exogenous surfactant for pulmonary delivery of antibiotics. We chose to study P. aeruginosa as it belongs to the group of bacteria that are developing MDR, and thus are increasingly difficult to treat.(24) Furthermore, it is a common pathogen causing chronic lung infections in compromised patients, like those with ventilator-associated pneumonia, cystic fibrosis, bronchiectasis, and chronic obstructive pulmonary disease. The treatment strategy can optionally include inhalation of different antibiotics, like nebulized polymyxin E.
In this manner, bacteria in the airways, which are difficult to reach by systemically administered antibiotics, can be more efficiently targeted. However, a potential problem is suboptimal spread to distant foci that often are occluded by mucus and lung parenchyma injury.(25) Another antibiotic used to fight P. aeruginosa infection is gentamicin that belongs to the aminoglycosides. Although polymyxin E is more toxic than gentamicin, both are associated with severe systemic side effects when given systemically(23,26); therefore, an improved method for local administration to the lung would be beneficial.
The activity of polymyxin E and gentamicin in the absence or presence of poractant alfa was evaluated using a P. aeruginosa in vitro assay. The presence of 40 mg/mL of the modified natural surfactant did not influence bacterial growth in vitro, indicating that the surfactant does not have intrinsic antibacterial properties or promote growth of P. aeruginosa. The surfactant had no statistically significant effects on the activity of gentamicin in vitro (Fig. 1H–K).
These results are in line with a previously published study where the activity of amikacin (another aminoglycoside) was shown to be unaffected in vitro.(11) However, the aminoglycoside tobramycin shows reduced antibacterial activity in vitro in the presence of pulmonary surfactant.(5) Interestingly, in vivo, the effects of the aminoglycosides amikacin and tobramycin are increased in the presence of pulmonary surfactant.(13–15)
In contrast to the results obtained with gentamicin, the antimicrobial activity of the cyclic lipopeptide polymyxin E was attenuated by surfactant at all concentrations tested in vitro, except for the highest concentration at which the effect of polymyxin E was unaffected when mixed with surfactant (Fig. 1C–F). These findings correlate with the ones of Schwameis et al. where it was shown that polymyxin E activity is reduced in the presence of pulmonary surfactant in vitro.(12)
The mechanisms of action of polymyxin E and gentamicin are different, as are their physicochemical properties (Fig. 1B, G), which could explain the different behavior of gentamicin and polymyxin E in the presence of lung surfactant in vitro. Aminoglycosides like gentamicin are polar and cationic with a net charge of approximately +3.5 at pH 7.4.(27) They act by interfering with bacterial translation by irreversible binding to the bacterial 30S ribosome subunit.(28) The polycationic polymyxin E is amphiphilic, due to the presence of a short fatty acyl chain and a polycationic head, and forms an oligomer that interacts with negative charges of Lipid A in LPS, which leads to destabilization of the outer membrane causing membrane permeabilization and disruption.(29)
It is plausible that polymyxin E also interacts with the phospholipids of the pulmonary surfactant, reducing its availability to interact with the bacteria and decreasing its antibacterial activity in vitro. This notion is corroborated by findings that polymyxins bind phospholipids of whole cell homogenates of different mammalian tissues, and lose antibacterial activity.(30) Furthermore, a natural derived surfactant preparation also inhibits the activity of daptomycin, another cyclic lipopeptide able to bind phospholipids.(10)
Instillation of poractant alfa alone did not affect the number of CFUs in the lungs (Fig. 4), indicating that the surfactant has no intrinsic effects on P. aeruginosa growth. The surfactant + polymyxin E mixture was more efficient in reducing the number of bacteria in the lungs compared to the NaCl + polymyxin E mixture. Since poractant alfa reduced the effect of polymyxin E in vitro, we suggest that the beneficial effects observed in vivo can be attributed to more efficient spreading of the antibiotic, mediated by an interaction with the surfactant phospholipids, although other explanations cannot be ruled out. For example, pulmonary surfactant could help to open up areas with atelectasis, thus positively influencing the distribution of the antibiotic.
A more efficient spreading, independent of underlying mechanism, would allow polymyxin E to reach distant foci of bacterial infection more effectively. Polymyxin E apparently partly associates with poractant alfa, resulting in that less polymyxin E is found in the supernatant after centrifugation of polymyxin E mixed with poractant alfa than after centrifugation of polymyxin E in saline (Supplementary Fig. S3). This supports the possibility that interactions between polymyxin E and poractant alfa promote efficient spreading, but further studies are required to explain the mechanisms underlying the enhanced bacterial killing observed for polymyxin E/poractant alfa mixtures.
We also analyzed the amount of polymyxin E in kidneys from three animals that had received polymyxin E + Curosurf and in three animals that received polymyxin in saline. In all cases, the amount detected in the kidneys were <2% of the amount of polymyxin given. This suggests that local administration of polymyxin E to the lungs gives lower systemic spreading compared to when polymyxin is administered intravenously, for which about 14% was detected in the kidneys.(31)
The effect of polymyxin E in combination with pulmonary surfactant against P. aeruginosa in animal models has not been studied before. However, in a similar study of Escherichia coli lung infection in near-term rabbits, where polymyxin E was administrated before the bacteria, it was shown that polymyxin E mixed with lung surfactant is more effective in reducing the amount of bacteria in the lungs than polymyxin E alone.(16) The same outcome was observed for polymyxin B, another cyclic lipopeptide with a similar mechanism of action to polymyxin E binding the LPS of Gram-negative bacteria.(16, 32)
In conclusion, treatment of P. aeruginosa-infected rabbits with surfactant + polymyxin E resulted in significantly reduced bacterial load in the lungs compared to treatment with NaCl + polymyxin E. This result suggests that polymyxin E combined with exogenous pulmonary surfactant has the potential to be used for future treatments of bacterial pneumonia.
Footnotes
Acknowledgments
We thank Marie Haegerstrand-Björkman and Bim Linderholm for technical assistance. The work was supported by the following grants; Swedish Research Council (2014–2408), Center for innovative medicine (CIMED), and FORMAS (2015-629), Chiesi Farmaceutici, Parma, Italy.
Author Disclosure Statement
The authors of this article obtained financial support from Chiesi Farmaceutici, Parma, Italy.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.