
Abstract
Initiation of an effective adaptive immune response against a foreign pathogen requires orchestrated encounters between lymphocytes and antigen-presenting cells. The tissues of the lymphoid system provide the ideal environment for increasing the efficiency of these encounters. Within the spleen, the mucosal-associated lymphoid tissues, and the lymph nodes, an intricate network of stromal cells, collagen fibers, and extracellular matrix exists that effectively compartmentalizes immune cells as they transit through these tissues. The stromal cells within lymphoid tissues are by no means homogenous, and it is now clear that these cells are not merely sessile bystanders during immune responses. Indeed, stromal cells within lymphoid tissues are the source of important cytokines and chemokines that guide and polarize immune cells. Here, we review the cytokines that maintain the integrity of this important stromal scaffold system within the lymphoid tissue, paying particular attention to the Lymphotoxin pathway, which is an important player in stromal cell biology. How cytokines maintain the organization of lymphoid tissues during development, in the adult animal, during inflammation and during disease will be discussed in sequence, and the clinical implications of targeting cytokines that regulate lymphoid tissue stroma will be considered.
Introduction
Development of lymphoid tissues
Lymphoid tissue development, for the most part, takes place during embryogenesis. The exceptions are Peyer's patches (PPs) that continue to develop during the postnatal period, ILF and nasal-associated lymphoid tissues (NALTs) that develop exclusively during the postnatal period, and inducible lymphoid tissues such as the bronchial-associated lymphoid tissues that develop in the adult animal in response to inflammation (Mebius 2003; Drayton and others 2006). Further, NALT, ILF, and bronchial-associated lymphoid tissue require environmental signals such as commensal bacteria for their establishment/maintenance; indeed, ILF formation can be altered in the absence of innate immune sensors of the commensal flora (Bouskra and others 2008).
In all cases, however, interactions between a few key cell types establish lymphoid tissue ontogeny. A complex program regulates lymphatic vasculature development, and this vasculature underpins the developing SLO anlage (Oliver 2004). Lymphoid tissue initiator cells interact with local mesenchymal cells in a RET/RET-ligand-dependent manner (Adachi and others 1997; Veiga-Fernandes and others 2007), and this is followed by subsequent interactions between 2 key cell types, lymphoid tissue inducer cells (LTi), and lymphoid tissue organizer cells (LTo). These 2 cell types have been intensively studied and lay the foundation for the subsequent development of an SLO, with iterative interactions between these 2 cell types being reinforced by several chemokine/cytokine feedback loops [reviewed by Ruddle and Akirav (2009)]. LTi cells, which are hematopoietically derived, are first attracted to chemokine gradients, particularly CXC-chemokine ligand (CXCL)13, which signal the CXC-chemokine receptor (CXCR)5 counter-receptor on the LTi. The CXCR5/CXCL13 axis is clearly required for SLO development, owing to its role as a chemoattractant for LTi, and later for mature lymphocytes (Forster and others 1996). For some time, it was a mystery as to how CXCL13 secretion was induced in locations of lymphoid tissue development; however, recent data point to retinoic acid as a local stimulator of CXCL13 expression (van de Pavert and others 2009). Once guided to these locations, LTi cells respond to cytokines within the developing tissue. For example, in the location of peripheral LN development, receptor activator of NF-κB–ligand (RANKL) interactions are critical, and this cytokine loop appears to act in both an autocrine and paracrine manner (Dougall and others 1999; Kim and others 2000). In the case of PP development in the gut, interleukin (IL)-7 but not RANKL is important (Adachi and others 1998), although IL-7 likely plays a role in SLO development outside of the gut since common γ-chain-containing cytokine receptors are needed for peripheral LN development as well (Cao and others 1995; Luther and others 2003).
LTi activation induces expression of Lymphotoxin (LT)αβ (Honda and others 2001; Yoshida and others 2002). This cytokine is a member of the tumor necrosis factor (TNF) superfamily of molecules and has diverse functions in both the developing and adult immune system (Gommerman and Browning 2003). In the case of most SLO development, LTαβ cannot be substituted by other TNF family member cytokines, with the exception of mesenteric LNs, which can also be supported by LTβ-receptor (LTβR) signaling by the alternative ligand LIGHT (lymphotoxin-like, exhibits inducible expression and competes with HSV glycoprotein D for HVEM, a receptor expressed by T lymphocytes) (Scheu and others 2002). Further, a reduction/loss of PPs in TNFα-deficient mice has been observed, implying that TNFα is critical for optimal PP generation (Kuprash and others 2005).
Lymphotoxin as a central player in LN development
LTβR-deficient (Futterer and others 1998), LTβ-deficient (Alimzhanov and others 1997; Koni and others 1997), and LTα-deficient (De Togni and others 1994) mice all exhibit profound defects in LN and PP development, although NALT development appears to proceed, albeit suboptimally, in the postnatal phase (Harmsen and others 2002; Ying and others 2005). The timing of appearance of various lymphoid tissues can be determined by in utero injection of an antagonist to the LTβR pathway, LTβR-immunoglobulin (Ig); indeed, agonists to this pathway can restore lymphoid tissue development in LTβ-deficient mice (Rennert and others 1998). Once LTαβ is expressed on LTi cells during development (Honda and others 2001), many important signaling events take place via signaling of the LTαβ counter-receptor, LTβR. At sites of SLO development, LTo cells express both intercellular adhesion molecule-1 (ICAM-1) and vascular cell adhesion molecule 1 (VCAM-1) adhesion molecules; however, increased expression of these molecules requires additional LTβR-signaling (Benezech and others 2010). LTβR signaling also induces the further secretion of chemokines such as CXCL13 that attract more lymphoid cells into the developing lymphoid tissue. The signaling of the CXCL13 counter-receptor (CXCR5) on LTi further induces stable expression of LTαβ (Ansel and others 2000). In this manner, a positive feedback loop is established, with LTαβ as the central cytokine critical for its propagation. The relevant downstream signaling via the LTβR for lymphoid tissue development is through the NF-κB pathway. In particular, aly/aly mice that harbor a mutation in the NF-kB-inducing kinase gene responsible for initiating the alternative NF-κB pathway (NF-κB2) largely phenocopy LTβR−/− mice in terms of their deficit in organized lymphoid tissue (Shinkura and others 1999). Therefore, a central role for LTβR signaling in lymphoid tissue development has been established, with additional cooperation from other cytokines (Fig. 1A).
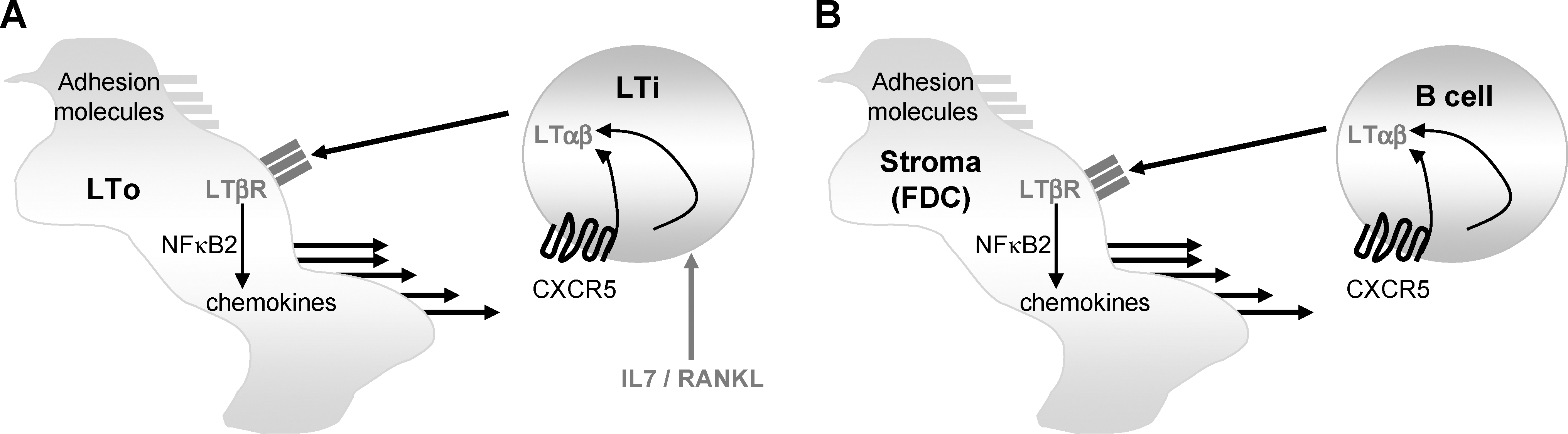
Iterative interactions between LTαβ-expressing hematopoietic cells and LTβR-expressing stroma initiate and maintain lymphoid tissue.
Organization of Lymphoid Tissues: A Central Role for Stroma/Cytokine Interactions
By studying the developmental steps that lead to lymphoid tissue establishment during embryogenesis, much can be extrapolated in terms of how lymphoid tissues are maintained in the adult animal (Fig. 1B). Indeed, the same LT-driven feedback loop that is required to establish lymphoid tissues during development is recapitulated in the spleen, LNs, and PPs as will be discussed below. Some aspects of SLO organization are shared by all lymphoid tissues. One hallmark of the SLOs is the segregation of B and T lymphocytes into discrete zones. Briefly, these zones are surrounded by a capsule in the case of the LN, and a marginal zone (MZ) in the case of the spleen. In the spleen, the combination of the B/T cell zones and the MZ make up a white pulp nodule that is surrounded by the red pulp, which is rich in macrophages. Lymphocytes enter the spleen through a central arteriole, and in the case of the LN, postcapillary venules called high endothelial venules (HEVs) represent the port of entry for naïve lymphocytes. The general architecture of LNs, spleen, and PPs is depicted in Fig. 2, and in this section we describe how cytokines modulate the organization of SLOs under homeostatic conditions in naïve animals.
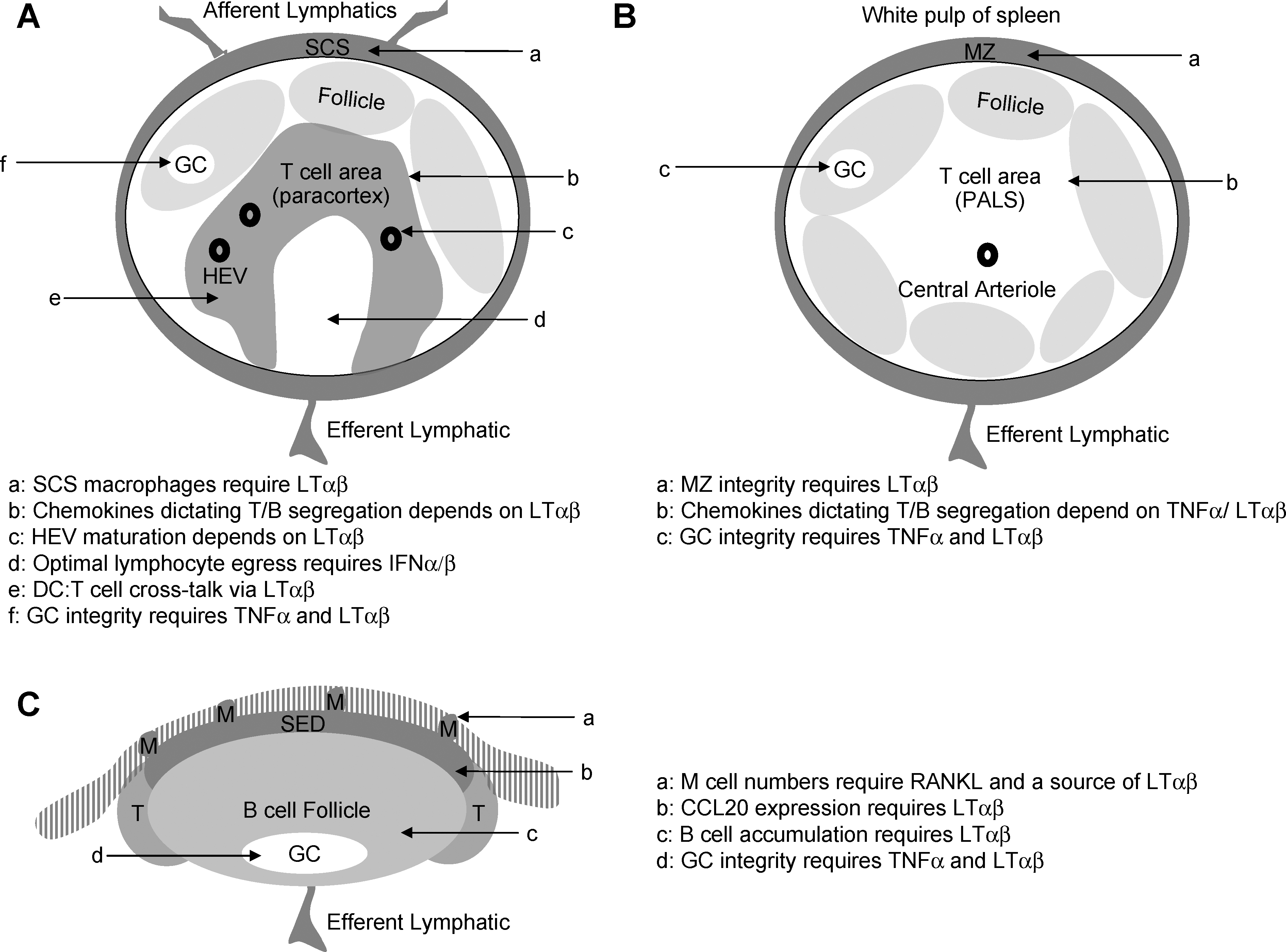
Homeostatic cytokines maintain the integrity of multiple lymphoid tissues. Cytokines such as LTαβ and TNFα as well as RANKL are critical for maintaining the integrity of lymphoid tissues such as lymph nodes
Spatial segregation of B and T lymphocytes in the lymphoid tissues
Constitutive LTβR signaling is required to maintain CXCL13 production in the follicles of SLOs (Ansel and others 2000). These follicles are largely populated by B cells, and can house ongoing germinal center reactions, sites of intense B cell activation. As one moves deeper into the lymphoid tissues, B cells become markedly sparser and T cell regions are established. In the spleen, this segregation between B and T cell zones is LT dependent, and in the case of the T cell-rich periarteriolar lymphoid sheath of the spleen, this is due to the LTβR-dependent production of CC-chemokine ligand (CCL)19/CCL21, which draws CC-chemokine receptor (CCR)7-expressing T cells into this compartment, with some overlapping albeit secondary function of TNFα in this process (Ngo and others 1999). Splenic organization appears to be largely dependent on LTαβ expression by B cells (Tumanov and others 2002). The LTβR-dependent effect on chemokine production is also observed in the PP where B cell accumulation in this tissue is strongly compromised in mice reconstituted with LTβ−/− B cells (Tumanov and others 2004). Thus, in the adult animal, the LTαβ cytokine and, to a lesser extent, TNFα are required to actively maintain lymphocyte compartmentalization. Indeed, as evidence of the homeostatic dependence on LTβR signaling in the maintenance of lymphoid tissue architecture, a single injection of LTβR-Ig results in the rapid dissolution of the B/T boundary in the SLOs (Mackay and others 1997).
Entry of lymphocytes into the lymphoid tissues
Whereas blood lymphocytes enter the spleen through a central arteriol, in the case of LNs, naïve lymphocytes enter the T-cell-rich paracortex region from the blood through the HEV. Peripheral LNs contain HEV that display peripheral LN addressin, whereas mucosal LNs have HEV that express mucosal vascular addressin cell adhesion molecule 1 (MAdCAM-1). HEV arise during LN development as well as during LN neogenesis in nonlymphoid sites. In the adult animal, the maintenance of HEV is more complex than was original supposed with HEV under a constant maturation program whereby the addressins they express such as peripheral LN addressin in the peripheral LNs require post-translational modification via sulfyltransferases. The expression levels of these enzymes are likewise under LT-control (Drayton and others 2003; Browning and others 2005).
Entry of Ag into the lymphoid tissues
Surrounding the follicle is the MZ of the spleen, or in the case of the LNs, the sub-capsular sinus (SCS). In the PP, the sub-epithelium dome (SED) overlays the B cell follicle. In all of these cases (MZ, SCS, and SED), there is a role for TNF family cytokines signaling in maintaining the integrity of these compartments that are rich in Ag-presenting cells such as macrophages and dendritic cells (DCs), and are also the primary route of entry for pathogens. For example, the splenic MZ of LT-deficient mice lacks expression of MAdCAM-1 on the MZ sinus and is devoid of MZ-B cells, MZ-macrophages, and metallophilic macrophages (Mackay and others 1997; Drayton and others 2003; Browning and others 2005). The absence of the MZ in the splenic lymphoid tissue compromises the immune response to systemic viral infection (Junt and others 2006). SCS macrophages may represent the LN equivalent of these cells; indeed, SCS macrophages are absent in LT-deficient mice. This has a profound affect on the transport of immune complexes in that they fail to reach the B cell compartment and humoral immunity is compromised (Phan and others 2009). Ag entry into the PP is through microfold (M) cells in the overlaying epithelium, and although these do not require LTαβ expression on B cells for their maintenance (Tumanov and others 2004), they are reduced in LTβR-Ig-treated mice (Debard and others 2001), suggesting that the relevant source of LTαβ in this situation is a non-B cell such as a gut-resident LTi-like cell, which persist in adult mice, in particular in the gut. This is consistent with recombination-activating gene (RAG)−/− mice having normal M cell numbers (Debard and others 2001). It is also clear that RANKL plays an important role in maintaining M cell numbers, with the outcome of reduced germinal center (GC) and IgA responses in this tissue (Knoop and others 2009). What remains to be further explored is the influence of cytokines on the composition of DC in the SED, as DC can perform Ag-capture functions from the gut lumen. Since the LT cytokine is required for CCL20 expression in this region, it is not unreasonable to suspect that the accumulation of CCR6+ DC may be compromised in the PP in LT-deficient mice (Rumbo and others 2004). Thus, TNF family cytokines control critical aspects of the microenvironments within lymphoid tissues that control pathogen entry.
Cytokines and stromal cell diversity/function
Reticular networks have been observed in SLOs for more than 100 years (Glegg and others 1953). It was initially assumed that the nonhematopoietic cells in SLOs exert mainly a structural function, facilitating Ag and Ab transport and assisting in the expansion and contraction of LN and the spleen. However, recent observations demonstrated that the transport of cytokines from the afferent lymph to HEV involves conduits formed by nonhematopoietic cells, which lead to a new appreciation of the functions of the lymphoid stroma (Anderson and Shaw 1993; Gretz and others 2000; Nolte and others 2003). During the last decade it has become increasingly clear that within lymphoid tissues, the migration, survival, and function of lymphocytes is intimately connected with the underlying stromal cell population. While difficult to study due to their rarity and their collagen-rich environment, identification of some subsets of stromal cells has been possible (Mueller and Germain 2009). On the basis of their phenotype, location, and function, nonhematopoietic stromal cell types in SLOs can be divided into at least 7 subsets (Table 1). We focus here on the stromal cell subsets that make up the parenchyma of SLOs, such as follicular DCs (FDCs), fibroblastic reticular cells (FRCs), marginal reticular cells (MRCs), splenic red pulp fibroblasts (SRPFs), and LN medullary fibroblasts. For details on blood endothelial cells (BEC) and lymphatic endothelial cells (LECs) that are involved in blood and lymph flow through SLOs, we refer to recently published reviews (von Andrian and Mempel 2003; Cueni and Detmar 2008; Jurisic and Detmar 2009).
BAFF, B cell activating factor belonging to the TNF family; CD, cluster of differentiation; CCL, CC-chemokine ligand; CR, complement receptor; CXCL, CXC-chemokine ligand; ER-TR, Erasmus University Rotterdam-Thymic Reticulum; ESAM, endothelial cell-specific adhesion molecule; Fc, fragment, crystallizable; FDC-M; follicular dendritic cell marker; gp, glycoprotein; HGF, hepatocyte growth factor; ICAM, intercellular adhesion molecule; IL, interleukin; JAM, junctional adhesion molecule; LN, lymph node; LTβR, lymphotoxin-beta receptor; LYVE, lymphatic vessel endothelial hyaluronan receptor; MAdCAM, mucosal vascular addressin cell adhesion molecule; MALT, mucosa-associated lymphoid tissue; MFG-E, milk fat globule epidermal growth factor; MHC, major histocompatibility complex; MZ, marginal zone; PDGFR, platelet-derived growth factor receptor; PNaD, peripheral node addressin; PP, Peyer's patch; PROX, Prospero homeobox protein; RANKL, receptor activator of NF-κB ligand; SCS, sub-capsular sinus; SMA, smooth muscle actin; SPL, spleen; TLO, tertiary lymphoid organ; TNFR, tumor necrosis factor receptor; VEGFR, vascular endothelial growth factor receptor; VCAM, vascular cell adhesion molecule; Zo, zonula occludens protein.
Follicular DCs
FDCs are characterized by expression of markers such as FDC-M1 (milk fat globule epidermal growth factor 8), FDC-M2 (complement C4), and cluster of differentiation (CD)157 (BP-3) and form a dense network in the center of B cell follicles in the spleen, LN, and other lymphoid tissues, including PPs, the MALT, and tertiary lymphoid organs (TLOs) (Tew and others 1997). They also express high levels of VCAM-1 and MAdCAM-1, as well as molecules that are common to all subsets of stromal cells in the lymphoid tissues, such as desmin and laminin (Balogh and others 2002). Expression of fragment, crystallizable receptors (FCRs) (such as CD16, CD23, and CD32), and complement receptors (such as CD21 and CD35) enables FDCs to capture and present unprocessed Ag, particularly in the form of immune complexes (Roozendaal and Carroll 2007). FDCs provide a scaffold for B cells, and in particular during immune responses, FDCs can aggregate to support GC-B cells (Tew and others 1990). Again, LTβR signaling on this type of stromal cell is critically required for the maintenance of FDCs as evidenced by their rapid disappearance upon LTβR-Ig treatment in mice (Mackay and Browning 1998) and primates (Gommerman and others 2002). Moreover, FDCs constitute the main source of CXCL13, produced in an LT-dependent manner, which is key to attract CXCR5+ B cells into follicles in the steady state. During an immune response, interactions between FDCs, B cells, and T follicular helper cell (Tfh) occur in the GC in a highly dynamic manner. While in the light zone of GCs, B cells interact with FDCs expressing CXCL13, interactions with CXCL12-expressing stromal cells occur in the dark zone. In addition, FDCs are now thought to secrete cytokines and growth factors that support GC-B cell survival within this niche such as IL-15 (Park and others 2004) and the TNF family member B cell-activating factor belonging to the TNF family (BAFF) (Hase and others 2004).
Fibroblastic reticular cells
The FRCs and FDCs form a continuous network that support leukocyte migration, lymphoid tissue homeostasis, and the control of immune responses by virtue of their ability to guide lymphocyte encounters (Bajenoff and others 2006). In the T cell zones of SLOs, FRCs surround the central arterioles and HEVs. FRCs express extracellular matrix components such as fibrillin, laminin, and fibronectin and intracellular molecules that are found in some fibroblasts, including desmin, α-smooth muscle actin, and transglutaminase, which are involved in the formation, expansion, and contraction of these collagen-rich reticular fibers (Van Vliet and others 1986; Farr and others 1992; Balogh and others 2004; Katakai and others 2004; Sixt and others 2005; Link and others 2007). Like many fibroblasts in different tissues and other stromal cell types in SLOs (Table 1), FRCs express integrin subunits such as ICAM-1 and VCAM-1 as well as Erasmus University Rotterdam-Thymic Reticulum 7 (ERTR-7) (Van Vliet and others 1986), but can be differentiated from other nonendothelial lymphoid stromal cell types by their expression of podoplanin, which is also known as glycoprotein 38 (Farr and others 1992). One function of the FRC network in the T cell zones of SLOs is to allow delivery of distinct signals through the conduit networks through size exclusion. While small molecules, such as chemokines and Ags, can enter the FRC network from the lymph in LNs and are delivered to T cell zones and HEVs, high-molecular-mass molecules cannot enter and are trapped by SCS macrophages that accumulate in the cortical sinuses or drain through efferent lymphatic vessels (Gretz and others 2000). In contrast, in the spleen, FRCs allow the entry of large blood-borne molecules in conduit structures of the T cell zone (Nolte and others 2003).
Lymphocytes within the FRC network of T cell zones have been shown to be in constant dynamic motion moving along strands formed by FRCs that function as guidance paths for cell migration, transporting lymph through the tissue for sampling by DC (Bajenoff and others 2006). Another critical function of FRCs is their production of the chemokines CCL19 and CCL21, which are key for delineating the T cell zones and through the chemoattraction of T cell and DCs that express CCR7 (Gretz and others 1997; Gunn and others 1998; Luther and others 2000b). Further, FRCs produce IL-7 that promotes the homeostasis of T cells in naïve mice (Link and others 2007). The full function of FRCs, at least in vitro, depends on expression of LTαβ by lymphocytes presumably due to LTβR:LTαβ cross-talk between FRCs and the lymphocytes themselves (Katakai and others 2004).
Marginal reticular cells
A layer of nonhematopoietic cells localized underneath the SCS in LNs, within the dome of the PP and within the MZ of the spleen has been characterized as MRCs since these stromal cells have a phenotype that is distinct from that found in B and T cell zones (Katakai and others 2008). Although MRCs express many markers in common with other subsets of stromal cells (Balogh and others 2004), they seem to uniquely express the TNF family member RANKL (tumor necrosis factor-related activation-induced cytokine), suggesting that these cells might be integral organizers of SLO structure during organogenesis (Katakai and others 2008). Importantly, many of the MRCs are localized at the edges of the B cell follicles, and similarly to the centrally located FDCs, MRCs express CXCL13 and function as a conduit network delivering Ag from the SCS/MZ to the B cell follicles (Junt and others 2007; Roozendaal and others 2009), suggesting that MRCs are a specialized subset of lymphoid stromal cells with important roles in the homeostasis of B cells and capture and delivery of Ag (Katakai and others 2008).
Splenic red pulp fibroblasts
Cords in the red pulp of the spleen are composed of a compact network of fibroblasts and reticular fibers, have a crucial role in filtering blood, and contribute to immune defense (Weiss 1991; Ellyard and others 2005). SRPF aid in the localization of different cells within the red pulp through expression of several integrin chains (van den Berg and others 1993). Moreover, red pulp fibroblasts express CXCL12, leading to the localization of short-lived CXCR4+LFA-1+ plasma cells (PCs) to the splenic red pulp where they secrete large amounts of Ig into the circulation (Hargreaves and others 2001; Ellyard and others 2005). Upon infection, stimulation by pro-inflammatory mediators such as IL-1 or microbe-associated molecular patterns such as lipopolysaccharide provokes SRPFs to produce cytokines such as IL-6 and to become activated and fused, forming the so-called barrier cells (Weiss 1991). This activation process alters and restricts blood flow through the spleen during times of stress. These changes of SRPFs in the marginal sinuses around the white pulp therefore control cell entry to or exit from the spleen during infection and inflammation.
LN medullary fibroblasts
In the medullary cords and sinuses of LNs, fibroblasts and reticular fibers form a cellular network wherein many mast cells, macrophages, and DC, as well as lymphocytes, including PC, can be found (Ushiki and others 1995; Katakai and others 2004; Willard-Mack 2006). Although no specialized stromal cell marker and function has been clearly assigned yet, LN medullary fibroblast have been shown to express CXCL12 in the steady state (Hargreaves and others 2001), suggesting that they might have similar roles to SRPFs in directing PC localization in LNs.
Taken together, most aspects of lymphoid tissue architecture are regulated by cytokines, and stromal elements within the SLOs also actively secrete cytokines/chemokines. In a naïve animal, these structural aspects of the SLOs optimize the entry of lymphocytes and their capacity to interact with the Ag-bearing Ag-presenting cell. In the next section, we explain how inflammation and infection changes the lymphoid tissue architecture so that cell–cell interactions in the SLOs are enhanced, thus promoting protracted contacts between immune cells.
SLOs During Immune Responses: Influence of Stroma/Cytokine Interactions
On a tissue-level, there are dramatic changes taking place within lymphoid tissues, which actively remodel the anatomy of SLOs (Junt and others 2008; Mueller and Ahmed 2008). This has a tremendous impact on the likelihood of encounters between Ag-specific immune cells and the Ag itself. Cytokines play a key role in regulating these changes in the lymphoid tissues, and without these cytokine-mediated changes, immune responses may proceed inefficiently. In this section we review the gross changes that occur within SLOs upon infection and how this can temporarily change the anatomy and function of these organs.
SLOs remodeling and vascularization
When a robust immune response develops, the cellularity of SLOs expands, increasing the pool of naïve lymphocytes for increased sampling of foreign Ag via regulation of the vascular input (Soderberg and others 2005). This growth is accompanied by increased proliferation of LEC and BEC, leading to angiogenesis of the lymph and blood vessels through increased expression of pro-angiogenic factors such as vascular endothelial growth factors (VEGFs) (Angeli and others 2006; Webster and others 2006). The vascular growth and increased expression of VEGF requires the presence of DC (Webster and others 2006), and the increase of DC migration upon inflammation is reduced by blocking VEGF receptors (Angeli and others 2006), demonstrating a close functional relationship between DC localization and production of pro-angiogenic factors upon inflammation. Importantly, the increase in DC and the enhanced expression of VEGF are markedly reduced in B-cell-deficient animals (Angeli and others 2006). Although the source as well as the mechanisms leading to the production of pro-angiogenic factors in SLOs are not well understood, FRCs have been shown to produce VEGF in an LT-dependent manner and constitute the principal VEGF-expressing cells in homeostatic conditions (Chyou and others 2008). Several other cell types such as monocytes, macrophages, T cells, and B cells have been shown to produce VEGF upon inflammation (Angeli and others 2006; Halin and Detmar 2008). Although cytokines such as TNFα, transforming growth factor-β, IL-1, IL-6, IL-8, and IL-18 have been suggested to exert direct effects (Halin and Detmar 2008), their individual roles in angiogenesis of blood and lymph vessels in SLOs remain largely elusive. The proliferative vascular expansion is then followed by a period of reduced endothelial cell proliferation and maintenance of the expanded endothelial cell number (Webster and others 2006), a process that once again requires the presence of DC (Tzeng and others 2010). Vice-versa, the inflamed lymphatic endothelium suppresses DC maturation and function (Podgrabinska and others 2009), demonstrating that an intimate cross-talk between vascular endothelial cells and DC is required for the successful priming of adaptive immune responses.
Stromal cell collapse and recovery during infection
Stromal cells in SLOs provide the structural and functional basis for the induction and regulation of immune responses, and the integrity of the lymphoid microenvironments determine the immunocompetence of the host (Junt and others 2008; Mueller and Germain 2009). Upon inflammation and infection, similar to LEC and BEC, stromal cell populations such as FRCs and FDCs in SLOs expand (Katakai and others 2004; Peduto and others 2009). However, the structural integrity of SLOs is often challenged during infection by the direct cytopathic effect of microorganisms or by immunopathological processes during acute and chronic immune responses. Lymphoid stromal cells, in particular FRCs and FDCs, are targeted by several intracellular pathogens such as viruses [lymphocytic choriomengitis virus, mouse cytomegalovirus (MCMV), simian immunodeficiency virus, Ebola, Marburg and Lassa viruses] (Mercer and others 1988; Pomeroy and others 1991; Davis and others 1997; Choi and others 2003; Benedict and others 2006; Mueller and others 2007a, 2007b; Scandella and others 2008; Steele and others 2009), parasites (Leishmania major, Leishmania donovani, and Plasmodium yoelii) (Weiss and others 1986; Weiss 1989; Smelt and others 1997; Bogdan and others 2000; Ato and others 2002), bacteria (Salmonella typhimurium) (St. John and Abraham 2009), infectious particles (eg, prions) (Heikenwalder and others 2008), and microbe-associated molecular patterns [eg, the Toll-like receptor 9 (TLR9) agonist CpG] (Heikenwalder and others 2004). Stromal cell targeting by pathogens leads to a marked disorganization of conduit structure and function, resulting in increased cell death, reduced expression of homeostatic chemokines, contributing to pathogen persistence and latency, systemic immune suppression, as well as a transient loss of immunocompetence against secondary infections. The destructive processes are in part driven by the cytokine storm accompanying acute infections, as the severe reduction of homeostatic chemokines in viral and bacterial infections have been attributed mainly to interferon (IFN)-γ (Heikenwalder and others 2004; Mueller and others 2007a). In contrast, TNFα and IL-10 have been shown to be key components in altering the function of stromal cells in a chronic model of infection with the parasite Leishman donovanii (Ato and others 2002). Moreover, stromal cell destruction is mediated exclusively by the action of virus-specific cytotoxic T lymphocytes upon infection with lymphocytic choriomengitis virus, as depletion of CD8+ T cells abrogated the downregulation of homeostatic chemokines and preserved the FRC network (Scandella and others 2008). Thus, identifying the effects of changes in stromal cell physiology are key for our understanding of antimicrobial immunity.
To cope with the cytopathic effects of pathogens, counter-regulatory mechanisms are used to maintain the integrity of the SLO microanatomy and secure immunocompetence, as exemplified by an LT-IFN-β axis regulating lymphocyte survival during MCMV infection (Banks and others 2005; Schneider and others 2008). Deficiency of LTα, LTβ, and LTβR leads to increased susceptibility to infection with MCMV, accompanied by a severe induction of T and B cell apoptosis (Banks and others 2005). LT expression by B cells and LTβR expression by stromal cells were observed to be critical for the initial part of a biphasic type I IFN response, which occurs in the absence of TLR signaling. Interestingly, stromal cells were shown to be the main producer of the early type I IFN response (Schneider and others 2008). Further studies will clarify if this functional axis plays a critical role in other infection models.
Lymphocyte retention within lymphoid tissues
The aforementioned cytokine networks are critical for attracting lymphocytes to the draining lymphoid tissue sampling the relevant infection. However, cytokines such as IFN-α/β can also influence the transit time of lymphocytes within the lymphoid tissue itself by virtue of a CD69-dependent modulation of sphingosine-1-phosphate receptors (Shiow and others 2006). In the case of influenza infection in the lung, it is the local draining mediastinal LNs that is responsive to IFN-α/β-mediated lymphocyte retention, and the same mechanism appears to be at play for plasmacytoid DC as well, providing an additional source of local IFN-α/β (Gao and others 2009). As mentioned, the LT pathway has been shown to be important for optimal Type I IFN responses derived from the nonlymphoid stromal compartment of lymphoid tissues. It remains to be determined if there is a role for the LT pathway in type I IFN release from plasmacytoid DC themselves.
Inducible Niches for Immune Cell Encounters: Regulation by Stroma/Cytokines
As mentioned, cytokines have a dramatic effect on remodeling of lymphoid tissues during infection by affecting the entry of lymphocytes, the de novo vascularization of the SLOs, stromal cell stability within the SLOs, and the retention of lymphocytes within the SLOs. In this section we briefly review examples of transient niches that form both within lymphoid tissues and in nonlymphoid inflamed tissue.
The Germinal Center as the prototypical inflammatory niche
The GC is one of the most intensively studied dynamic niche that persists for the duration of an infection. Significant changes occur within this inducible compartment that are designed to optimize the B cell response to Ag (O'Sullivan and Thomas 2003). As T cells are activated in the T cell areas of the LNs and spleen by Ag-bearing DC, these T cells upregulate inducible costimulator (ICOS) and migrate to B cell follicles by virtue of their induced expression of CXCR5, where they adopt a follicular-helper cell phenotype (Tfh) (Breitfeld and others 2000; Schaerli and others 2000). As the GC niche develops within the follicle, GC-resident FDCs produce CXCL13 to attract additional Tfh, which secrete cytokines optimal for B cell help such as IL-4 and IL-21 (Nurieva and others 2008; Vogelzang and others 2008). Further, T cells that have been activated via ICOS can induce expression of CD40-ligand. Ag-specific B cells that receive cognate T cell help in the form of cytokines/CD40-ligand upregulate LTαβ, which can then induce LTβR signaling in the resident FDCs (Vu and others 2008). This type of cytokine cross-talk (Fig. 3) likely encourages FDCs to aggregate and form a niche that is ideally suited for Ag capture via Fc and complement receptors. Thus, the GC is an excellent example of how cytokines drive an inducible niche within the lymphoid tissues to support an ongoing immune response.
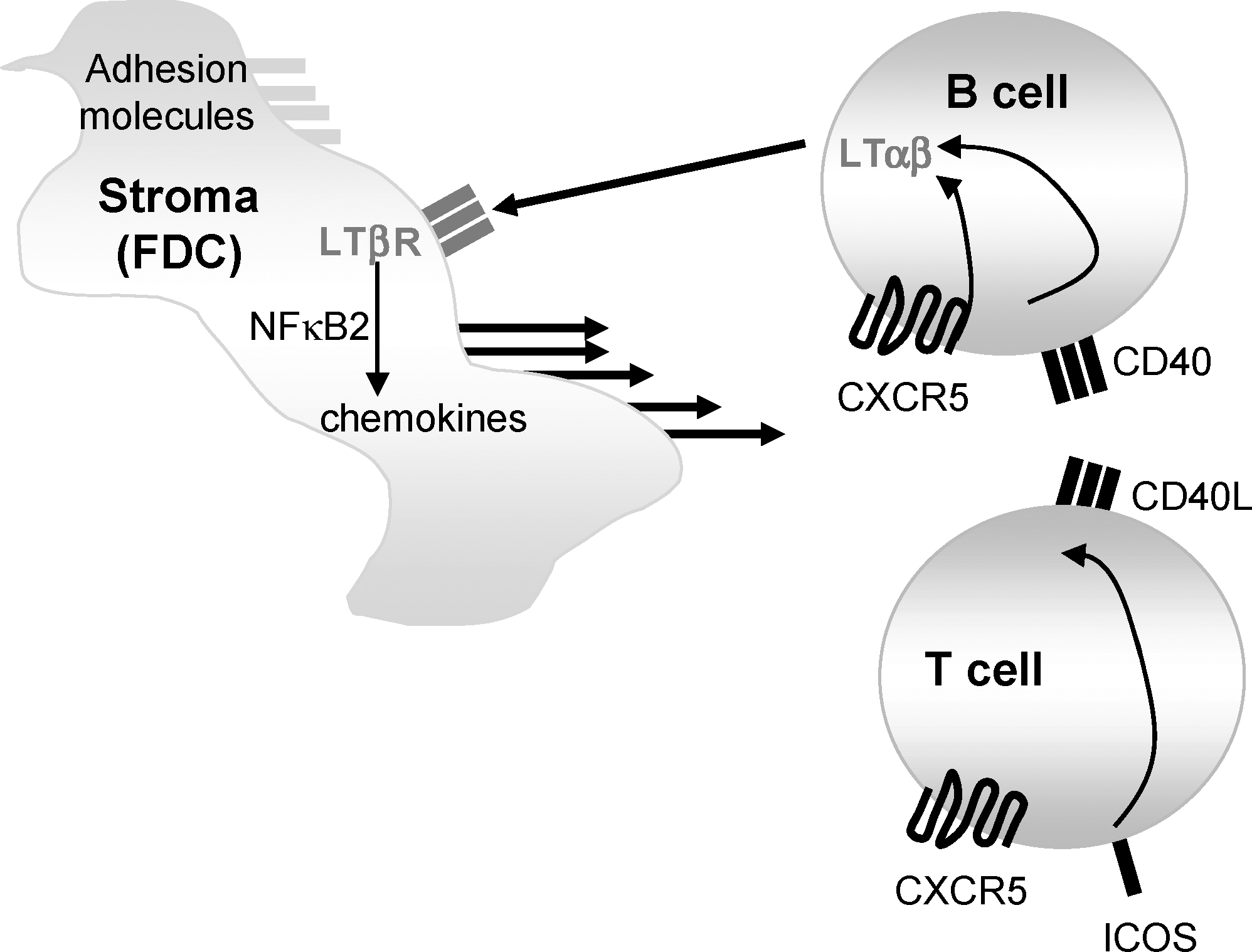
Multiple cytokine interactions propagate germinal center reactions. The convergence of Ag-specific B cells and Ag-specific helper T cells within the B cell follicle results in an interchange of cytokines. Expression of CD40-ligand on T cells is triggered by ICOS ligation and is required to stimulate Ag-specific B cells to up-regulate LTαβ to greater levels than what is normally observed in the nonimmune primary follicle. This expression of LTαβ on GC-B cells drives the formation of tight FDC networks that form a scaffold that supports the formation of the GC niche. The GC niche is transient. Ag, antigen; CD, cluster of differentiation; CXCR, CXC-chemokine receptor; ICOS, inducible costimulator.
ILFs in the gut
While microbes are not required to establish and maintain the general organization of the spleen, LNs, and PPs (Eberl 2005; Tsuji and others 2008a), in contrast, ILFs form after birth, when the intestine is colonized by commensals or upon intestinal inflammation. ILFs are localized between the crypts of the antimesenteric border, predominantly in the most distal part of the small intestine (the ileum) and the colon, and are rich in B cells and DCs, but contain only small numbers of T cells. Although they structurally resemble B cell follicles in LNs and PPs (Eberl 2005; Tsuji and others 2008a), ILFs are inducible and reversible lymphoid tissue aggregates, generating T cell-independent IgA-mediated Ag-specific immune responses to commensal bacteria as well as intestinal pathogens (Lorenz and Newberry 2004; Bouskra and others 2008; Tsuji and others 2008b). The formation of ILFs starts during the second and third week after birth. Clusters containing LTi that are surrounded by a thin-layer of DC start to form. This initial lymphoid structure is often referred to as a cryptopatch (CP), and may develop independently of the gut flora (Kanamori and others 1996). In response to a growing intestinal microflora CP then develop further into ILFs. Similar to fetal LTi, intestinal LTi express IL-7Rα, which is essential for the formation of CP and ILFs (Kanamori and others 1996; Hamada and others 2002). Signaling through IL-7Rα induced by IL-7 and possibly other ligands released by the intestinal epithelium and stromal cells are believed to stimulate expression of LTβR ligands (Eberl 2005). The LT-pathway plays an essential role in the formation of these intestinal lymphoid structures, since LTαβ- and LTβR-deficient animals lack intestinal lymphoid aggregates such as CP and ILFs (Pabst and others 2005). The formation of ILFs is altered in animals deficient for TLRs 2/4, its downstream adaptor proteins myeloid differentiation protein 88, and TIR-domain-containing adapter-inducing interferon-β, as well as in animals lacking the nucleotide-binding oligomerization domain protein (Nod)-like receptors Nod1 and Nod2, demonstrating that innate immune recognition of the intestinal microflora by TLR and Nod-like receptor contributes to the formation and function of ILFs (Bouskra and others 2008). Because these responses eliminate/control their own source of stimulation, ILFs self-regulate their number and size, with an increase in the number of bacteria present in the intestinal lumen inducing an increase in the number and size of ILFs and vice-versa. Thus, aggregated lymphoid structures vary in size and cellular composition, with significant variations depending on species, age, mouse strain, and housing conditions (Pabst and others 2005). Thus, CP and ILFs are dynamic structures of intestinal lymphoid aggregations with a great deal of inborn plasticity.
Tertiary lymphoid structures in the periphery
The same cytokine interactions that influence lymphoid tissue development can be recapitulated during inflammation in nonlymphoid tissues, resulting in the generation of ectopic lymphoid-like structures termed TLOs, which are very similar to the gut-located ILFs. These have been found in inflamed organs during chronic disease such as in the brain during Multiple Sclerosis, in the joint in Rheumatoid Arthritis, in the salivary glands in Sjögren's syndrome, and in infectious disease settings such as hepatitis C, Helicobater pylori–induced gastritis, and chronic Lyme disease (Aloisi and Pujol-Borrell 2006). Although the connection between these ectopic follicles and disease pathogenesis remains under investigation, it is generally agreed that the abnormal influx of immune cells provides a source of cytokines that drives their formation. This includes LTαβ expression on invading lymphocytes, particularly B cells, which can then induce the local production of chemokines by resident stromal cells, with the proof of concept coming from LT-expressing transgenic animals (Luther and others 2000a; Drayton and others 2003). In contrast to the stromal cells in the lymphoid tissues, the stromal cells in these inflammatory foci remain ill-defined.
Conclusions and Perspectives
The role of cytokines in regulating lymphoid tissue has been reviewed here at multiple levels: during SLO development, in maintaining the integrity of SLO microarchitecture in the adult, during the immune response, and during chronic inflammation. Clearly, the regulation of lymphoid tissue is a central aspect to immunity and autoimmunity. As such, it is reasonable to contemplate targeting the cytokines that regulate lymphoid tissue function/integrity as a means of quieting autoimmune disease or for enhancing immunity to tumors. Caution should be observed, however, in that stromal cell populations in the SLOs can have negative effects on the immune responses by ensuring tolerance to auto-Ag (Fletcher and others 2010). Nevertheless, the appearance of ectopic TLO in settings of chronic autoimmunity encourages us to examine the potential of such therapeutic approaches. Time will tell whether a consideration of SLO topology will factor into vaccine design and therapies that treat significant human clinical problems.
Footnotes
Acknowledgments
The authors wish to acknowledge funding from the Canadian Institutes of Health Research (CIHR, MOPs 67157 and 89783 to J.L.G.) and from the Austrian Academy of Sciences (APART fellowship to J.H.F).
Author Disclosure Statement
No competing financial interests exist.