
Abstract
Cellular stress enhances inflammatory cytokine gene expression by inducing cEBP homologous protein (CHOP). Engaging cell stress via thapsigargin induced CHOP and selectively prolonged lipopolysaccharide-stimulated interleukin-6 (IL-6) expression in bone marrow-derived macrophages from wild-type (WT) but not CHOP knockout (KO) mice. To determine the impact of this mechanism in vivo we employed dextran sodium sulfate (DSS)-induced colitis in irradiated mice reconstituted with bone marrow from WT or CHOP KO mice. WT recipients of CHOP KO bone marrow exhibited more rapid recovery from disease than did mice reconstituted with WT bone marrow as reflected in increased survival, reduced clinical scores, and colonic histopathology. No differences in mesenteric lymph node cell populations were observed between mice with WT or CHOP KO bone marrow during colitis. CD11b+ macrophages infiltrating the lamina propria were, however, reduced in DSS-treated mice reconstituted with CHOP KO bone marrow. CHOP expression was observed within the infiltrating inflammatory CD11b+ macrophages. Furthermore, IL-6 expression within the inflamed colon was significantly lower in mice with CHOP-deficient bone marrow. Our findings indicate that CHOP expression in myeloid cells plays an important role in determining the magnitude and duration of inflammatory response in vivo by modulating expression of proinflammatory cytokines such as IL-6 in infiltrating macrophages.
Introduction
T
The production of cytokines in inflammatory cells is triggered through signaling events downstream of pattern recognition receptors [PRRs, eg, Toll-like receptors (TLRs)] that recognize and respond to molecular markers of injury or infectious challenge that couple directly or indirectly to altered transcriptional and post-transcriptional mechanisms (Akira and others 2006; Bauer and others 2009). Tissue injuries often result in cellular stress responses as a consequence of associated ischemic, oxidative, and genotoxic insults (Hotamisligil 2011; Hasnain and others 2012; Wang and Kaufman 2012). Such diverse events are often integrated through a collection of intracellular mechanisms that serve to correct deficiencies, prevent further cellular damage, restore normal structure and function, or, if necessary, initiate programmed cell death pathways (Ron and Walter 2007). Several of these pathways lead to the elevated expression of the cEBP homologous protein (CHOP), a transcription factor that may play both positive and negative roles in controlling the expression of a broad range of downstream genes (Li and others 2008; Namba and others 2009; Woo and others 2009). CHOP is often associated with promoting the induction of cellular apoptosis but can also directly impact on the patterns of inflammatory cell activation and promote inflammatory cytokine expression in a variety of cell populations including endothelium and macrophages (Zinszner and others 1998; Tabas and Ron 2011). Importantly, CHOP has also been reported to potentiate cytokine gene expression in myeloid cells following stimulation through PRRs including members of the TLR family (Li and others 2005; Goodall and others 2010).
In this study we determined the impact of cellular stress and the associated induction of CHOP on expression of several proinflammatory cytokines in mouse macrophages and further evaluated the importance of CHOP in regulating the magnitude or duration of myeloid inflammatory response in dextran sodium sulfate (DSS)-induced colitis in mice. Prior work reported that mice with targeted deletion of the CHOP gene (ddit3) showed reduced colon injury and inflammatory response following treatment with DSS, apparently reflecting the loss of CHOP-dependent apoptosis in the colonic epithelium (Namba and others 2009). To evaluate the importance of CHOP within the hematopoietic compartment in DSS colitis we utilized irradiated mice reconstituted with bone marrow from wild-type (WT) or CHOP-deficient mice. WT mice reconstituted with CHOP KO (KO) bone marrow exhibited more rapid recovery and prolonged survival following DSS treatment compared to mice with WT bone marrow. Furthermore, immunopathology revealed greater tissue damage and increased inflammatory CD11b+ macrophage infiltration in colon sections from DSS-treated WT mice reconstituted with WT compared to CHOP knockout (KO) bone marrow. The reduced disease severity correlated with selective reduction of interleukin-6 (IL-6) in colon tissue from mice reconstituted with CHOP KO bone marrow compared with recipients of WT bone marrow. Hence, CHOP expression within the hematopoietic compartment influences the magnitude and duration of DSS-colitis, perhaps reflecting its effect on expression of cytokines including IL-6 in inflammatory myeloid cells.
Materials and Methods
Reagents
Dulbecco's modified Eagle's medium (DMEM), Dulbecco's phosphate-buffered saline (PBS), RPMI 1640, sodium pyruvate solution,
Mice
Pathogen-free C57BL/6 mice (WT) were obtained from Charles River. CHOP KO mice were purchased from Jackson Laboratory and maintained within the Biological Resource Unit (BRU) of the Lerner Research Institute. All procedures were approved by the Institutional Animal Care and Use Committee.
Preparation of bone marrow-derived macrophages
Bone marrow cells were isolated from the femurs and tibia of 10- to 12-week-old donor mice (WT, CHOP KO) by flushing the bone marrow with Hanks' balanced salt solution (HBSS) into a sterile 50 mL tube. After centrifugation, the supernatant was aspirated, red blood cells were lysed in ACK lysing buffer. Cells were passed through a 100 μm strainer, centrifuged, and the pellets resuspended in complete DMEM media supplemented with 50 ng/mL of mouse M-CSF and cultured for 6 days. Bone marrow-derived macrophages (BMDM) were used in experiments as described in the text and figure legends.
Measurement of mRNA
Total RNA was prepared using TRI Reagent and analyzed either by northern blot hybridization or reverse transcription and real-time polymerase chain reaction using 1 μg total RNA as previously described (Datta 2008; Zhao and others 2014). Autoradiographs were quantified by image analysis using ImageJ software. CXCL1 and IL-6 mRNA levels were normalized to levels of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA measured in the same RNA sample. Specific primers for amplification of CXCL1, IL-6, and GAPDH are described previously (Zhao and others 2014).
Bone marrow reconstitution
Female C57BL/6 recipient (WT or CHOP KO) mice were irradiated twice with 600 rad with a 4-h interval between doses resulting in a total dose of 1,200 rad. Mice were reconstituted postirradiation with 1.5×107 donor bone marrow cells by tail vein injection using the following donor/recipient pairs: WT to WT, WT to CHOP KO, and CHOP KO to WT. After transfer, mice were intraperitoneally injected with 1.5 mg gentamicin; a second dose was given after 48 h. Hematopoietic reconstitution with donor cells was evaluated after 6 weeks by genotyping blood from tail vein and by flow cytometric analysis of spleen and mesenteric lymph node cell suspensions.
Induction of DSS colitis
Bone marrow chimera mice were subjected to 2.5% (w/v) DSS prepared in sterile drinking water for 7 days followed by regular water for the subsequent days. Mice were monitored daily for weight loss and clinical score. Clinical score was calculated based upon the severity of diarrhea and rectal bleeding each over a range of 0–3 with 0, none; 1, modest; 2, intermediate; and 3, severe. The sum of both measures was then the score used for each mouse.
Spleen and lymphoid tissue cell composition
Spleens and mesenteric lymph nodes were obtained from naïve and DSS-treated WT, CHOP KO and bone marrow-reconstituted mice and cell suspensions were collected by centrifugation and resuspended in FACS buffer [0.1% bovine serum albumin (BSA) in PBS]. Lamina propria cell populations were obtained from colon tissue as described previously (Do and others 2014). Colons from DSS-treated mice at day 8 or 11 were isolated and cleaned in HBSS. Colons were cut into small pieces and resuspended in HBSS with 0.5 μM EDTA and 15 μg/mL DTT and shaken for 15 min twice at room temperature. Colons were resuspended in complete RPMI with 400 μg/mL DNase and 1 mg/mL collagenase and shaken at 37°C for 90 min. The colon digest was passed through a 70-μM strainer and washed. Cells were resuspended in a 33% Percoll gradient and spun at room temperature for 20 min. Splenocytes and mesenteric lymph node cells were stained with FITC-conjugated CD4; PerCP-conjugated CD3; APC conjugated-CD8, Cd11c, and CD11b Abs. Lamina propria cell suspensions were surface stained with anti-CD4 (FITC), anti-CD3 (PerCP) anti-CD8 (APC), or anti-CD11b according to manufacturer's recommendations. Cells were analyzed using a FACSCalibur (Becton Dickinson, San Diego, CA).
Measurement of cytokines and chemokines from colon culture by ELISA
Segments from the distal colon of mice undergoing DSS colitis were washed with cold PBS and placed in 12-well flat-bottom culture plates containing serum-free RPMI supplemented with high concentration of penicillin/streptomycin to prevent bacterial growth and incubated at 37°C for 24 h. Supernatants were collected, centrifuged at 300 g, and stored at −20°C until analysis. Subsequently, CXCL1, IL-1β, and IL-6 levels were measured using ELISA according to the manufacturer's instructions and results were normalized by colon tissue weight.
Immunofluorescence microscopy
Distal colon tissues were embedded in O.C.T. compound and snap frozen in 2-methylbutane (isobutene). Frozen sections were air dried and fixed for 30 min with IC fixation buffer followed by blocking in 2% of BSA and 10× Triton-X for 1 h, and incubated with primary rat anti-mouse IL-6, rat anti-mouse CXCL1, or polyclonal rabbit anti-CHOP overnight at 4°C. After being washed in PBS, sections were incubated with the appropriate concentration of secondary antibodies for 2 h at room temperature. CD11b cells were identified using rat anti-mouse CD11b conjugated with Alexa Fluor 488. After staining, sections were washed in PBS and mounted in Vectashield mounting medium with DAPI. Colon sections from mice undergoing DSS colitis were stained with hematoxylin-eosin.
Statistical analysis
The data were expressed as the mean±standard deviation. Parametric data were analyzed using the Student's t-test: P<0.05 was considered significant.
Results
CHOP controls TLR-induced IL-6 expression in BMDM
Multiple studies have linked endoplasmic reticulum stress with enhanced inflammatory cytokine expression (Goodall and others 2010; Zeng and others 2010; Martinon and Glimcher 2011). Treatment of BMDM with Tg, an inhibitor of endoplasmic reticulum Ca++ ATPase activity known to induce cellular stress responses, resulted in substantially elevated expression of CXCL1 and IL-6 following stimulation with LPS (Fig. 1A). The effects of Tg-induced cell stress are seen with a broad spectrum of TLR agonists including agents that activate through TLR2 (P3C), TLR3 (pIC), TLR4 (LPS), and TLR7 (R848) (Fig. 1B). Enhanced expression of IL-6 during cell stress has been previously reported to depend upon expression of CHOP. In BMDM from CHOP KO mice the stress-dependent amplification of IL-6 mRNA and protein was compromised (Fig. 1C, D). Though expression of CXCL1 and other cytokines including IL-1β (not shown) was also enhanced by stress, CHOP deficiency had less impact on expression of these inflammatory gene products.
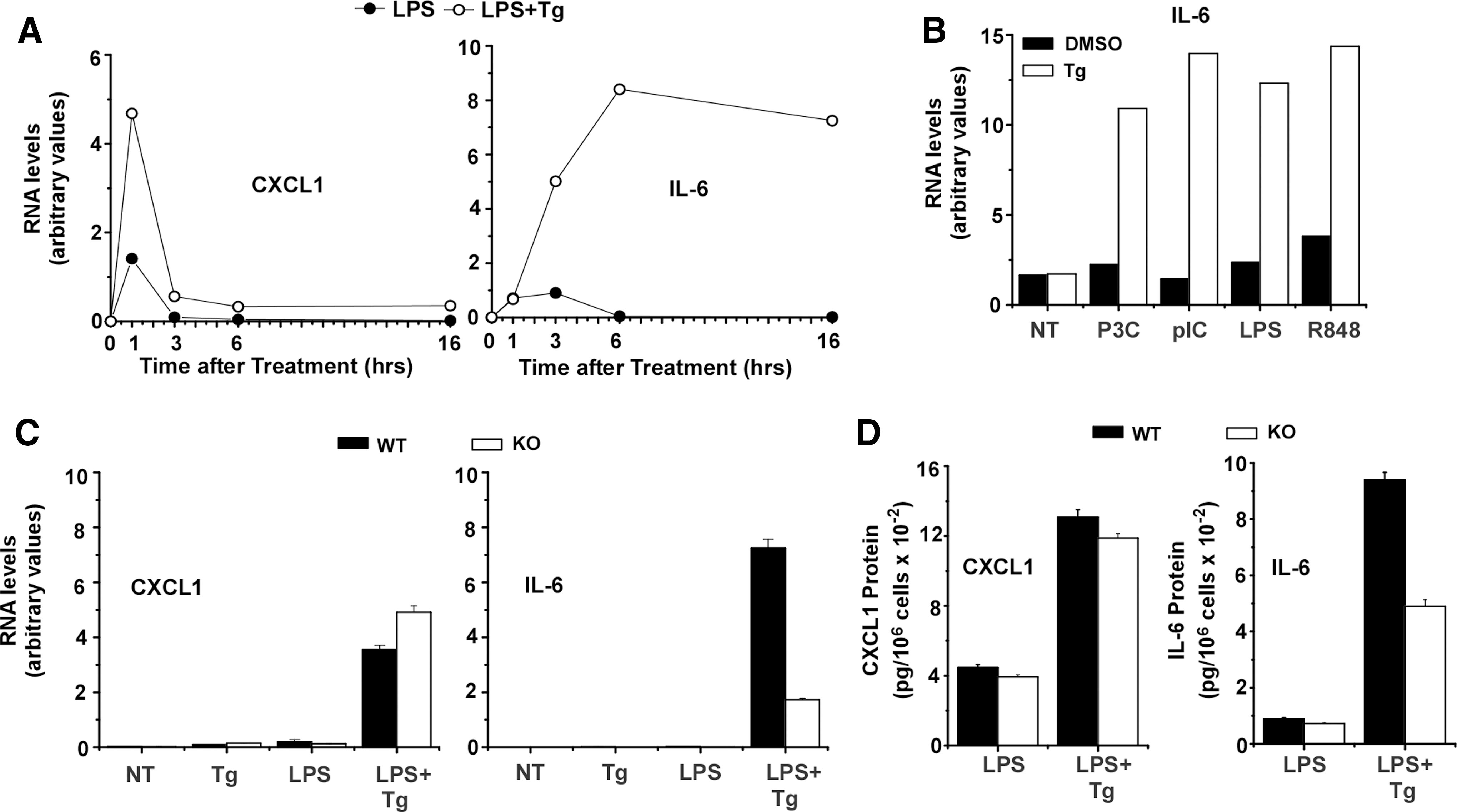
CHOP is necessary for stress to enhance LPS-induced IL-6 expression in BMDM.
Characteristics of irradiated mice reconstituted with WT or CHOP KO bone marrow
Mice with targeted deletion of the CHOP gene have been reported to exhibit reduced disease severity in the mouse model of colitis induced by DSS (Namba and others 2009). Because TLR-stimulated macrophages obtained from CHOP KO mice have reduced cytokine expression (eg, IL-6), we reasoned that CHOP deficiency in the myeloid cell compartment might alter the course of DSS-induced colitis. To determine the effects of CHOP gene deficiency in the hematopoietic compartment we created chimeric mice using total body irradiation of WT mice followed by reconstitution with bone marrow from WT or CHOP KO mice. Fully chimeric mice were used to evaluate different hematopoietic cell populations in cell suspensions prepared from the mesenteric lymph nodes and the spleen (Fig. 2A, B). The percentages of CD4+ and CD8+ T cells, CD19+ B cells, CD11b+ myeloid cells, and CD11c+ dendritic cells were comparable in mice reconstituted with WT or CHOP KO bone marrow. Similar findings were obtained when CHOP KO mice were reconstituted with WT bone marrow (not shown). While WT and CHOP KO mice were given equivalent doses of radiation before reconstitution, mixed chimerism was occasionally seen in CHOP KO mice reconstituted with WT bone marrow likely due to the greater resistance to radiation-induced cell death in this genotype. Only mice that exhibited apparent full chimerism (based upon genotype of peripheral blood mononuclear cell) were used in experiments.
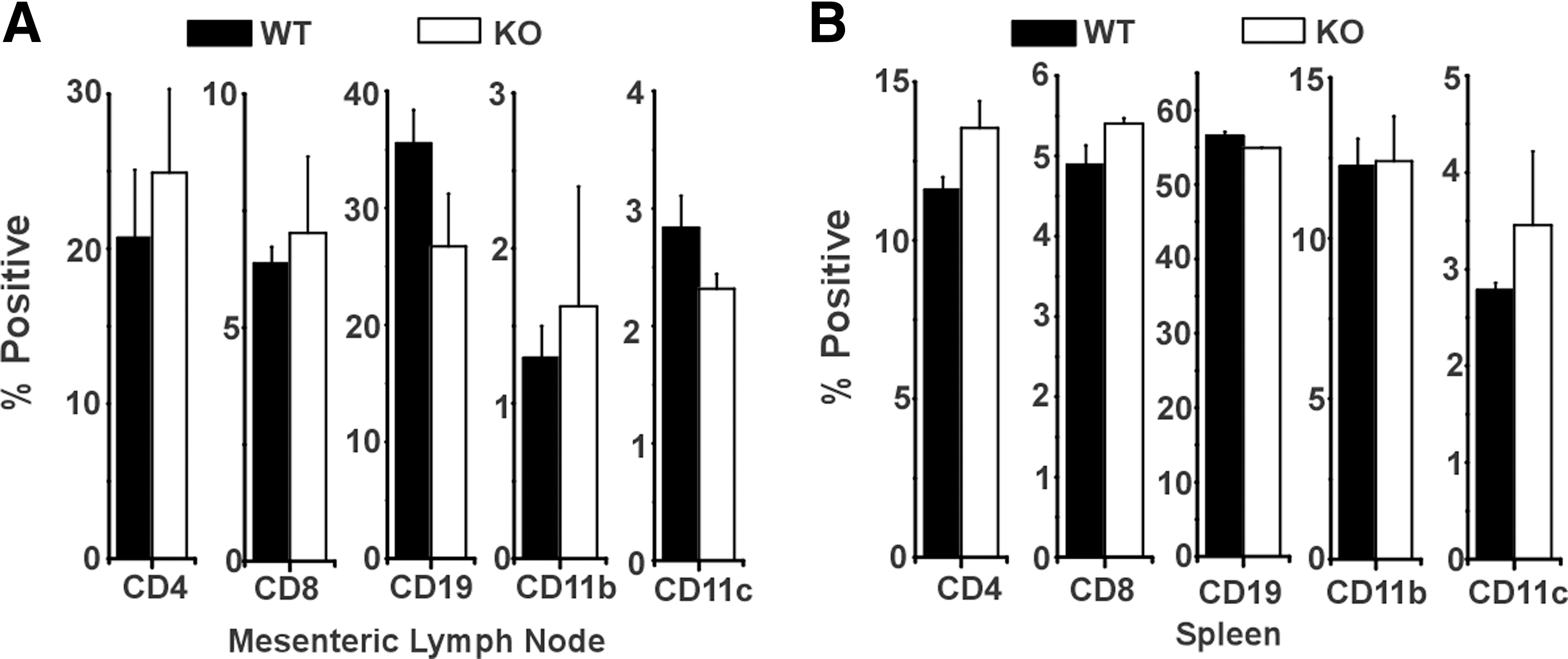
WT mice with KO bone marrow have normal hematopoietic cell reconstitution. WT mice were irradiated and reconstituted with either WT or KO bone marrow. After 6 weeks of recovery, mice were sacrificed and mesenteric lymph nodes
CHOP KO bone marrow protects mice from DSS colitis
WT mice reconstituted with WT or CHOP KO bone marrow were compared following exposure to 2.5% DSS in the drinking water for 7 days. All animals developed clinical evidence of colitis (diarrhea and rectal bleeding) within 7 days after exposure to DSS though the severity of such symptoms was reduced in mice reconstituted with KO bone marrow (Fig. 3A). Furthermore, mice with WT bone marrow continued to decline and 80% died between 8 and 11 days after the initiation of DSS treatment. In contrast, mice with bone marrow from CHOP KO mice began to recover following removal of DSS and 60% survived beyond 11 days (Fig. 3B). While all animals lost weight comparably during the initial 7 days period, the differential disease severity was reflected in the number of mice with CHOP-deficient bone marrow that gained weight between 8 and 11 days (Fig. 3C). In a separate experiment we also examined irradiated CHOP KO mice that were reconstituted with bone marrow from WT donors. In comparison with WT mice receiving WT bone marrow, CHOP KO mice reconstituted with WT bone marrow exhibited delayed development of clinical symptoms and weight loss (Fig. 3D, F). Though CHOP deficiency in either compartment (hematopoietic vs. nonhematopoietic) reduced susceptibility to DSS colitis, WT mice with CHOP KO bone marrow significantly differed from CHOP KO mice receiving WT bone marrow in that the former group showed significant recovery from disease while all mice in the latter group died (compare Fig. 3B and E). This finding confirms the prior report that CHOP deficiency in the nonhematopoietic compartment contributes to the disease phenotype, most likely by reducing the magnitude of colonic epithelial injury. This mechanism, however, produces a distinct outcome (reduced magnitude of injury) compared to that seen when CHOP deficiency is restricted to inflammatory macrophages (frequency of recovery).
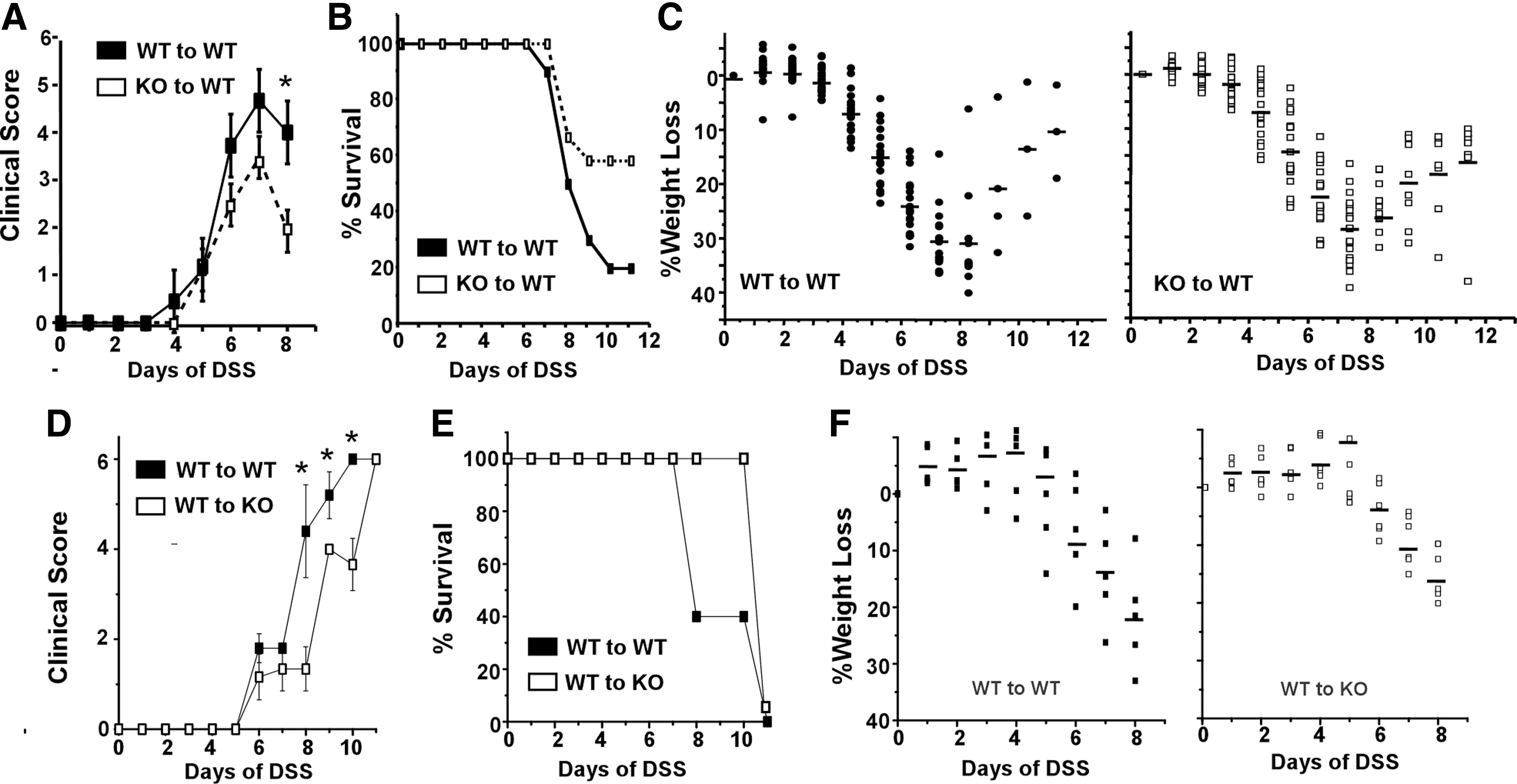
CHOP contributes to the severity of DSS-induced colitis.
Hematopoietic CHOP deficiency alters gut inflammation
Histologic analyses of sections from the distal colon were consistent with the measures of clinical disease (Fig. 4A). Tissues harvested from WT mice reconstituted with either WT or CHOP KO bone marrow on day 8 exhibited comparable evidence of destruction of the colonic epithelium with swelling of the submucosal layer and substantial pockets of inflammatory infiltrate. Differences between groups were, however, apparent in sections taken from mice sacrificed on day 11. While sections prepared from WT recipients of CHOP KO and WT bone marrow both showed substantial epithelial hyperplasia at 11 days, mice with WT bone marrow still had areas without epithelial cover and substantially greater inflammatory cell infiltration compared with mice reconstituted with CHOP KO bone marrow. The decrease in inflammation was confirmed by staining sections from mice harvested at day 8 versus day 11 for CD11b+ myeloid cells (Fig. 4B). This revealed comparable numbers of inflammatory cells within the lamina propria from mice with WT or CHOP KO bone marrow on day 8. There was, however, a significant reduction in inflammatory CD11b+ cells in sections from mice with CHOP KO bone marrow compared to mice with WT bone marrow when measured on day 11. There were no differences in the relative number of CD4 or CD8 T cells within the draining mesenteric lymph nodes in any mice (Fig. 4C). Consistent with the histologic findings, however, the number of CD11b+ cells in the lamina propria of mice with CHOP-deficient bone marrow was markedly reduced (Fig. 4D).
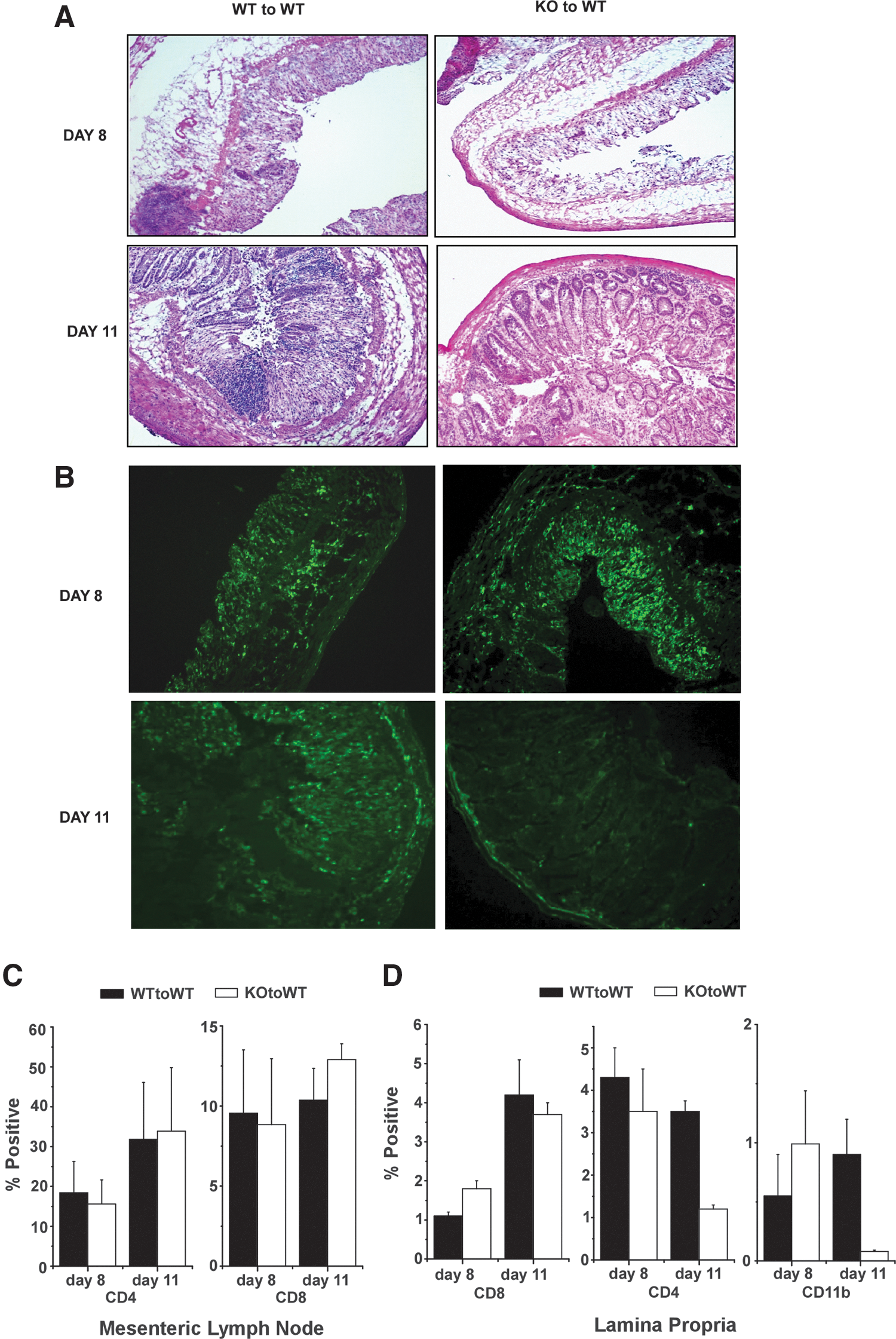
Colon histology in DSS exposed WT mice reconstituted with WT or KO bone marrow.
CHOP is expressed in CD11b+ cells in inflamed colon
The effects of CHOP deficiency in the hematopoietic cells on DSS-induced colitis raises the question of whether CHOP is expressed within hematopoietic cell populations in the inflamed colon. Sections prepared on day 8 from DSS-treated WT mice reconstituted with either WT or CHOP KO bone marrow were stained with antibody against CD11b+ to mark myeloid cells (green) and antibody against CHOP (red). Mice with WT or CHOP KO bone marrow both exhibited substantial CD11b+ cell infiltration by 8 days of DSS (Fig. 5). Consistent with previous work, the expression of CHOP was readily detectable in sections of colon from DSS-treated mice (Namba and others 2009). While the colonic epithelium appeared to represent the majority of CHOP-expressing cells, cells staining for both CD11b+ and CHOP (yellow in the merged panel of Fig. 5) were observed in sections from mice with WT bone marrow but not (as expected) in tissues from mice with CHOP KO bone marrow. Specificity of CHOP antibody was tested in several ways (Supplementary Fig. S1; Supplementary Data are available online at
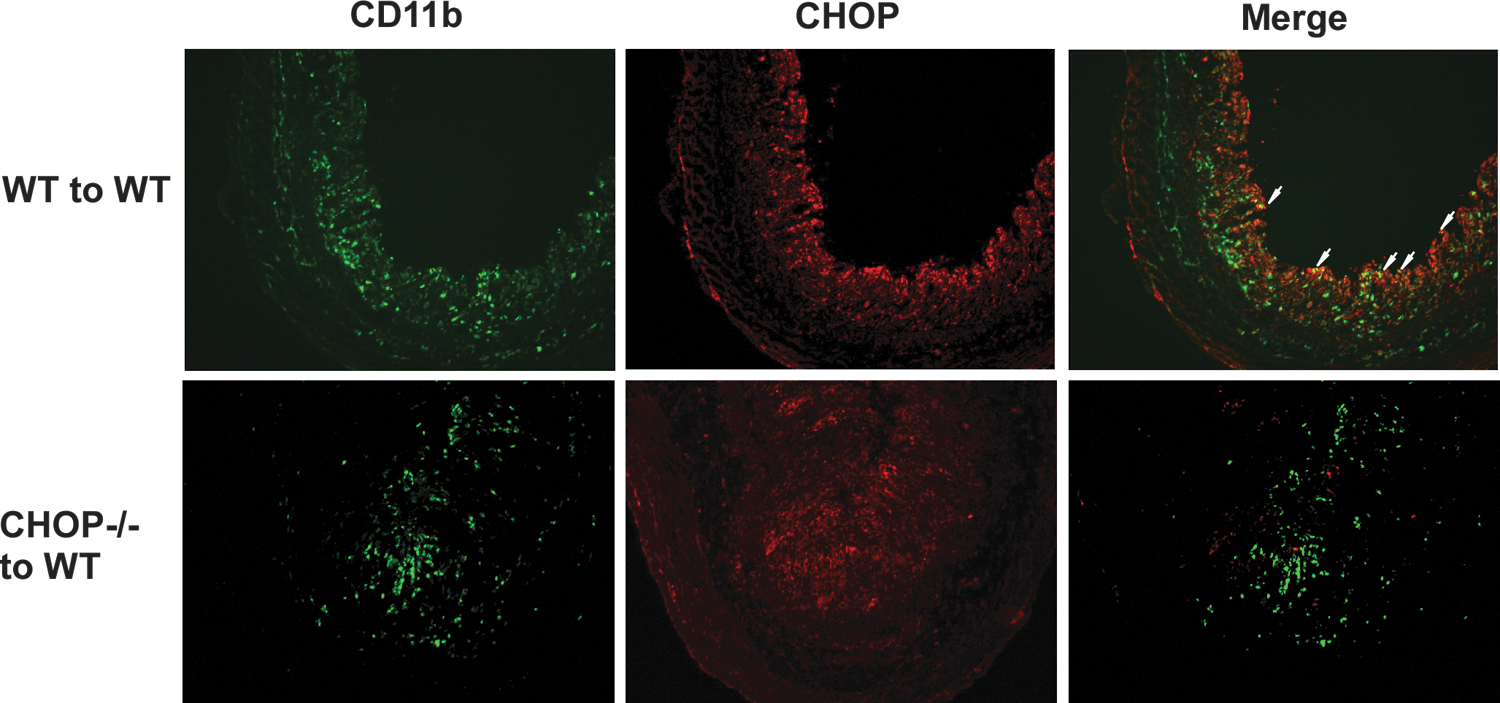
CHOP is expressed in CD11b+ myeloid cells in the colon of DSS-treated mice. WT mice reconstituted with WT or KO bone marrow were given 2.5% DSS in drinking water for 7 days. Mice were sacrificed on day 8 and colon tissue was dissected, embedded in OCT compound, and snap frozen in isobutene. Frozen sections were stained for antibodies against CD11b (green) or CHOP (red). The CD11b/CHOP images were merged (yellow) and double positive cells are indicated by white arrows. Similar results were obtained in 4 separate mice of each chimeric genotype.
IL-6 expression is selectively reduced during colitis in mice with CHOP KO bone marrow
IL-6 is known to be an important determinant of the inflammatory response during colitis and the severity of DSS-induced colitis is reduced in IL-6-deficient mice (Naito and others 2004; Matsumoto and others 2010). The effects of CHOP deficiency on expression of IL-6 (Fig. 1) and on the severity of DSS-colitis suggested that mice with CHOP KO bone marrow might exhibit reduced expression of IL-6 in association with the increase in frequency of recovery. To test this, we determined the frequency of CD11b+/IL-6+ cells within the colon by immunofluorescence microscopy (Fig. 6A). While IL-6-expressing inflammatory cells were present at comparable levels in recipients of WT and CHOP KO bone marrow on day 8, there was marked reduction in CD11b+ cells staining for IL-6 from mice with CHOP-deficient bone marrow on day 11. This differential was also evident in measurement of IL-6-secreted protein in supernatants from cultured colon tissue (Fig. 6A). Interestingly, expression of the chemokine CXCL1 was quantitatively comparable in both WT and CHOP KO bone marrow recipients despite the reduction in frequency of CD11b+ cells seen in CHOP KO tissues (Fig. 6B). This is consistent with the finding that CXCL1 expression appears to be largely within the colon epithelium. Hence, the magnitude of inflammatory cytokine expression within the myeloid cell compartment during colitis is dependent upon the stress-induced CHOP gene. Measurements of IL-1β in colon culture supernatants did not show significant differences between mice with WT or KO bone marrow (not shown).
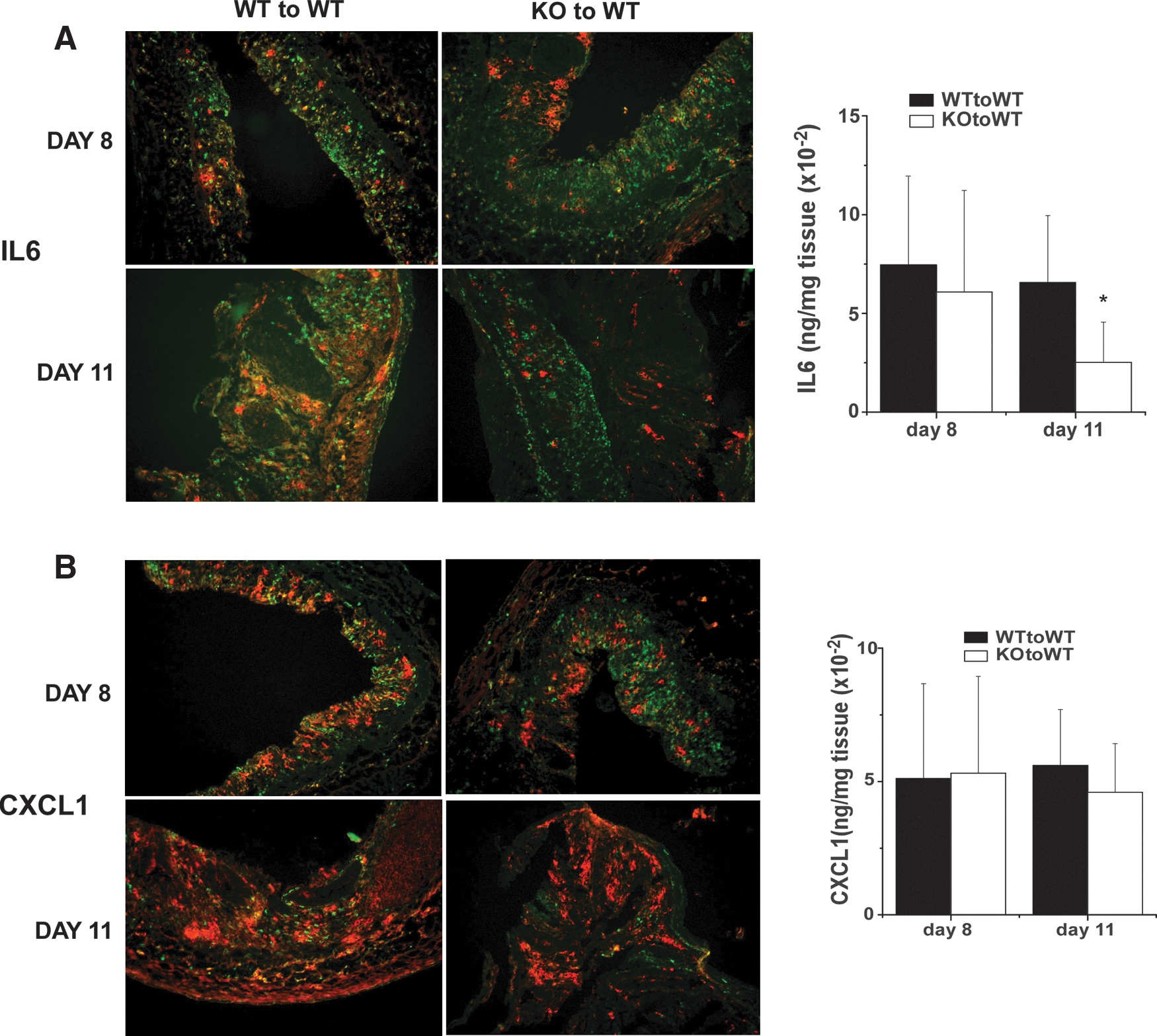
Differential IL-6 and CXCL1 expression in recipients of WT or KO bone marrow during DSS-colitis. Colon tissue from mice treated with DSS for 7 days and sacrificed at 8 or 11 days as described in the legend to Fig. 4 were stained CD11b (green) and for IL-6
Discussion
CHOP (ddit3) is well known as a cell stress-inducible gene that contributes to apoptosis (Wang and Ron 1996; Zinszner and others 1998; Marciniak and others 2004). In this context it has been reported that mice with targeted deletion of the CHOP gene are protected from DSS colitis due to reduced apoptosis in colonic epithelial cells and diminution of the intestinal damage and the accompanying inflammatory sequellae in this model of colitis (Namba and others 2009). CHOP has also been implicated in magnifying proinflammatory cytokine gene expression within myeloid cell populations (Hattori and others 2003; Li and others 2005; Chen and others 2009; Goodall and others 2010). The objective of this study was to explore the potential role of CHOP-dependent control of myeloid cell inflammatory cytokine expression in inflammatory disease using the DSS-colitis model. The results support the following conclusions: (a) prolonged cytokine gene expression in stressed BMDM is dependent upon CHOP. (b) Chimeric mice reconstituted with CHOP KO bone marrow have reduced duration of disease severity in DSS colitis. While the initiation of disease is comparable in mice with either WT or CHOP KO bone marrow, those with CHOP deficiency in the hematopoietic compartment recover more efficiently. (c) CHOP KO mice reconstituted with bone marrow from WT mice also exhibit protection from colitis but the pattern of protection when CHOP is operative in hematopoietic versus nonhematopoietic compartments is distinct. CHOP KO recipients of WT bone marrow develop disease more slowly than WT recipients of WT bone marrow though the severity of disease is ultimately comparable. (d) Though the number and percentage of CD4 and CD8 T cells in the draining mesenteric lymph nodes are comparable in mice reconstituted with either WT or CHOP KO bone marrow, the infiltration of CD11b+ myeloid cells within the lamina propria is selectively reduced at later times and this is accompanied by a reduction in IL-6 cytokine expression. Furthermore, CHOP is detectably expressed within the infiltrating CD11b+ inflammatory cell population.
Prior reports demonstrate that CHOP is an important mechanistic feature contributing to the ability of cellular stress to promote increased magnitude and/or duration of inflammatory cytokine gene expression (Hattori and others 2003; Li and others 2005; Chen and others 2009; Goodall and others 2010). These effects have been reported to exhibit some gene specificity, with IL-6 and IL-23p19 being among the most sensitive target genes (Li and others 2005; Goodall and others 2010). Our results demonstrate that the ability of coincident engagement of TLR stimulation and cell stress results primarily in prolonging the duration of IL-6 expression. This is likely to reflect the time of appearance of CHOP and of cEBPβ, known to be a major transcription factor controlling IL-6 production (Akira and others 1992). While CHOP itself has limited DNA-binding activity and targets a distinct nucleotide sequence compared to other members of the cEBP family, it can form heterodimers with other cEBP factors (Hattori and others 2003; Li and others 2008). Indeed, it has been suggested that the mechanism by which CHOP expression enhances IL-6 transcription is through formation of heterodimers with the liver enriched inhibitory protein (LIP) isoform of cEBPβ, which is known to exhibit inhibitory activity (Hattori and others 2003). Thus, the appearance of CHOP might serve to sequester LIP and enable the active isoform of cEBPβ to engage transcription at the IL-6 promoter. Interestingly, other cell stress response mechanisms have also been connected to enhanced production of IL-6, particularly involving the spliced form of X box-binding protein 1 (XBP1) (Martinon and others 2010). In this regard, we have recently reported that the TLR-induced expression of multiple proinflammatory chemokines and cytokines, including IL-6, is amplified in mouse macrophages when cell stress is engaged before TLR engagement (Zhao and others 2014). Though this mechanism appears to be independent of both CHOP and XBP1, it is evident that cell stress can broadly impact on cytokine expression.
It is noteworthy that WT mice with CHOP deficiency in the bone marrow exhibit a distinct colitic phenotype in comparison with CHOP KO mice that have been reconstituted with WT bone marrow. WT mice with WT or CHOP KO bone marrow lose weight with similar kinetics but a greater proportion of mice with CHOP KO bone marrow recover while recipients of WT bone marrow decline and 80%–100% of animals die by 11 days after initiation of DSS exposure. In contrast, CHOP KO mice with WT bone marrow lose weight and develop clinical signs more slowly than WT mice with WT bone marrow but both ultimately succumb to disease by 11 days. This latter finding confirms the observations made using mice with complete CHOP deficiency and likely reflects the role of CHOP in promoting stress-mediated colon epithelial apoptotic death (Namba and others 2009). CHOP deficiency in the bone marrow appears to reduce the magnitude and duration of the inflammatory response within infiltrating inflammatory leukocytes with the outcome reflecting more rapid tissue recovery. Irradiation followed by bone marrow reconstitution replaces multiple immune cell types including T and B lymphocytes and many myeloid cell populations. CHOP deficiency could alter the nature of the response of all of these cell populations to DSS-driven disruption of colon epithelium. Though we cannot rule out contributions from lymphoid populations, the evidence favors a role for myeloid cells with respect to the protective impact of CHOP deficiency. All hematopoietic cell populations were comparably restored following bone marrow reconstitution (Fig. 2) and there were no significant differences in the number or percentage of T-cell and B-cell populations in the draining mesenteric lymph nodes between mice with WT or CHOP KO bone marrow at either day 8 or 11. In contrast, the number of CD11b+ cells within the lamina propria of recipients of CHOP KO bone marrow was markedly reduced at day 11. Hence, this difference appears likely to reflect the impact of CHOP in controlling the duration of inflammatory cytokine gene expression within the myeloid compartment. Our studies do not discriminate between monocyte/macrophage and neutrophils and both cell populations are potential contributors to the differential outcomes obtained.
The reduced inflammation and increased recovery and survival of mice with CHOP deficient myeloid cells are reflected in the reduction in IL-6 expression in colonic tissue at later stages of disease. Importantly, IL-6 is well known to be an important cytokine in controlling the colitic response to DSS exposure (Naito and others 2004; Matsumoto and others 2010) and its neutralization results in the enhanced recovery from disease, reminiscent of the effects of hematopoietic CHOP deficiency seen in our experiments (Sommer and others 2014). Thus, the ability of CHOP to participate in controlling TLR-stimulated IL-6 expression within myeloid cells may be at least partially responsible for the protection from DSS colitis afforded by animals with CHOP KO bone marrow. It should be noted, however, that IL-6 has also been closely linked with colon epithelial cell regeneration and hence the functional consequences may be complex (Bollrath and others 2009; Grivennikov and others 2009; Kuhn and others 2014). Importantly, IL-6 function is closely aligned with levels of soluble GP130 and we have not determined the impact of CHOP deficiency on this parameter. Also, it is important to keep in mind that the reduction in IL-6 levels occurs in parallel with reduction in inflammatory cell content of the colon tissue. Nevertheless, the reduction in cytokine expression may also reflect the mechanisms demonstrated in BMDM (Fig. 1). In this regard, though CHOP is strongly expressed within the colonic epithelium, we observe co-distribution of cells expressing both CHOP and CD11b in mice with WT bone marrow that is not detectable in mice reconstituted with CHOP-deficient bone marrow. These findings support the hypothesis that CHOP and other molecular features of cellular stress have a significant role in determining the characteristics of inflammatory function within infiltrating leukocytes and hence may represent potential targets for therapeutic intervention in inflammatory disease settings.
Footnotes
Acknowledgment
This work was supported by USPHS Grant CA039621.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.