
Abstract
Interferon (IFN) is believed to be one of the most effective anti-melanoma agents. Specifically, IFN-β has the ability to induce apoptosis of melanoma cells. Induction of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) has also been suggested to have a critical role in IFN-β-induced apoptosis. To characterize the signaling pathway involved in IFN-β-induced apoptosis, we analyzed the biological effects of IFN-β on the cell death and caspase activation of melanoma cells. IFN-sensitive cell lines, MM418, SK-mel-23, and SK-mel-118, showed increased apoptotic populations correlated with the activation of caspase-2 and caspase-3 by IFN-β. IFN-β-induced apoptosis was significantly suppressed by inhibitors for caspase-2 or caspase-3, but not by inhibitors for caspase-8 or caspase-9 in these cell lines. TRAIL expression was observed in IFN-β-treated cells of SK-mel-23 and SK-mel-118, but not in those cells of MM418, which showed massive IFN-β-induced apoptosis and resistance to exogenous TRAIL-mediated apoptosis. G361 was resistant to IFN-β-induced apoptosis but sensitive to exogenous TRAIL-mediated apoptosis. Furthermore, IFN-β pretreatment significantly increased the sensitivity against exogenous TRAIL-mediated apoptosis and activation of caspase-2 in G361. These results suggested that caspase-2 activation is commonly associated with induction of IFN-β-induced apoptosis in IFN-β-sensitive melanoma cells.
Introduction
T
Although the mechanisms of IFN-mediated cell death have not been fully elucidated, IFN is known as an effective anti-tumor agent (Fisher and others 1985; Pfeffer and others 1998; Stark and others 1998). It was reported that IFN-β induced apoptosis in melanoma cells more significantly than IFN-α and -γ did (Chawla-Sarkar and others 2001; Leaman and others 2003; Merchant and others 2004). It induces many biological responses by regulating IFN-stimulated genes (ISGs) (Chawla-Sarkar and others 2003). Several ISGs, such as double-stranded RNA-activated protein kinase (PKR), myxovirus resistance protein A (MxA), melanoma differentiation-associated gene-5 (MDA-5), and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL), are related to the anti-tumor effects of IFN (Leaman and others 2003; Kang and others 2004).
Recently, it has been clarified that IFN has apoptotic effects on several tumors. TRAIL, known as Apo2 ligand, is also a member of the TNF family of transmembrane proteins, which leads tumor cells to apoptosis by stimulating death receptors (DRs) (Griffith and others 1998; Kimberley and Screaton 2004; Zhang and Fang 2005). It has been suggested that the involvement of TRAIL is essential for the apoptotic cascade induced by IFNs in certain melanoma cell lines, as well as in other tumor cell lines (Chawla-Sarkar and others 2001; Chen and others 2001; Meng and El-Deiry 2001; Morrison and others 2005). Furthermore, other ISGs, such as Ras association domain family 1A (RASSF1A) and XIAP (X-linked inhibition of apoptosis protein)-associated protein (XAF1), also contribute to IFN-β-induced and TRAIL-induced apoptosis (Chawla-Sarkar and others 2003; Reu and others 2006).
Apoptosis can be included by 2 signaling pathways. One is through apoptosis-inducing ligands such as Fas ligand, TRAIL, and TNF-α-stimulating DRs. The other is through cytochrome c release from mitochondria. In these pathways, it is evident that the functions of caspase-3, −8, and −9 are essential. Caspase-2, which is considered to be an initiator caspase and involved in upper stream of the apoptotic pathways, is located diversely in the cytoplasm and nuclei. However, the biological role of caspase-2 remains controversial compared with those of other caspases (Zhivotovsky and others 1999; van Loo and others 2002; Zhivotovsky and Orrenius, 2005). The correlation between TRAIL and caspase-2 has been indicated recently in TRAIL-mediated apoptotic pathways, in which caspase-2 processes procaspase-8 or cleaves Bid, a proapoptotic Bcl-2 family member (Wagner and others 2004; Shin and others 2005). Although the interaction among apoptosis-inducing ligands, IFNs, and caspase-2 has recently been clarified to some extent, the role of caspase-2 in IFN-induced apoptosis remains to be elucidated.
In this study we were interested in the activities of caspases, especially caspase-2 in the IFN-β-induced apoptotic pathway. Therefore, we examined 2 classes of human melanoma cell lines that were sensitive and resistant to apoptosis induced by IFN-β treatment.
Materials and Methods
Cell cultures and reagents
The human melanoma cell lines, SK-mel-118, SK-mel-23, MM418, and G361 (Yamashita and others 2001), were grown in Dulbecco's modified Eagle's medium (DMEM) supplemented with 5% fetal bovine serum and antibiotics in 5% CO2 incubators at 37°C. SK-mel-23 and MM418 are pigmented cell lines, and the others are non-pigmented cell lines. IFN-α2 was purchased from Serotec (Oxford, UK). IFN-β was supplied by Mochida Pharmaceutical (Tokyo, Japan). Cells were treated with 1,000 IU/mL of IFNs for different time periods depending on the experiment. Recombinant human TRAIL (R&D Systems, Minneapolis, MN, USA) was used at the final concentration of 100 ng/mL. Recombinant human TNF-α (R&D systems, Minneapolis, MN, USA) was used at the final concentration of 50 ng/mL. The anti-Fas (CD95) antibody (MBL, Nagoya, Japan) was used at the final concentration of 500 ng/mL.
Cell viability assay
Cells were plated in 100 mL of medium in 96-well plates in triplicate with 1,000 cells/well. After 24 h, cells were treated with IFN-α2, or IFN-β (72 h), or treated with IFN-β (24 h) followed by addition of TRAIL, TNF-α, or the anti-Fas (CD95) antibody (48 h) after PBS washing. At 48 h after plating, cells were also treated with TRAIL, TNF-α, or the anti-Fas (CD95) antibody (48 h) to be compared with IFN-β-pretreated cells. The viability of treated cells was determined with the MTT assay, which is a formazan-formation assay, using the Cell Counting Kit-8 (DOJINDO, Kumamoto, Japan) according to the manufacturer's protocol.
Semiquantitative reverse transcriptase-polymerase chain reaction (RT-PCR)
Total RNA was extracted from cells using an RNeasy mini kit (Qiagen, Hilden, Germany). Complementary DNA was synthesized using Super Script™-III Rnase H (Invitrogen, Carlsbad, CA, USA) from total extracted RNA (50 ng). The quantitative PCR was validated by the linearity of the determination curves at various concentrations of cDNA. Specific primers (Sigma-Genosys, Ishikari, Japan) were constructed as follows: 5′-TTGGCTCAGGTGG ATTTGG-3′ and 5′-GGCTTTTCTTCCACACAGTC-3′ for PKR, 5′-GCATCCCACCCTCTATTACT-3′ and 5′-TGTCTT CAGTTCCTTTGTCC-3′ for MxA, 5′-GGAAGTACAATGAG GCCCTACAAA-3′, 5′-TCCTCAGTCCTAGTATATTGCTCC-3′ for mda-5, 5′-GGCTATGATGGAGGTCCAGG-3′ and 5′-GGTCCATGTCTATCAAGTGCTC-3′ for TRAIL, 5′-AGC GTGCCAACGCGCTGCGCAT-3′ and 5′-CAGGCTCGTC CACGTTCGTGTC-3′ for RASSF1A, 5′-GCCTGCGGTTCCTG GTCCTG-3′ and 5′-GCTGGGCGAGCATGCGGTGC-3′ for XAF1, 5′-CCAACAAGACCTAGCTCCCCAGC-3′ and 5′-AA GACTACGGCTGCAACTGTGACTCC-3′ for DR4, 5′-GTCCT GCTGCAGGTCGTACC-3′ and 5′-GATGTCACTCCAGG GCGTAC-3′ for DR5, 5′-TCCACCACCCTGTTGCTGTA-3′ and 5′-ACCACAGTCCATGCCATCAC-3′ for glyceraldehyde-3-phosphate dehydrogenase (GAPDH). PCR using Taq DNA polymerase (Promega, Wisconsin, WI) as follows: denaturation, 94°C for 30 s; annealing, 55°C for 30 s; and extension, 72°C for 1 min. The number of cycles was 35.
Fluorescence-activated cell sorting analysis
Analysis of TUNEL assay of apoptotic cells was done by fluorescence-activated cell sorting (FACS). Cells were plated in 8 mL of medium in 10-cm dishes with 2 × 104 cells/mL. After 72 h, cells were harvested by scraping. For TUNEL assay, cells were assayed using the Apo-BrdU In situ DNA fragmentation Assay Kit (MBL, Nagoya, Japan) according to manufacturer's protocol. These stained cells were analyzed with a FACS Caliber and Cell Quest software (Becton Dickinson, San Jose, CA).
Caspase activity assay
Cells were plated in 8 mL of medium in 10-cm dishes with 2 × 104 cells/mL. After 24 h, cells were treated with IFN-α2 and IFN-β (48 h), or with IFN-β (24 h) followed by PBS washing and addition of TRAIL, TNF-α, and the anti-Fas (CD95) antibody (24 h). At 48 h after cell plating, cells were also treated with TRAIL, TNF-α, and the anti-Fas (CD95) antibody (24 h) to be compared with IFN-β-pretreated cells. Cells were harvested by scraping, and were assayed with a commercially available caspase-2, −3, −8, −9, and −10 fluorometric assay kit (MBL, Nagoya, Japan) as procedure was described. Caspase activity was measured by spectrofluorometer Fluoroskan Ascent FL (Labsystems, Helsinki, Finland). The excitation wavelength was 390 nm and the emission wavelength was 510 nm. Western blotting analysis of caspase protein was performed as described elsewhere (Yamashita and others 2001). Rabbit anti-caspase-2 antibody was purchased from Santa Cruz Biotechnology (Santa Cruz, CA).
Caspase inhibitor treatment
Caspase inhibitors, Z-VDVAD-FMK (specific for caspase-2), Z-DQMD-FMK (specific for caspase-3), Z-IETD-FMK (specific for caspase-8 and granzyme B), and Z-LEHD-FMK (specific for caspase-9) were purchased from Calbiochem (San Diego, CA). The cells were treated with 1,000 IU/mL IFN-β for 48 h in the presence or absence of the caspase inhibitor at a concentration of 100 mM. Cell viability was determined with MTT assay.
Statistical analysis
Statistical comparisons were made using Student's t-test.
Results
IFN-β inhibited the viability of melanoma cell lines SK-mel-118, SK-mel-23, and MM418, but not G361.
To assess the viability of IFN-α and -β-treated melanoma cell lines, MTT assay was performed (Fig. 1A). The cell viabilities of SK-mel-118, SK-mel-23, and MM418 were reduced by IFN treatments. Detached cell death and cell growth inhibition was microscopically detected in these cell lines. The reduction of cell viability by IFN-β was significantly higher than by IFN-α2. However, slight cell growth inhibition and no detached cell death of G361 were microscopically detected by IFN treatments. The cell viability was slightly inhibited by IFN-α2 or IFN-β but no significant difference was observed between untreated cell viability and IFN-treated cell viability. And more cell death was not induced by higher dose (>1,000 IU/mL) of IFN-β in G361 and SK-mel-118 (data not shown). These results indicated that these 4 melanoma cell lines were divided into 2 groups, one in which cell viability inhibition and detached cell death was observed by IFN, and the other in which slight cell viability inhibition and no detached cell death was observed.
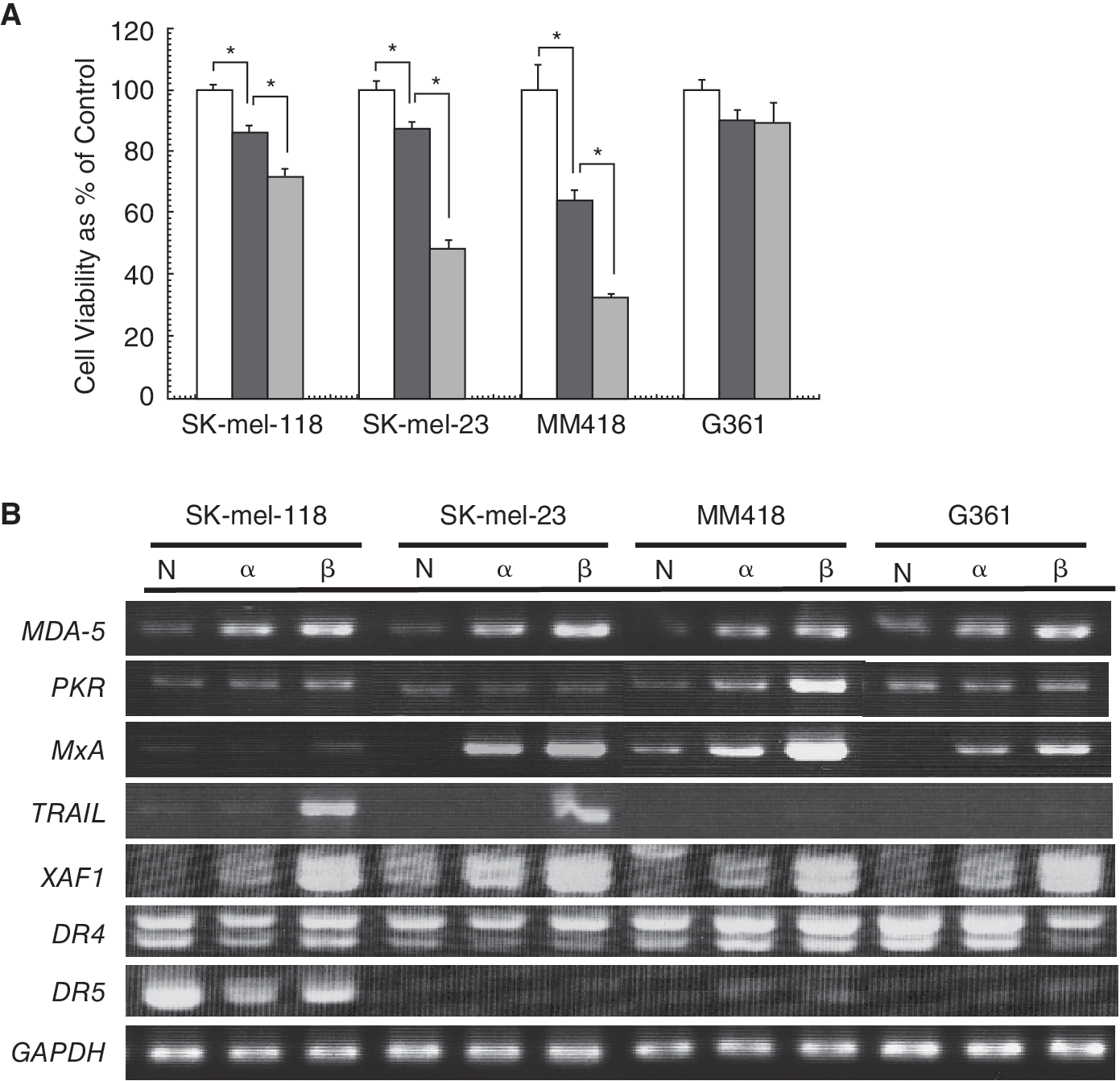
Cell viability and mRNA expression of interferon (IFN)-stimulated genes after treatment with IFN-α2 and IFN-β. (
Induction of ISGs after treatment with IFNs
To assess whether IFN treatment could transduce intracellular signaling in melanoma cell lines, we investigated mRNA expression of ISGs, which associated with cell death, such as MDA-5, PKR, MxA, TRAIL, RASSF1A, and XAF1 in IFN-treated cell lines (Fig. 1B). The mRNA expressions of MDA-5 were significantly up-regulated by IFN treatment in all the cell lines, including G361, which was resistant to the anti-viability effects of IFN. Those of PKR, MxA, and XAF1 also tended to be up-regulated in the tested cell lines. These results indicated that the IFN-signaling pathway was not suppressed in any of the tested cell lines. TRAIL-mRNA inductions of SK-mel-118 and SK-mel-23 by IFN-β treatment were markedly higher than those of MM418 and G361. RASSF1A mRNA was not detected in all cell lines. These results suggested that expression levels of any ISGs so far examined did not correlate with the degree of IFN-induced apoptosis. Furthermore, we examined expression levels of proapoptotic DRs for TRAIL. DR5 mRNA strongly expressed in SK-mel-118, which is highly sensitive to TRAIL, and decreased by treatment with IFN. DR4 mRNA was reduced by IFN treatment in G361 and it did not show any significant changes by IFN treatment in other cell lines. These results suggested that DRs for TRAIL had little to do with the effect of IFN-induced cell death.
Induction of apoptosis in melanoma cells by IFN-β treatment
We investigated the mechanism of cell death induced by IFN-β. We performed TUNEL assay in these 4 cell lines to assess whether the cell viability reduction and detached cell death by IFN-β occurred via apoptosis (Fig. 2). This assay showed that BrdU-FITC-positive cells corresponding to an apoptotic population, which underwent DNA fragmentation, increased in IFN-treated SK-mel-118, SK-mel-23, and MM418, whose cell viabilities were reduced by IFN-β in MTT assay. Apoptotic cells in G361, whose cell growth was slightly inhibited, were not increased by treatment with IFN-β (Fig. 2). It was reported that TRAIL is essential for the apoptosis cascade induced by IFN (Chawla-Sarkar and others 2001). In SK-mel-118 and SK-mel-23 that expressed TRAIL mRNA after IFN-β treatment, the induction of apoptosis seemed to be related to TRAIL expression. However, MM418 did not induce TRAIL-mRNA expression even if massive apoptosis was induced by IFN-β treatment. Thus the IFN-induced apoptotic pathway of MM418 was speculated to occur not through TRAIL induction by IFNs.
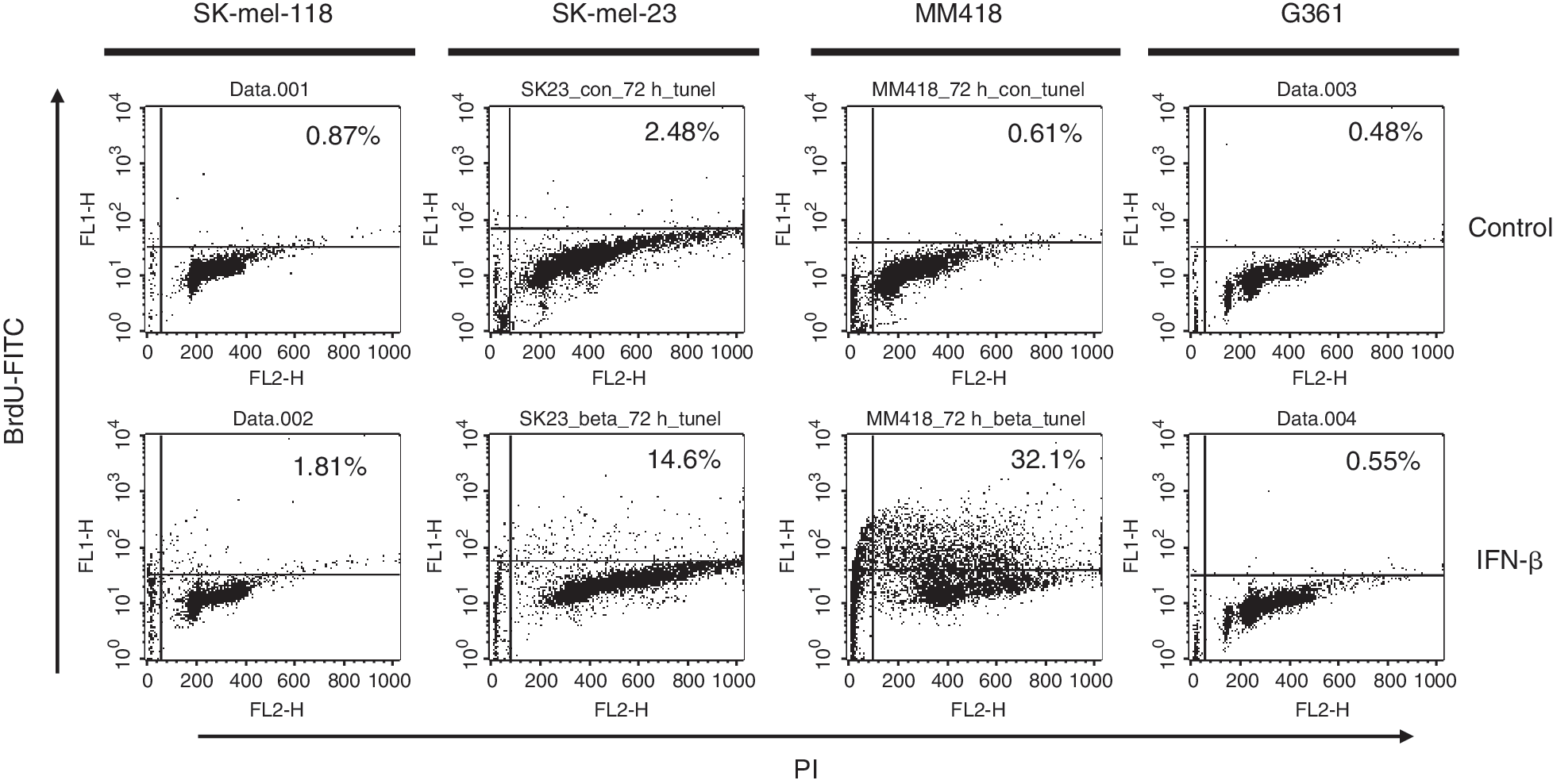
FACS analysis of DNA fragmentation by TUNEL assay. Cells were plated on 10-cm dishes with 2 × 104 cell/mL. The cell lines were treated with 1,000 IU/mL IFN-β. After 72 h, cells were harvested and fixed. They were labeled with bromo-dUTP (BrdU) by the enzyme TdT, and then stained with an FITC-labeled anti-BrdU antibody. The percentage of FITC-positive cells was assessed by FACS.
Up-regulation of caspase-2 activity was related to IFN-β-induced apoptosis in melanoma cell lines.
Activation of a caspase cascade is critical for initiation of apoptosis. We performed caspase fluorometric assay to detect up-regulation of caspase activity when apoptosis was induced in melanoma cells by IFN-β treatment. Caspase-3 activities in IFN-β-treated SK-mel-118, SK-mel-23, and MM418 showed about 2.5- to 7-fold increases, but not in G361 (Fig. 3). These results were consistent with the extent of apoptotic population of the TUNEL assay (Fig. 2). To characterize the related regulation of the caspase cascade, we furthermore analyzed activation of caspase-2, −8, −9, and −10 in IFN-β-treated cell lines. Activations of caspase-2, −3, and/or −9 were observed in IFN-β sensitive cell lines, SK-mel-118, SK-mel-23, and MM418. IFN-resistant G361 cells had no significant up-regulation of any caspase activity. Activation of caspase-2 was also confirmed by the finding that protein levels of procaspase-2 decreased by treatment with IFN-β as shown by Western blotting (Fig. 4). Active caspase-2 fragments were not detected, although degradation of procaspase-2 was able to be detected. Probably because of the sensitivity, Western blotting was not enough to detect the active caspase fragment in these IFN-β-treated cells. The levels of procaspase-2 and −3 were not up-regulated by treatment with IFN-β in all cell lines (data not shown). The IFN-β-induced apoptosis in MM418, SK-mel-23, and SK-mel-118 was significantly suppressed by the addition of inhibitor for caspase-2 or caspase-3, and these inhibitors showed equivalent anti-apoptotic effect against IFN-β-induced apoptosis. However, the inhibitors for both caspase-8 and caspase-9 did not show any anti-apoptotic effect by IFN-β (Fig. 5). These results indicated that caspase-2 and caspase-3 induced had critical role on IFN-β-induced apoptosis as an initiator and an effector for caspases, respectively.
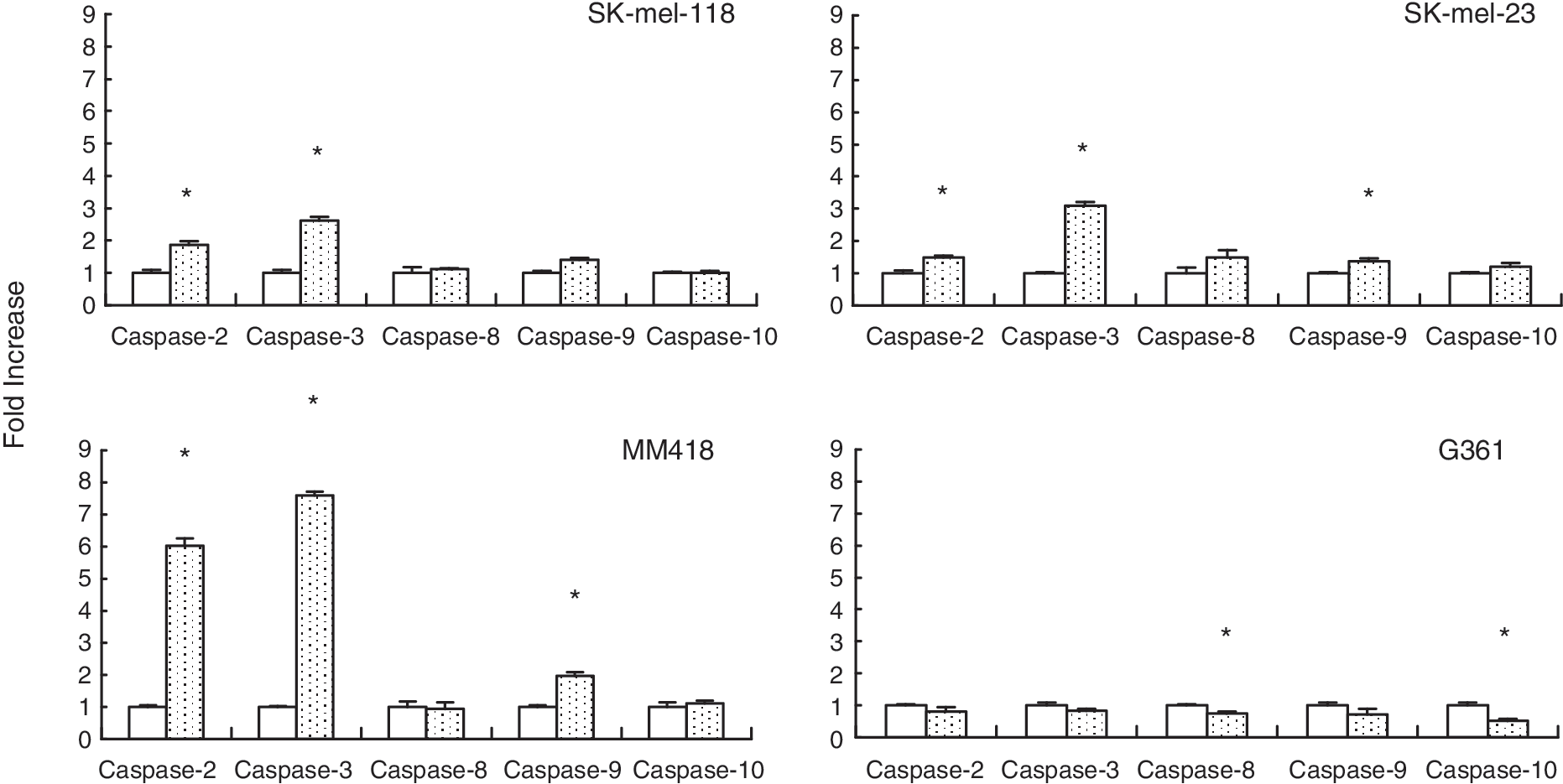
Caspase assay after treatment with interferon (IFN)-β. Cells were plated on 10-cm dishes with 2 × 104 cells/mL. The cell lines were treated with 1,000 IU/mL IFN-β in triplicate. After 48 h, cells were harvested and caspase activity was assessed by caspase fluorometric assay as described in the manufacturer's protocol. The caspase activity is expressed as the fold increase compared with the untreated cells. The results are expressed as relative values (mean ± S.D.) to untreated controls. Untreated: empty column, IFN-β: dotted column. *Significant difference (P < 0.01) from the value in the untreated control.
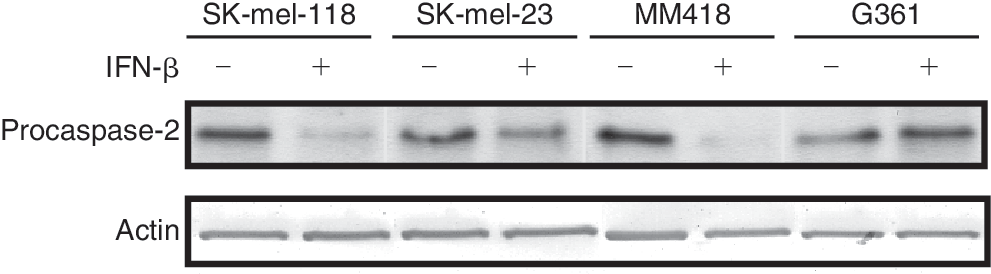
Activation of caspase-2 detected with degradation of procaspase-2 protein by Western blotting. The cells were treated with interferon (IFN)-β at 1,000 IU/mL for 48 h. The treated cells were lysed, and processed to Western blotting.
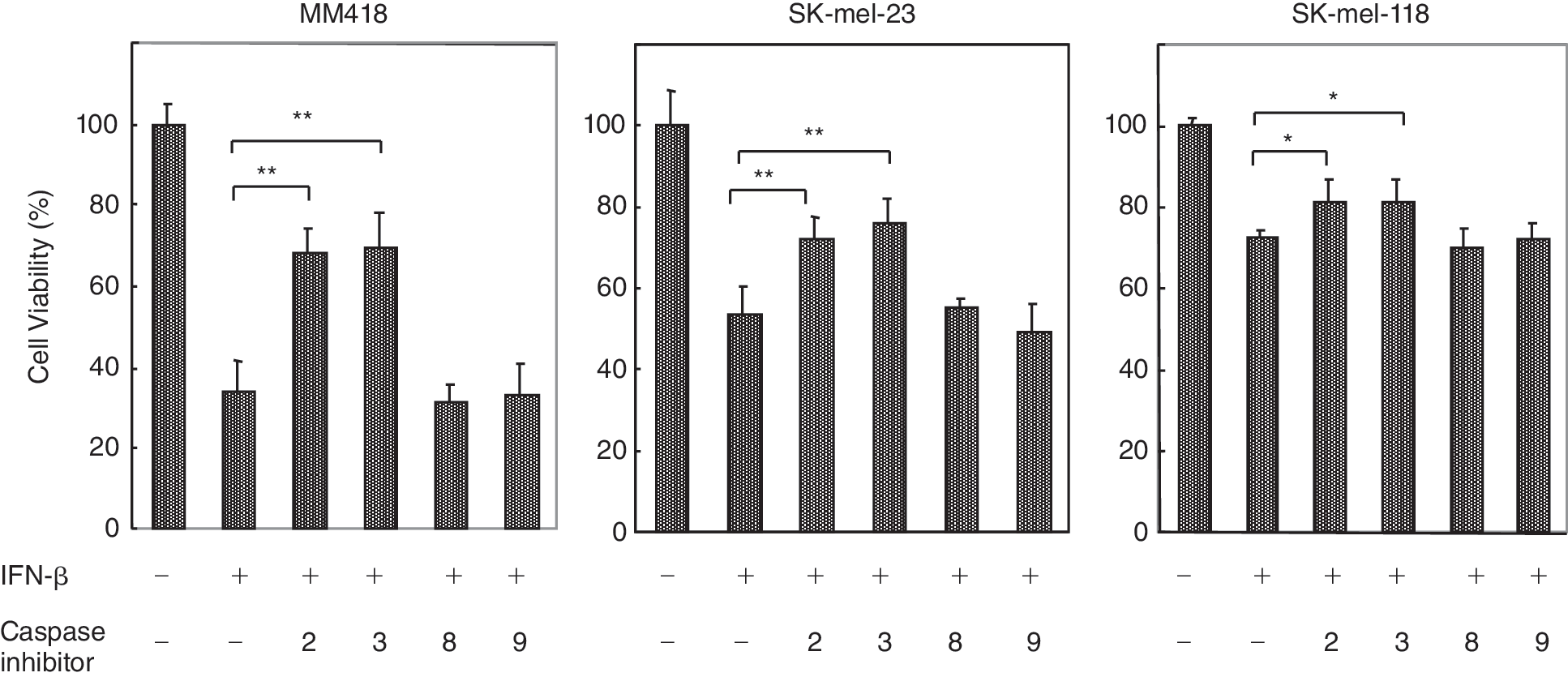
Effect of caspase inhibitors on interferon (IFN)-β-induced apoptosis. The cells were treated with IFN-β at 1,000 IU/mL in the presence of caspase inhibitor (specific for caspase-2, −3, −8, or −9) at a concentration of 100 mM. After 48 h treatment, cell viability was determined by MTT assay. Each experiment was performed in quadraplicate. The results are expressed as relative values (mean ± S.D.) to untreated controls. **P < 0.01, *P < 0.05.
Exogenous TRAIL caused apoptosis in melanoma cells
To elucidate the mechanism of IFN resistance in melanoma cells, we treated the 4 melanoma cell lines by adding exogenous apoptosis-inducing ligands (Fig. 4). When G361 and SK-mel-118 were treated with TRAIL, TNF-α, or an anti-Fas (CD95) antibody, TRAIL significantly reduced their cell viability. The anti-Fas (CD95) antibody also reduced the cell viability of SK-mel-118. The cell viabilities of SK-mel-23 and MM418 were not influenced with treatment of exogenous apoptosis-inducing ligands (Fig. 6). We also examined the effect of anti-TRAIL antibodies on the IFN-β-induced apoptosis. Anti-TRAIL antibody slightly suppressed IFN-β-induced apoptosis but did not show significant anti-apoptotic effect induced by IFN-β in SK-mel-118 and SK-mel-23. IFN-β-induced apoptosis was not suppressed by anti-TRAIL antibody in MM418 (data not shown). Furthermore, the cell viability of those tested cells was assessed after treatment with IFN-β, followed by apoptosis-inducing ligands treatment. IFN-β pretreatment followed by TRAIL stimulation significantly reduced the cell viability compared with TRAIL stimulation alone in G361 (Fig. 6). In IFN-sensitive melanoma cell lines, IFN-β pretreatment followed by TRAIL stimulation reduced the cell viability of SK-mel-23 compared with IFN-β treatment alone, although exogenous apoptosis-inducing ligands treatment did not reduce its cell viability. However, IFN-β pretreatment followed by any apoptosis-inducing ligands did not reduce the cell viability of MM418 compared with IFN-β treatment alone. In IFN-resistance G361 cells, IFN-β pretreatment followed by TRAIL stimulation significantly reduced cell viability compared with that of TRAIL treatment alone.
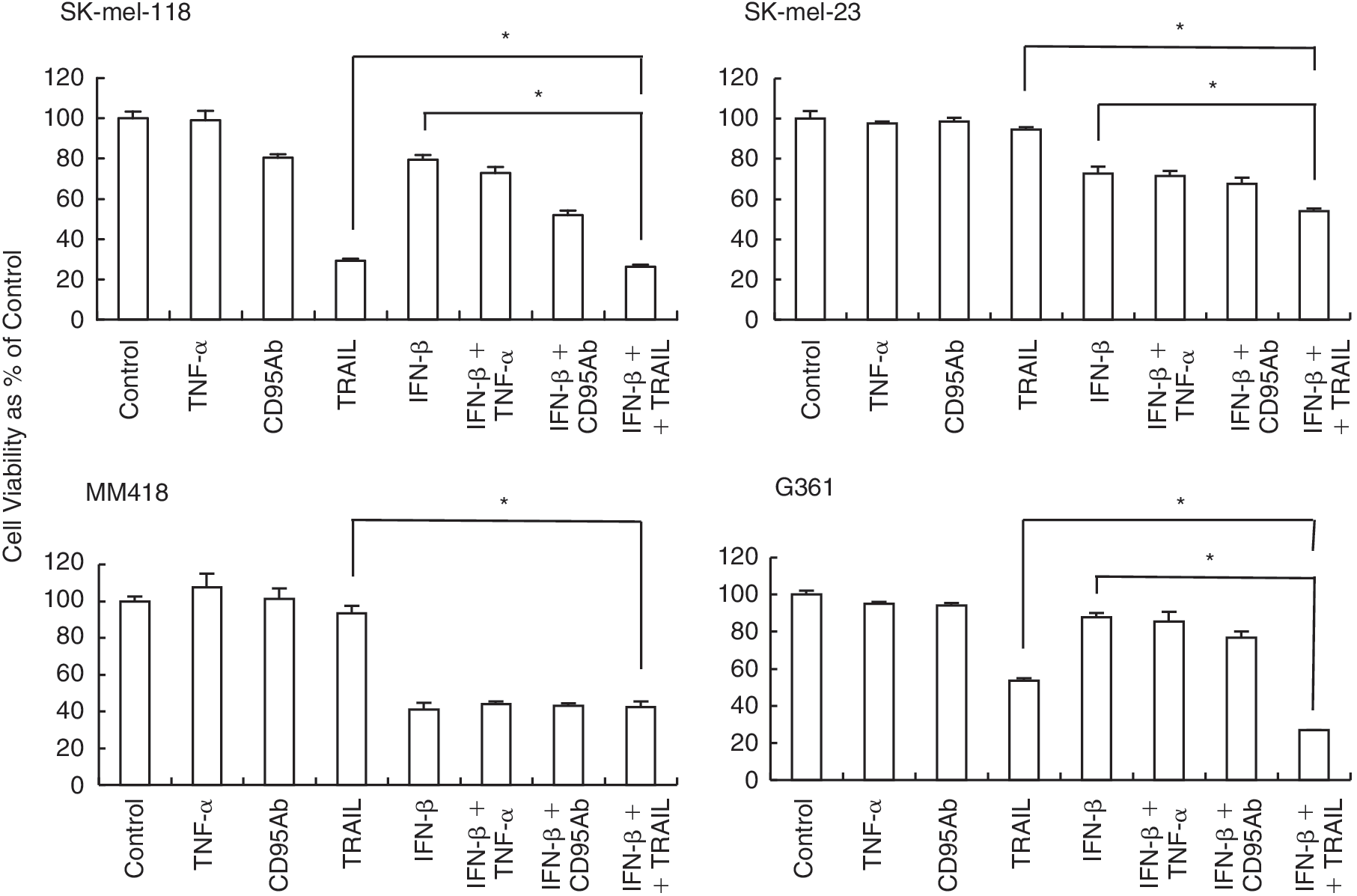
Cell viability assay after treatment with exogenous apoptosis-inducing ligands and interferon (IFN)-β pretreatment followed by apoptosis-inducing ligands. Cells were plated in 96-well plates with 1 × 103 cells/well. Cells were treated with TNF-α, the anti-Fas (CD95) antibody, or tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) for 48 h at the concentrations of 50, 500, and 100 ng/mL, respectively, or with 1,000 IU/mL IFN-β (24 h) pretreatment followed by apoptosis-inducing ligands for 48 h at the same concentrations as above. Cell viability was determined by the formazan-formation assay in triplicate. The results are expressed as relative values (mean ± S.D.) to untreated controls. *P < 0.05.
IFN-β pretreatment enhanced up-regulation of caspase-2 activity in G361 treated with TRAIL.
We next analyzed the up-regulation of caspase activity of G361, which is an IFN-resistant cell line, when cells were treated with TRAIL, and with IFN-β followed by TRAIL. The cells treated with TRAIL showed up-regulation of caspase-2 and −3 activities as did IFN-sensitive cell lines, in which IFN-β induced apoptosis. Furthermore, IFN-β pretreatment enhanced the up-regulation of caspase-2 and −3 (Fig. 7), correlated with inducibility of apoptosis (Fig. 6).
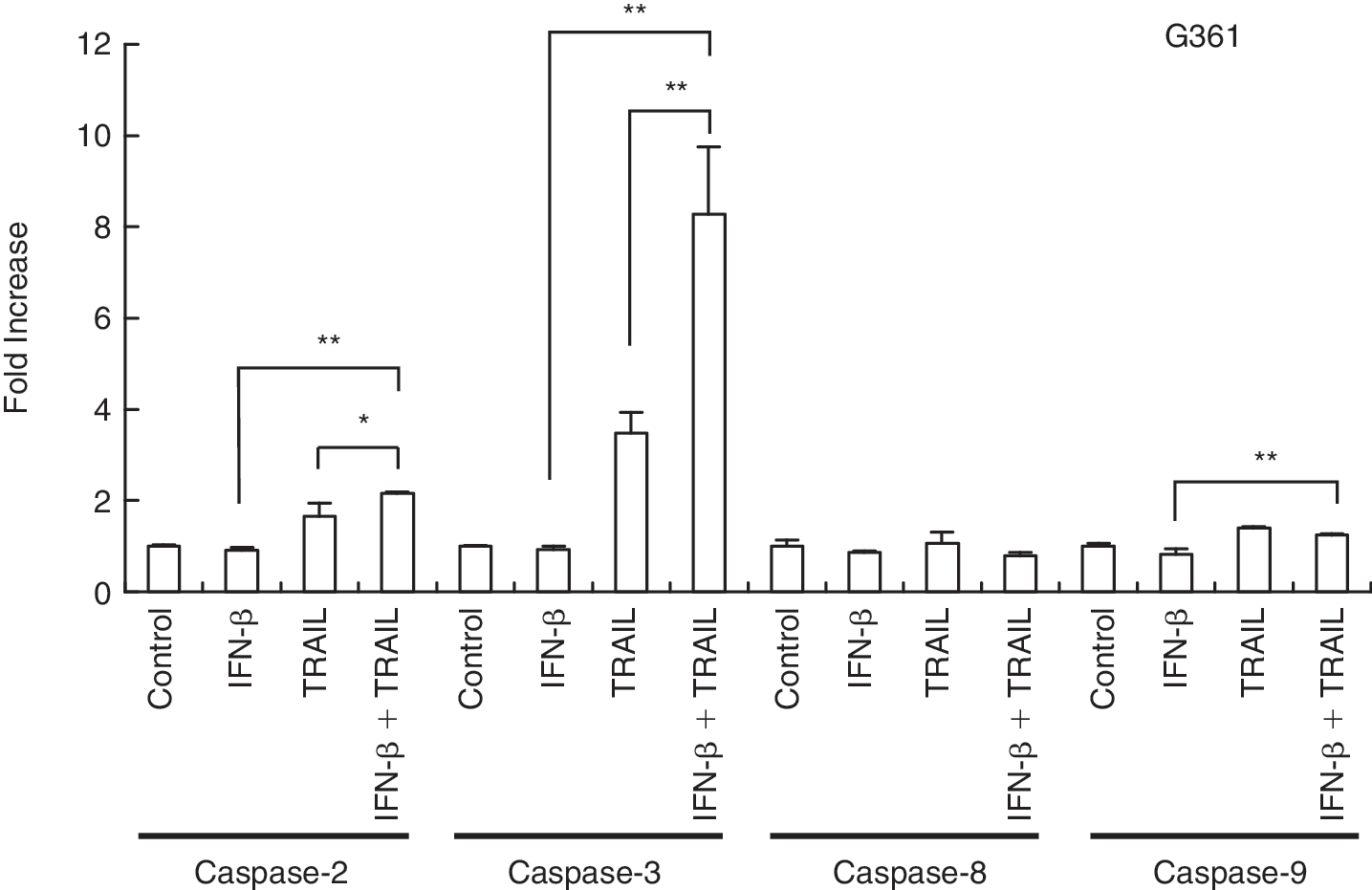
Caspase assay after treatment with apoptosis-inducing ligands and interferon (IFN)-β pretreatment followed by apoptosis-inducing ligands. G361 cells were plated in a 10-cm dishes with 2 × 104 cells/mL. Cells were treated with TRAIL for 24 h at the concentration of 100 ng/mL, or with 1,000 IU/mL IFN-β (24 h) pretreatment followed by a PBS wash and addition of TRAIL for 24 h at the same concentration as above. Cells were harvested and caspase activity was assessed by caspase fluorometric assay as in the manufacturer's protocol. The caspase activity is expressed as the fold increase compared with the untreated cells. TRAIL, tumor necrosis factor-related apoptosis-inducing ligand; TNF-α, tumor necrosis factor α; **P < 0.01, *P < 0.05.
Discussion
In this study we found that cell death was promoted by IFN-α2 and -β treatments in human melanoma cell lines (SK-mel-118, SK-mel-23, and MM418), and that IFN-β preferentially induced cytotoxic effects compared to IFN-α2. We also found that this cell death induced by IFN-β was induced by apoptosis.
It has been reported that IFN-β treatment induces apoptosis that is dependent on TRAIL induction in melanoma cells. It was indicated that the cells that failed to express TRAIL after IFN-β treatment were resistant to IFN-induced apoptosis, whereas those that induced TRAIL by IFN-β were sensitive to IFN-induced apoptosis (Chawla-Sarkar and others 2001). The induction of TRAIL by IFN-β has been shown to initiate the apoptotic cascade in a wide variety of tumor cells (Shin and others 2005; Vogler and others 2007). It was shown in melanoma cells that TRAIL had a more significant tumoricidal effect than other members of the tumor necrosis factor family such as TNF-α and Fas ligand (Griffith and others 1998). We also showed a similar result that TRAIL had more significant apoptotic effect than the other apoptosis-inducing ligands, such as TNF-α and anti-Fas antibody, in TRAIL-sensitive cell lines, SK-mel-118 and G361 (Fig. 6). The induction of TRAIL by IFN-β was shown to be necessary, but still insufficient to induce apoptosis (Chawla-Sarkar and others 2002). In our study 2 melanoma cell lines, SK-mel-118 and SK-mel-23, exhibited such biological reactions in IFN-β-induced apoptosis, and markedly up-regulated TRAIL mRNA in response to IFN-β treatment. In contrast, in G361, in which ISGs were induced by IFN-β treatment, IFN-β did not induce apoptosis and significant expression of TRAIL mRNA, although exogenous TRAIL induced apoptotic cell death. However, anti-TRAIL antibody slightly suppressed IFN-β-induced apoptosis but did not show any significant anti-apoptotic effect by IFN-β on SK-mel-118 and SK-mel-23 cells. These results indicated that a TRAIL-independent pathway may exist, although TRAIL expression contributes to IFN-β-induced apoptosis to some extent.
Our study showed a new and unique finding in the study of MM418. MM418 significantly underwent apoptosis induced by IFN-β treatment but did not exhibit significant expression of TRAIL mRNA (Fig. 1B). Furthermore, exogenous TRAIL did not induce apoptosis in MM418, and caspase-2 activation of MM418 by IFN-β treatment was much higher than in SK-mel-118 and SK-mel-23, which are thought to undergo TRAIL-mediated apoptosis to some extent. These findings suggested that MM418 cells underwent apoptosis via an alternative pathway, such as a TRAIL-independent apoptotic pathway induced by IFN-β.
Several recent studies demonstrated TRAIL-mediated apoptosis through the caspase-2 pathway in other tumor cell lines (Wagner and others 2004; Shin and others 2005). They suggested that caspase-2 was required in the upstream of Bid, which leads to caspase-9 activation in the mitochondrial apoptotic pathway (Wagner and others 2004; Bonzon and others 2006). Samraj and others indicated that caspase-2 activation was absent in a mutant Jurkat T-cell line with a defect of caspase-9 when treated with an anticancer drug releasing cytochrome c (Samraj and others 2007). Shin and others also indicated that caspase-2 processed procaspase-8 (Shin and others 2005). Thus caspase-2 appears to have diverse functions in apoptotic processes. Our study suggested the possible presence of an alternative IFN-induced apoptotic pathway that activates caspase-2 directly without induction of TRAIL. In a previous study, TRAIL was indicated to be a novel therapeutic modality useful for the management of melanomas (Ren and others 2004). We, however, identified TRAIL-independent IFN-β-induced apoptosis in one cell line, MM418.
Another new finding in this study was for G361. It did not show IFN-β-mediated apoptosis, but its IFN signal transduction seemed to be normal. Griffith and others previously reported that only TRAIL among apoptosis-inducing ligands had a significant cytotoxic effect (Griffith and others 1998). In this study TRAIL mRNA was not induced by IFN-β in G361, although other ISGs were expressed. Thus, this loss of TRAIL-mRNA induction may be a meaningful biological process for the resistance to IFN-β-induced apoptosis because exogenous TRAIL induced significant apoptosis in G361 (Fig. 6). It has also been suggested that IFN-β pretreatment sensitizes TRAIL-mediated apoptosis (Chawla-Sarkar and others 2002). In this study, we also showed that IFN-β pretreatment increased sensitivity against TRAIL-mediated apoptosis in G361 and SK-mel-23, SK-mel-118. When G361 was treated with IFN-β followed by additional TRAIL treatment, caspase-2 activity was up-regulated compared with that after TRAIL treatment alone. Thus not only TRAIL but also other factors induced by IFN seem to be important for apoptotic processes, because synergetic activity of TRAIL and IFN-β treatment was observed in this study after treatment with TRAIL and IFN-β in G361 and specifically in SK-mel-23, in which TRAIL alone did not induce apoptosis although IFN-β pretreatment enhanced apoptosis by TRAIL stimulation. Along with a previous report (Chawla-Sarkar and others 2002), we found in this study that the TRAIL/caspase-2 system was important for induction of apoptosis in 3 (SK-mel-118, SK-mel-23, G361) of the 4 melanoma cell lines tested. On the other hand, in MM418 IFN-induced caspase-2 and −3 activation and apoptosis occurred without TRAIL expression.
Our results further suggested that those melanoma cell lines (SK-mel-118, SK-mel-23, MM418), in which apoptosis was induced by IFN-β, commonly had up-regulated caspase-2 activity with (SK-mel-118, SK-mel-23) or without (MM418) involvement of a TRAIL-related pathway. Furthermore, caspase-2 and −3 inhibitors remarkably suppressed IFN-β-induced apoptosis that was equivalent to that in IFN-β-sensitive cell lines. The finding was a common phenomenon in these cell lines, and the increased apoptotic population had up-regulation on caspase-2 activity, but not on TRAIL induction.
Thus our present results clearly indicate that the extent of the IFN-β-induced apoptosis depends on up-regulation of caspase-2 activity more strongly than on induction of TRAIL. These findings suggest that measurement of caspase-2 activity in primary culture cells from excised melanoma tissues can be a novel marker for estimating the extent of the cytotoxic effect of IFN-β adjuvant therapy for melanoma. Caspase-2 activity may be useful for IFN-induced sensitization of chemotherapeutic drugs to melanoma cells.
Footnotes
Acknowledgments
We are indebted to Drs. Akinori Kawakami, Kenji Yanagisawa and Masae Okura of Sapporo Medical University for their valuable contributions. This study was supported by Grants-in Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Culture, Japan (16390319, 16024218) and the Ministry of Health, Labour and Welfare, Japan (H17-nano-004).
Author Disclosure Statement
No competing financial interests exist.