
Abstract
Within monocyte heterogeneity, subsets represent discrete, well-characterized phenotypes. Although many studies have highlighted differences between subsets, there is evidence that subpopulations represent contiguous stages in a maturational series. As CD14hiCD64hi monocytes have higher proliferative potential than CD14hiCD64lo monocytes, the surface marker profile on 4 subsets defined by CD14 and CD64 was measured. The profiles were compared to that of subsets defined by the high-affinity IgE receptor (FcɛRIα), CD16, and CD14; further differences in size, granularity, and buoyancy were measured in subsets delineated by these markers. There was a positive correlation between proliferative monocyte (PM) prevalence and CD64 expression on the classical monocyte subset, and also between PM prevalence and circulating FcɛRIα+ monocytes. The expression of CD64, the high-affinity IgG receptor, on canonical human monocyte subsets was determined before and after short-term culture, and in response to interleukin (IL)-6, IL-10, macrophage colony-stimulating factor, granulocyte/macrophage colony-stimulating factor and interferon-γ; the influence of these cytokines on monocyte subset transition was also measured. The loss of FcɛRIα expression preceded an increase in CD16 expression in whole blood cultures. These data indicate that high-affinity Fc receptors are expressed on less mature monocytes and that FcɛRIα+ monocytes are developmentally antecedent to the canonical classical and intermediate monocyte subsets.
Introduction
M
Although schemes based upon the expression of CX3CR1, CD2, CD13, CD33, or CD64 have been used (Grage-Griebenow and others 1993; Thomas and others 1993; Crawford and others 1999; Geissmann and others 2003), human monocytes are now classified into subpopulations based upon differential expression of CD14 and CD16 (Passlick and others 1989; Ziegler-Heitbrock and others 2010); the majority of monocytes exhibit the classical subset phenotype (CD14hiCD16−), typically 10% have the nonclassical phenotype (CD14loCD16++) leaving the intermediate subset as the least prevalent subset in the steady state (Ziegler-Heitbrock and others 2010). Differences in transcriptional profiles (Ancuta and others 2009) and tissue homing (Geissmann and others 2003) have confirmed functional differences observed in vitro; however, few studies have attempted to determine the developmental relationships between monocyte subsets. A proliferative monocyte (PM) subpopulation has been identified, which is ostensibly less mature than the bulk of the monocytes, and provides evidence for differences in maturity between human monocyte subsets (Cheung and Hamilton 1992; Clanchy and others 2006; Clanchy and Hamilton 2012b). Monocytes with relatively high expression of CD64 have higher proliferative potential compared to monocytes with low expression of CD64 (Clanchy and others 2006), indicating that less mature, more proliferative CD14hi monocytes have higher expression of CD64.
A comparison of surface marker profiles indicated that the expression of the high-affinity Fc receptor for IgE (FcɛRIα) significantly overlapped with that of monocytes with the highest expression of CD64, which is also a high-affinity Fc receptor. As PM have increased osteoclastic potential (Clanchy and others 2006) and an as yet undetermined role in human physiology, the aim of this study was to determine the relationship between CD64 expression and monocyte maturity or activation on canonical human monocyte subsets, which are defined by expression of CD14 and CD16 (Ziegler-Heitbrock and others 2010). Subsets defined by FcɛRIα, CD16, and CD14 were also characterized and the prevalence of PM was measured.
Materials and Methods
Isolation and culture of PBMC
Human leukocytes were isolated from buffy packs as described previously (Clanchy and others 2006). After centrifugation, the peripheral blood mononuclear cell (PBMC) layer was transferred to a new tube and washed repeatedly with phosphate-buffered saline (PBS) to deplete platelets. For cytokine stimulation experiments, PBMC were cultured at a concentration of 2.5 × 106/mL of medium [RPMI with 10% fetal bovine serum (FBS), 1% P/S]. Cytokines (PeproTech) were added to media at the following concentrations—interleukin-6 (IL-6) 50 ng/mL, interleukin-10 (IL-10) 20 ng/mL, macrophage colony-stimulating factor (M-CSF) 20 ng/mL, granulocyte/macrophage colony-stimulating factor (GM-CSF) 20 ng/mL, or interferon-γ (IFN-γ) 20 ng/mL.
Whole blood culture
Medium [1% P/S, 10% FBS, alpha-minimal essential medium with 2 mM
CFSE-labeled cell tracking
Whole blood cultures (WBC) from buffy packs were initiated as described. PBMC from the same donor were isolated as described above. After labeling PBMC with FACS antibodies (FcɛRIα-FITC and CD14-PE), monocytes were sorted. The sorted cells were resuspended at 1 × 105 cell/mL in PBS. Carboxyfluorescein succinimidyl ester (CFSE, Invitrogen) was added to a final concentration of 0.5 μM and cells were incubated for 10 min at room temperature with frequent mixing. An equal volume of FBS was added and cells were washed and added to the WBC for the remaining ∼12 h of the assay. PBMC from WBC were isolated using Ficoll-Paque then stained with CD16-PerCP and CD14-APC; the phenotype of the labeled monocytes was assessed by gating on CFSE+ cells.
Buoyancy
Percoll (Sigma-Aldrich) was diluted with 10× PBS to make iso-osmotic 100% Percoll, which was diluted with PBS for lower concentrations of Percoll. For determining the relative buoyancy of monocyte subsets, 3 mL of 30% Percoll was layered over 3 mL of 40% Percoll in a 10 mL tube. PBMC (∼1.5 × 106) were resuspended in 3 mL of PBS and layered over the 30% Percoll. The tube was spun at 1,000 g for 20 min with no brake. Cells from the interfaces and soft pellet were analyzed by flow cytometry. Leukocytes accumulating at Percoll interfaces were defined as low density (upper interface), high density (lower interface), and pellet. To determine the relative subset proportion at each interface, the percentage of a subset at each interface was expressed relative to the original subset percentage, before Percoll separation. To determine the relative subset buoyancy for each subset, the relative subset proportion of the low-density interface was expressed relative to the relative subset proportion of the high density. See Supplementary Fig. S1 (Supplementary Data are available online at
Flow cytometry
All antibodies were from BD Biosciences, except for FcɛRIα-FITC (Abcam, Inc.); isotype and compensation controls were used to set gates for sorting and analysis; propidium iodide (Sigma-Aldrich) was used to identify nonviable monocytic cells (identified by forward scatter [FSC] vs. side scatter [SSC]), which were of similar prevalence in in vitro cultures (Supplementary Fig. S2). For cell sorting, PBMC were stained with 10 μL of antibody for 25 min on ice in the dark then washed 3 times with FACS buffer. Sorting was performed using a FACSVantageSE (BD Biosciences). FACS acquisition was performed with a FACSCalibur flow cytometer and subsequent analysis was performed using FlowJo (Treestar).
BrdU assay
FACS-sorted CD14hi monocytes were assessed for cell cycle entry as previously described (Clanchy and others 2006). Briefly, monocytes were cultured in 96-well U-bottom plates at a concentration of 5 × 104 cells per well in 200 μL of media (α-MEM with 3% human serum, 1% P/S, 2 mM
Statistics
Significant differences were determined by Student's t-test or analysis of variance with post hoc analysis as detailed in figure legends. Significant correlations were determined by Pearson correlation coefficient. Analyses were performed with Excel (Microsoft) and Prism (GraphPad).
Results
CD64-defined subsets delineate a maturing monocyte
The monocytic expression of CD64 is highest on CD14hi monocytes, particularly the CD16− classical subset, which accounts for the majority of monocytes (Grage-Griebenow and others 1993, 2001b). As proliferative potential is higher in monocytes with the CD14hiCD64hi expression, compared with CD14hiCD64lo monocytes, the expression of several well-characterized myeloid surface markers was measured on subsets of monocytes defined as CD14hiCD64hi, CD14hiCD64med, CD14hiCD64lo, and CD14loCD64lo to determine whether these subsets might represent a maturation series, as evidenced by corresponding changes in the surface marker expression.
Previously, PM was determined to have a CD11bloCD33hi CD13loCD64hi phenotype, where high and low expressions were defined as the highest and lowest ∼15% of expression of a particular surface marker on CD14hi monocytes (Clanchy and others 2006). To further refine the description of PM, the expression of surface markers was measured on peripheral blood monocytes divided into subsets based upon expression of CD14 and CD64. High and low expressions were defined as the highest and lowest ∼20% of expression within CD14hi monocytes for each donor and intermediate expression of CD64 was defined as the 20% of CD14hi monocytes that had an expression of CD64 equidistant between the high and low gates; CD14loCD64lo monocytes accounted for 5%–10% of monocytic cells (Fig. 1A).
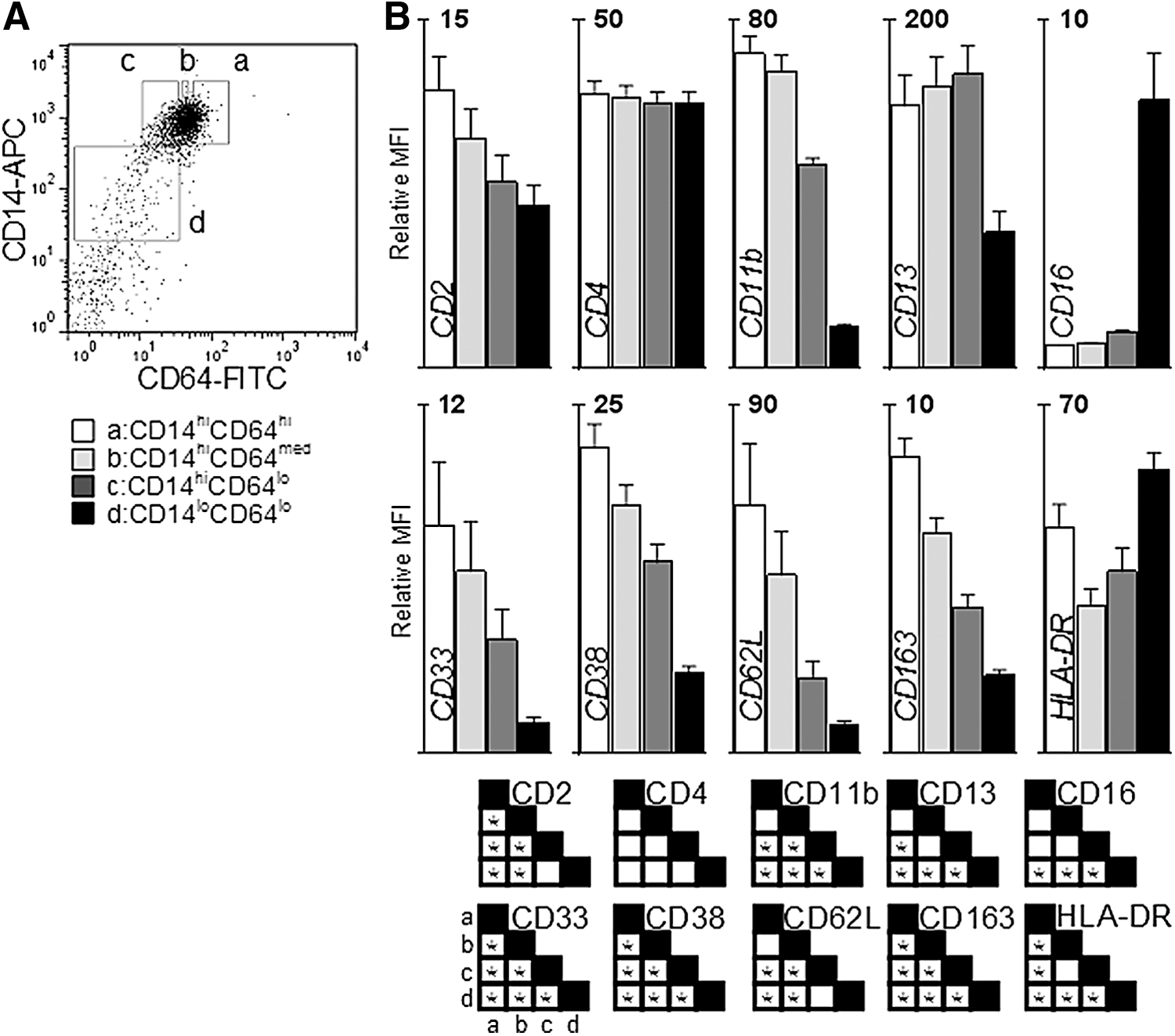
Relative CD64 expression is indicative of a maturing monocyte surface profile.
As shown in Fig. 1B, the expression of CD16 was low on CD14hiCD64hi/int monocytes, slightly higher on CD14hiCD64lo monocytes, and highest on the CD14loCD64lo subset, as has previously been described (Grage-Griebenow and others 1993, 2000). The expression of CD2, CD11b, CD33, CD38, CD62L, and CD163 was highest on CD14hiCD64hi monocytes and is reduced at a uniform rate across CD14hiCD64med, CD14hiCD64lo, and CD14loCD64lo monocytes. CD14hi monocytes with low expression of CD13 were previously found to have higher proliferative potential compared to CD14hiCD13hi monocytes (Clanchy and others 2006) and CD14hiCD64hi monocytes have lower expression of CD13 compared to CD14hiCD64lo monocytes. HLA-DR was expressed in a bi-modal fashion, with highest expression on the CD14hiCD64hi and CD14loCD64lo monocytes, but lower on the other subsets. The expression of CD4 was similar in all subsets.
FcɛRIα, CD16, and CD14 define 5 monocyte subsets
The expression of CD2 was higher on CD64hi monocytes. Monocytes which express higher levels of CD2 have been described as a putative DC precursor since, in the presence of IL-4 and GM-CSF, they differentiate rapidly into CD83+ cells with higher antigen presentation potential compared to that in CD2− monocytes (Crawford and others 1999; Di Pucchio and others 2003). These monocytes bear some resemblance to CD14loCD16−CD33hi monocytes, which have been characterized as myeloid DC precursors as they display a high antigen presentation capacity, DC morphology, and DC FACS profile after culture (Thomas and others 1993; Fagnoni and others 2001); the high expression of CD2 on monocytes from patients with myeloproliferative disorders further indicates a lack of monocyte maturity associated with this marker (Shoda and others 2005). The CD2hi monocyte subset has been reported to express the high-affinity receptor for IgE (FcɛRIα), the expression of which on monocytes correlates with the amount of IgE present in the blood in allergic conditions (Cheng and others 2006).
The expression of CD14, CD16, and FcɛRIα was measured on freshly isolated cells and is shown in Fig. 2A–C, demonstrating the mutually exclusive expression of CD16 and FcɛRIα on both CD14hi and CD14lo monocytes. When defined by the expression of CD14, CD16, and FcɛRIα, monocytic cells can be divided into 5 subsets—CD14loFcɛRIα+, CD14hiFcɛRIα+, CD14hi (FcɛRIα/CD16)−, CD14hiCD16+ (the intermediate subset), and CD14loCD16++ (the nonclassical subset); CD14hiFcɛRIα+ and CD14hi(FcɛRIα/CD16)− comprise the classical subset. The expression of common myeloid cell surface markers was measured on each subset (Fig. 2D). While CD34 (not shown) was not significantly expressed, generally the pattern of expression across the 5 subsets falls into 3 categories. CD33 and CD38 had a similar profile with highest expression on the CD14hiFcɛRIα+ subset and much lower expression on the nonclassical CD14loCD16++ subset. CD2, CD62L, and CD163 were expressed at higher levels on FcɛR1α+ monocytes, but reduced at varying rates in a stepwise fashion on the other populations. CD11b and CD13 have high expression on the most prevalent CD14hi(FcɛRIα/CD16)− subset and lowest expression on both CD14lo subsets; this pattern is inverted for HLA-DR. Of particular interest was the CD11bloCD13loCD33hiCD64hi phenotype of CD14hiFcɛRIα+ monocytes, as cells expressing this combination of markers have previously been shown to have higher proliferative potential, and are thus potentially less mature (Clanchy and others 2006). The prevalence of each subset varied from donor to donor, although FcɛRIα+ monocytes were less abundant than CD16+ monocytes, which were in turn far less abundant than CD14hi(FcɛRIα/CD16)− monocytes (Fig. 3A).
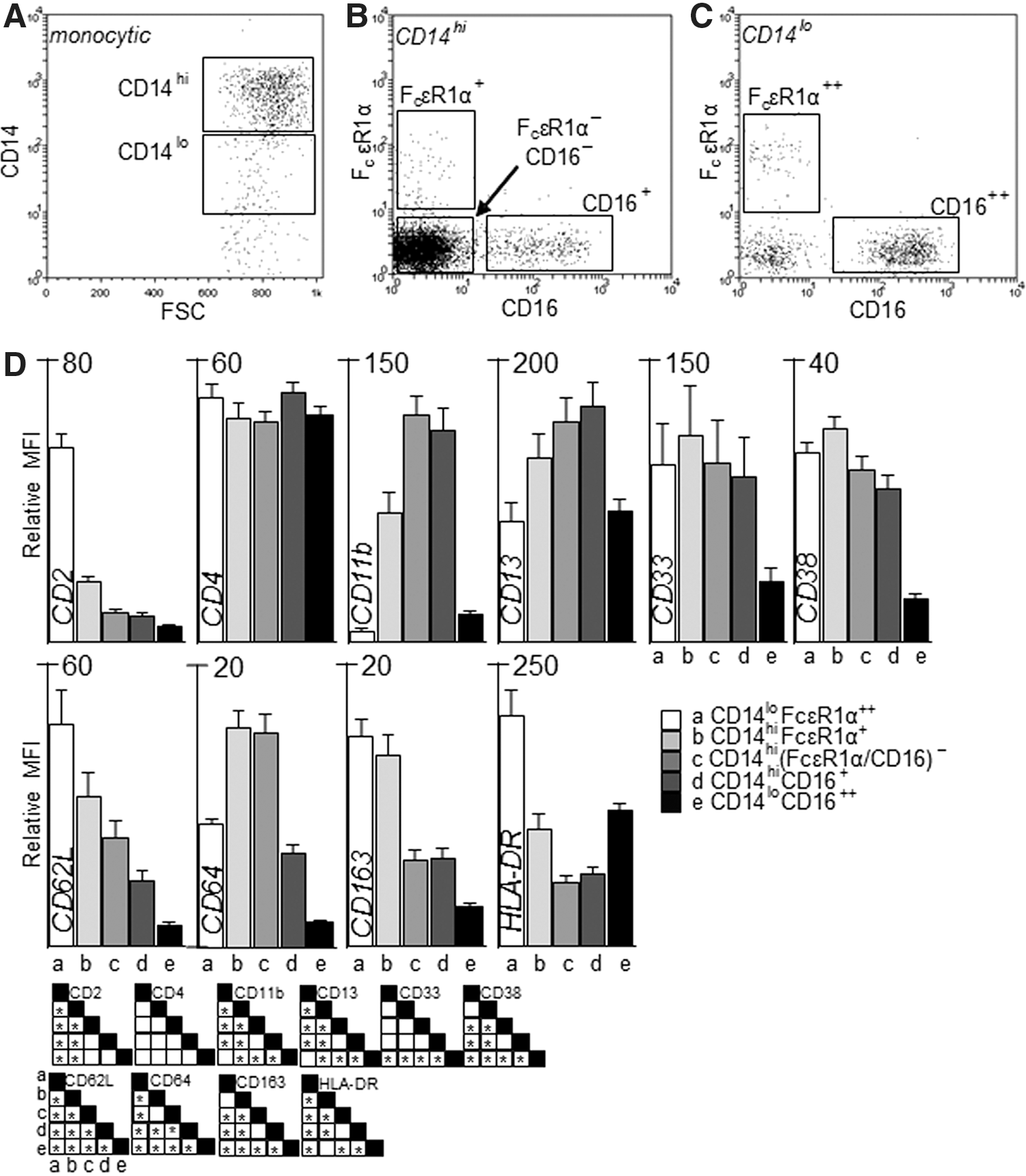
Five human monocyte subsets defined by the expression of CD14, FcɛRIα, and CD16. Within monocytic cells (based on FSC vs. SSC gating of PBMC), subsets were defined by the expression of
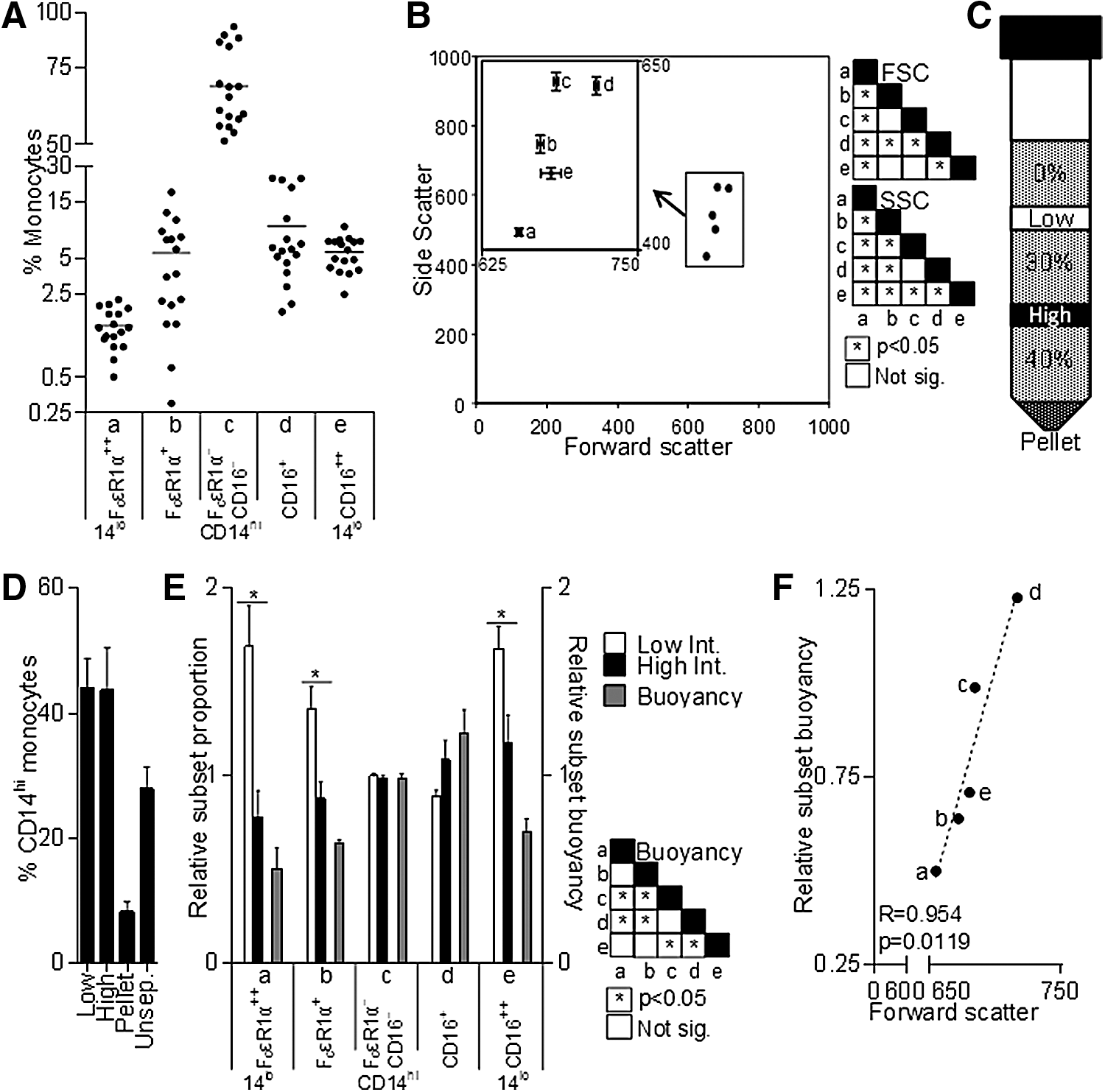
Prevalence, size, granularity, and buoyancy of monocyte subsets.
Size, granularity, and buoyancy
Early techniques for human monocyte separation, based on size and density [reviewed in Grage-Griebenow and others (2001a)], identified 2 subsets broadly defined as large and small, which appear to correlate with CD14hi and CD14loCD16++ monocytes, respectively (Ziegler-Heitbrock and others 1991). To determine differences in size and granularity (approximated by forward and size scatter, respectively) between the 5 subsets defined by the expression of CD14, CD16, and FcɛRIα, the flow cytometric scatter profiles of antibody-labeled PBMC were ascertained. Figure 3B illustrates the relative differences in mean forward and side scatter between the 5 subsets. The size and granularity of the CD14loCD16++ and CD14hiFcɛRIα+ subsets were similar, as were those of the CD14hi(FcɛRIα/CD16)− and CD14hiCD16+ subsets; CD14loFcɛRIα+ cells have the lowest size and granularity.
To determine the differences in buoyancy in the 5 human monocyte subsets defined by expression of CD14, CD16, and FcɛRIα, PBMC were layered over a 2-step discontinuous Percoll gradient and, after centrifugation, cells at each interface and the pellet were analyzed by flow cytometry (Materials and Methods, Fig. 3C). As seen in Fig. 3D the low- and high-density interfaces both had ∼43% CD14hi monocytes, whereas denser cells formed the pellet and averaged ∼8% CD14hi monocytes. The proportion of monocytes from each subset at both the high- or low-density interfaces was expressed relative to the respective proportion of each subset in unseparated PBMC for each donor, with no change being equal to 1 (Fig. 3E). The proportion of CD14hi(FcɛRIα/CD16)− monocytes in the high- and low-density fraction was equal to that in unseparated PBMC. In the high-density fraction, the relative proportion of each subset appears to increase as the expression of FcɛRIα decreased and CD16 increased. This pattern is inverted in the low-density fraction, with the exception of the CD14loCD16++ subset, which has a higher prevalence. To compare the differences in buoyancy between subsets, the high-density interface values were expressed relative to low-density interface values for each subset (Fig. 3E, see Supplementary Fig. S1 for further particulars). CD14hiCD16+ and CD14hi(FcɛRIα/CD16)− monocytes are significantly more buoyant than the other subsets using this analysis. There was a strong correlation between forward scatter and relative buoyancy for the 5 subsets (Fig. 3F), suggesting that each monocyte subset may represent a stage in the development from FcɛRIα+ monocytes to monocytes that lack FcɛRIα/CD16 and then to CD16+ monocytes, although the nonclassical subset does not appear to be an immediate continuation of the intermediate subset based on this analysis of physical properties.
PM prevalence is associated with high-affinity Fc receptor expression
CD64 expression on canonical monocyte subsets or the prevalence of a monocyte subset, as defined by FcɛRIα, CD16, and CD14, was measured to determine the association between these parameters and circulating PM. A proliferation assay was performed using CD14hi monocytes (Fig. 4A) and the relative expression of CD64 on each canonical subset at day 0 was correlated with the proportion of BrdU+ cells (Fig. 4B). There was an association between CD64 expression and PM prevalence for the classical subset and a weak trend for the intermediate subset; there appeared no correlation between PM prevalence and CD64 expression for nonclassical monocytes.
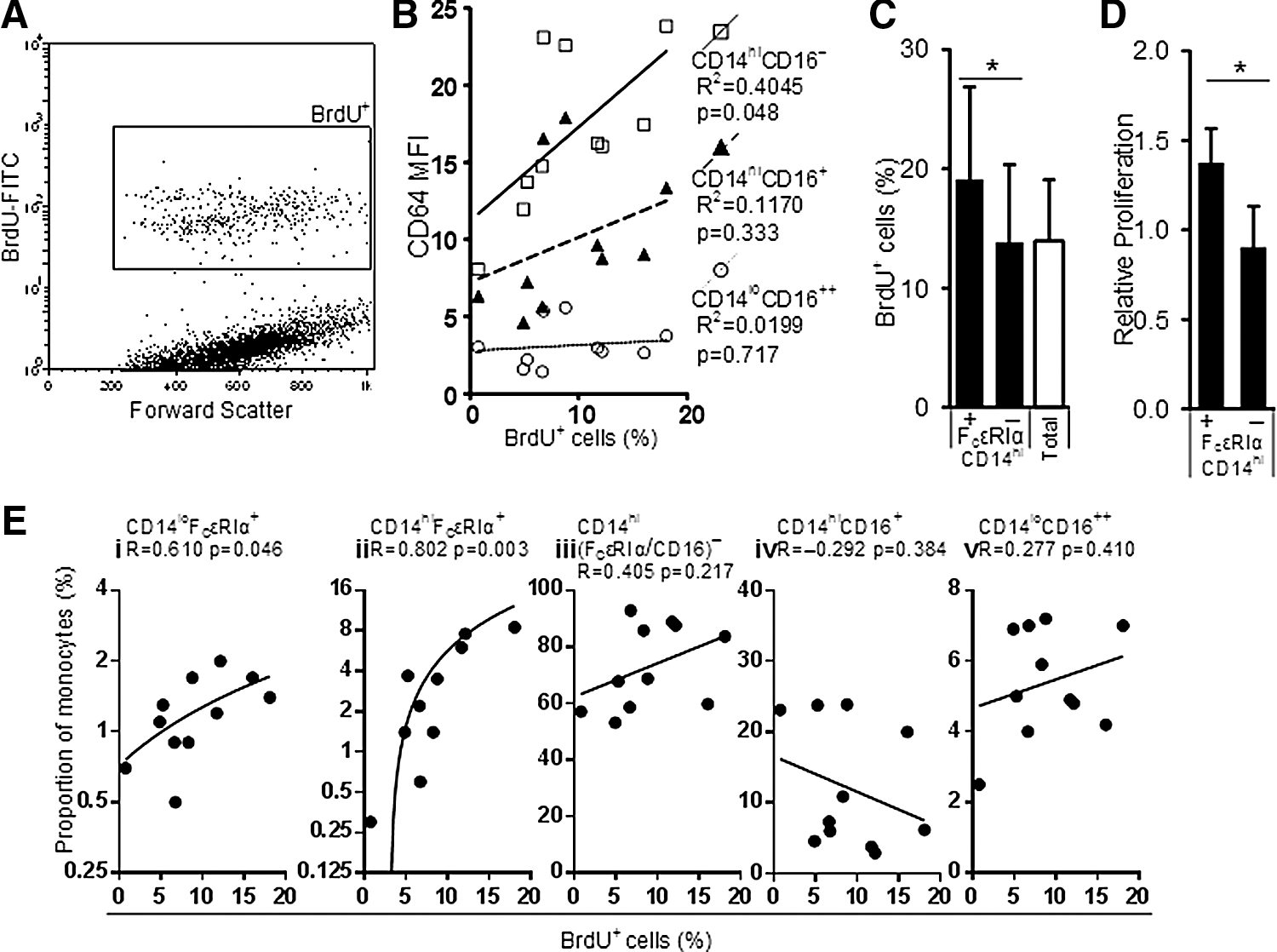
Circulating proliferative monocyte (PM) prevalence is associated with high-affinity FcR expression.
As CD14hiFcɛRIα+ monocytes share the same surface marker phenotype as PM (CD14hiCD11bloCD13loCD33hi CD64hi) (Clanchy and others 2006), it is unsurprising that proliferation, as measured by BrdU assay, was also significantly higher in cultures containing sorted CD14hiFcɛRIα+ monocytes, compared to sorted CD14hiFcɛRIα− monocytes (Fig. 4C, D). As shown in Fig. 4E, there was a significant positive correlation between the proportions of day 0 CD14hi/loFcɛRIα+ monocyte subsets and circulating PM, as determined by BrdU assay performed using the relatively abundant CD14hi monocytes sorted from each donor.
Monocyte subset CD64 expression is uniformly modulated by cytokine stimulation
A measurement of CD64 expression in different culture conditions was undertaken to determine whether the expression of CD64 on monocytes was upregulated due to surface marker modulation or an alteration in the composition of monocyte subsets. Previous studies of monocyte differentiation found a reduction in CD64 expression during culture (Chapuis and others 1997; Li and others 2009); however, the expression of CD64 on cells defined as canonical monocyte subsets has not been measured. In addition, the effects of cytokines that can modulate the expression of CD64 (Chapuis and others 1997; Li and others 2009) and PM activity (Cheung and Hamilton 1992; Clanchy and Hamilton 2012a), which have been investigated using total monocytes, were investigated within monocyte subsets over the same period.
The surface expression of CD64 was measured on canonical monocyte subsets before and after cytokine stimulation (Fig. 5). For each subset, the effect of individual cytokines was broadly similar after culture. IL-6 modestly increased CD64 expression, whereas M-CSF stimulation induced no change and GM-CSF slightly decreased the expression. In all subsets, the presence of IL-10 or IFN-γ significantly increased CD64 expression, compared to medium alone. The increase in CD64 expression in the presence of IFN-γ was most pronounced in the monocytes with an intermediate or nonclassical phenotype after culture, compared to the expression on these monocyte subsets before culture (Fig. 5D–F).
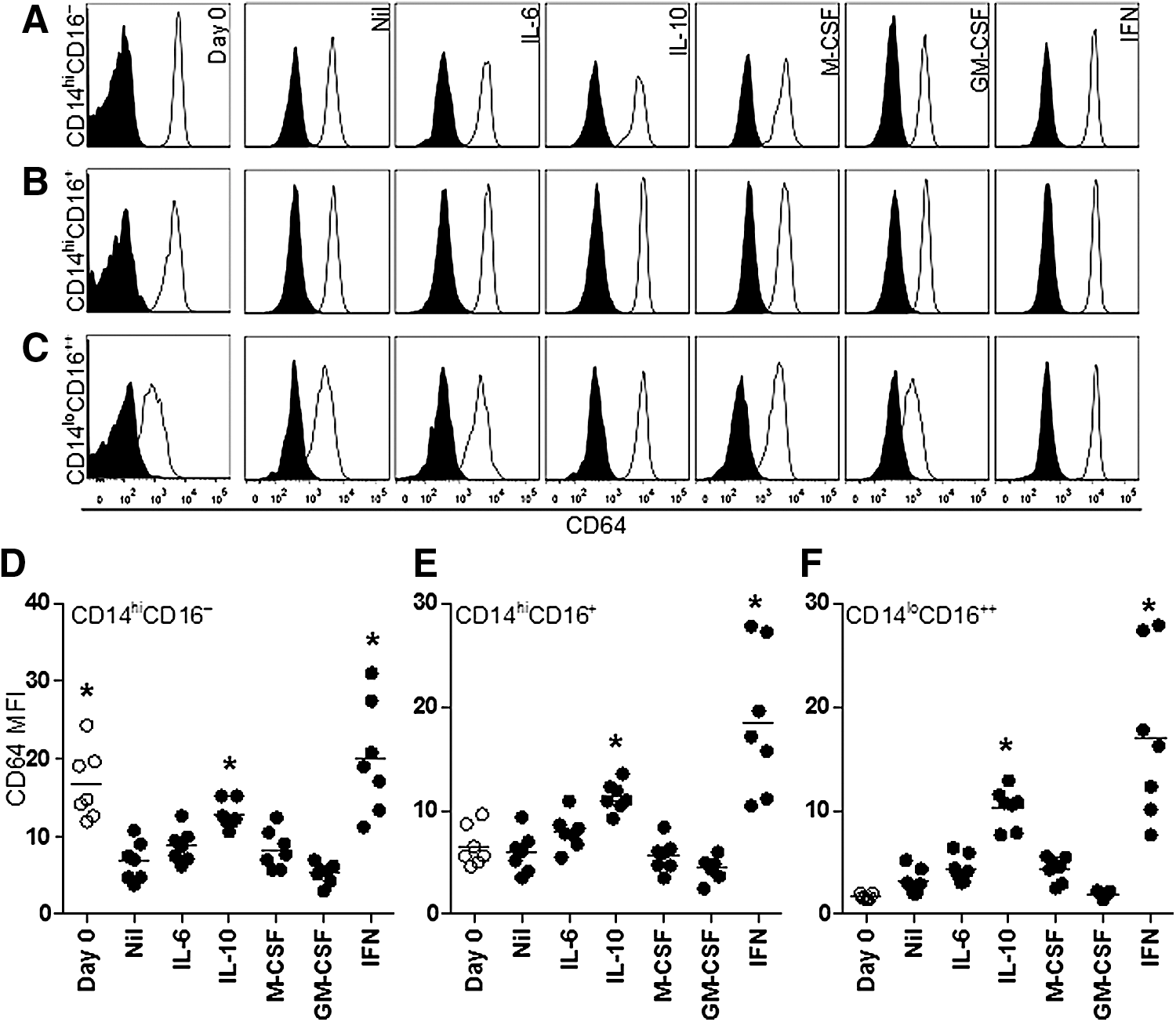
Monocyte subset CD64 expression is modulated uniformly after cytokine stimulation. Histograms detailing CD64 expression on monocytes in the
Cytokine stimulation modulates monocyte subset transition
The proportion of monocytes with the phenotype of a canonical monocyte subset was determined before and after cytokine stimulation (Fig. 6). Before culture, the classical monocytes were most prevalent and after culture the intermediate monocytes predominated; monocytes with the nonclassical phenotype were slightly reduced in proportion after culture, in the absence of cytokine stimulation. The presence of GM-CSF or IFN-γ significantly attenuated the reduction in classical monocytes and consequent increase in the proportion of intermediate monocytes. The presence of M-CSF, GM-CSF, or IFN-γ modestly elevated the proportion of nonclassical monocytes compared to medium alone, to a level similar to precultured cells. The apparent transition from classical to intermediate subset without a significant change in the nonclassical subset (Fig. 6E–G) was also observed in WBC stimulated with or without LPS (see Materials and Methods section). Based on these data, changes in the composition of monocyte subset phenotypes in the absence of cytokine stimulation indicate an increase in the proportion of the intermediate monocyte phenotype at the expense of classical monocytes.
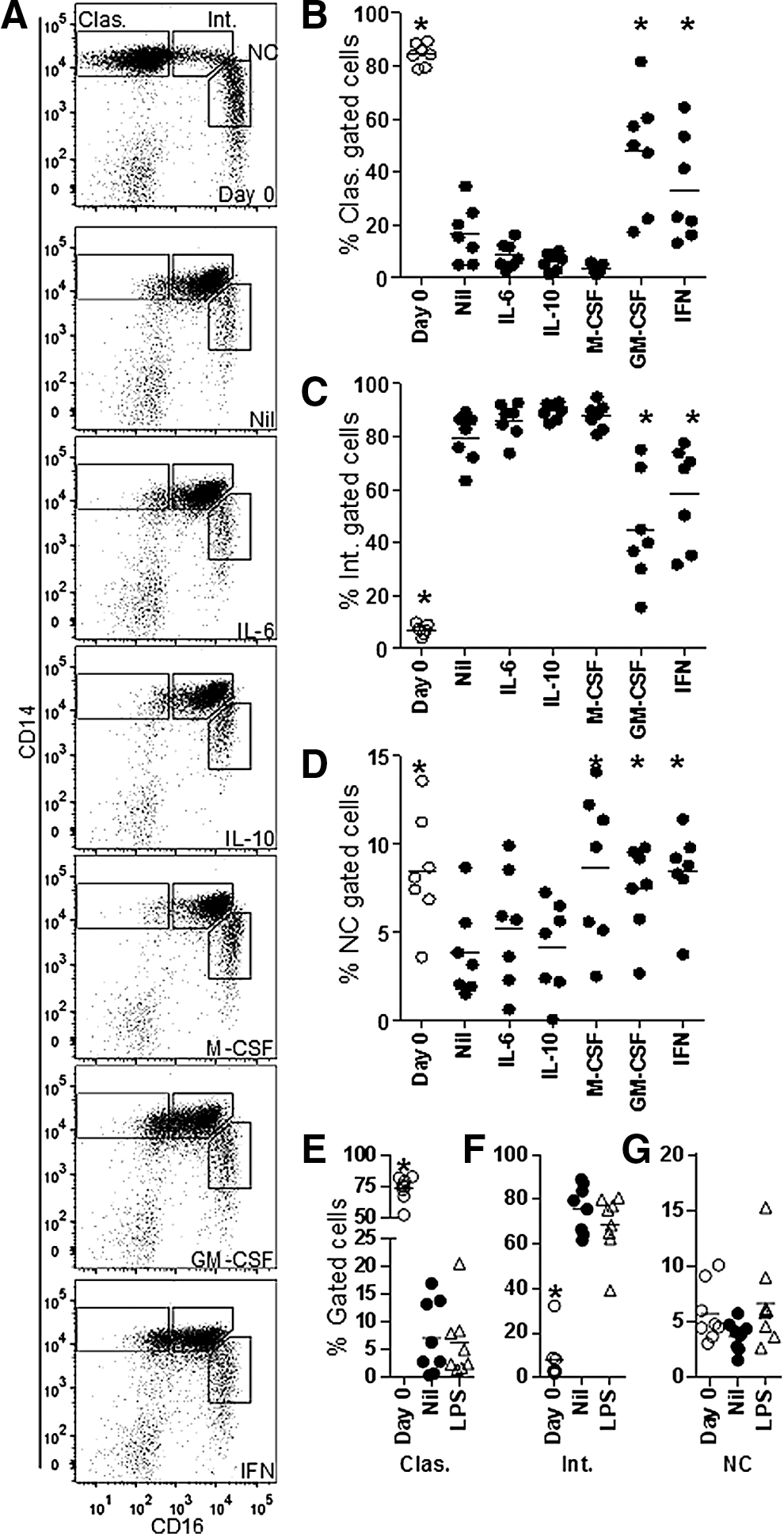
Monocyte subset transition is modulated by cytokine stimulation.
Loss of FcɛRIα precedes expression of CD16
To determine whether the 3 CD14hi subsets defined by expression of CD16 and FcɛRIα represent incremental steps in development, WBC were used to mimic maturation that occurs in the circulation in vivo (Skinner and others 2005). A comparison of freshly isolated PBMC and PBMC isolated from WBC shaken for 20 h indicates that on CD14hi monocytes (Fig. 7A, B), the expression of CD16 is upregulated (Fig. 7C); for the donors tested, the increase in fluorescent intensity was 6-fold higher (Fig. 7D). As indicated in Fig. 7E and F, there is a decrease in the proportion of CD14hiFcɛRIα+ monocytes and a concurrent increase in the proportion of CD14hiCD16+ monocytes between 0 and 20 h; of note is the increase in background staining and autofluorescence at 20 h (Fig. 7G), which was also observed in Fig. 7C. As detailed in Fig. 7H, the proportions of CD14hi monocytes in each quadrant (labeled α, β, γ, δ in Fig. 7E) indicate that the decrease in CD14hiFcɛRIα+ (α) and CD14hi(FcɛRIα/CD16)−(β) monocytes and increase in CD14hiCD16+ monocytes (γ) occurs without a transitional phase, where FcɛRIα and CD16 are coexpressed, as, in this in vitro model, the maturing cells do not transit through the δ quadrant. This aspect of the in vitro model reflects what is seen in freshly isolated monocytes, which also do not coexpress FcɛRIα and CD16.
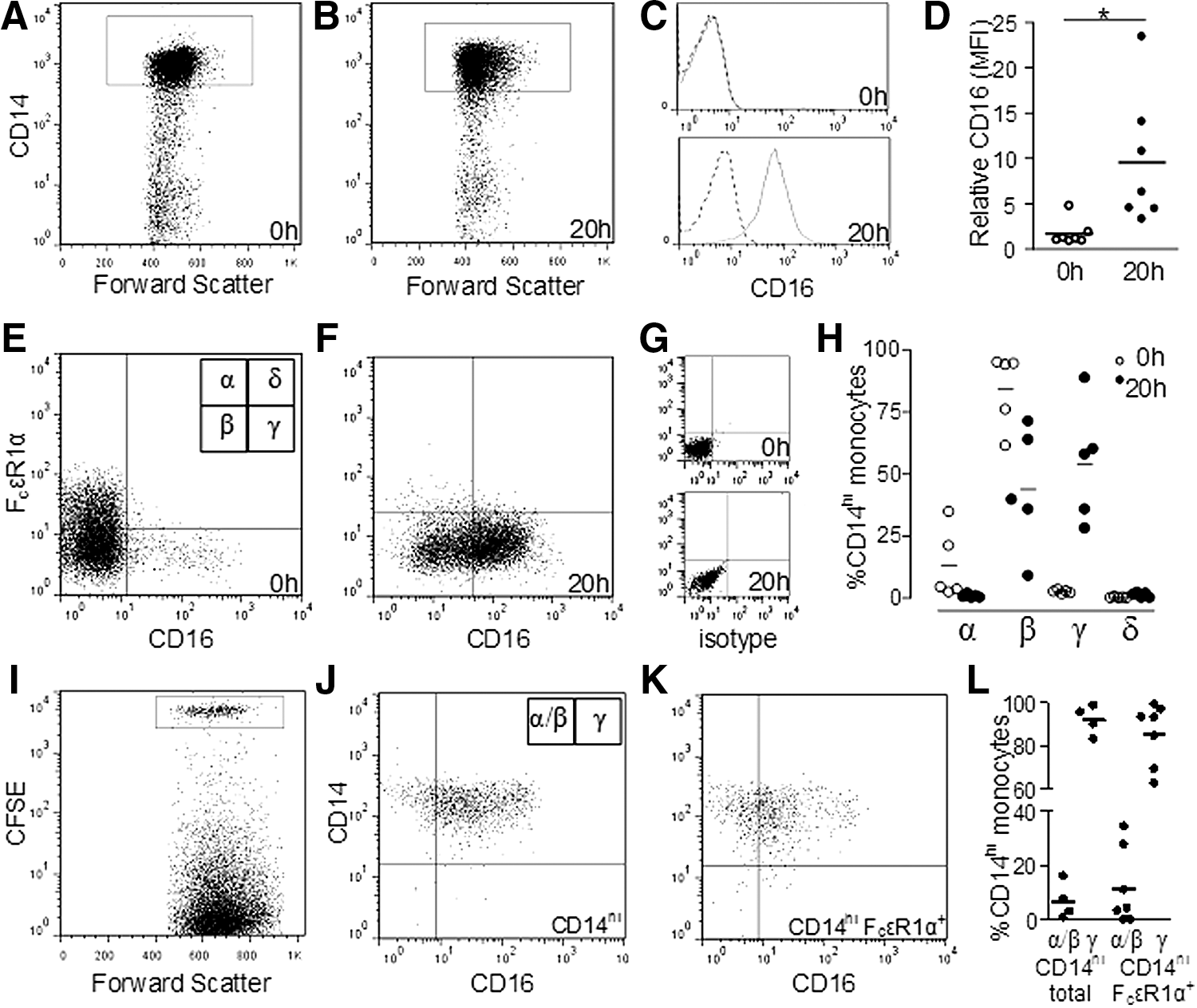
Loss of FcɛRIα precedes CD16 expression in WBC.
To confirm that the monocytes are losing FcɛRIα and gaining CD16, FACS-sorted monocytes (CD14hi or CD14hiFcɛRIα+) were labeled with CFSE and added to WBC. After short-term culture, the PBMC were then isolated as before. As expected, after gating on CFSE+ monocytes from the cultured blood, the sorted CD14hi monocytes expressed CD16 on most cells after culture; however, a large proportion of sorted monocytes that were originally CD14hiFcɛRIα+ also expressed CD16 after culture (Fig. 7J and K, respectively). Figure 7L details the proportions of CFSE+ CD16− and CD16+ monocytes (corresponding to quadrant labels α/β and γ in Fig. 7J).
Discussion
Surface markers delineated by the high-affinity Fc receptors CD64 or Fc ɛRIα
Several markers were differentially expressed on CD14hi CD64hi monocytes, although none was found solely or inordinately on this subset. The differences between surface profiles of monocyte subsets defined by high-affinity Fc receptor expression indicated a gradual change in monocyte phenotype from the classical to intermediate subset phenotype. Expression of CD16, along with CD14, is currently the main differentiator of human monocyte subsets and, as previously observed, CD64 expression was lowest on the CD14loCD16++ nonclassical subset (Grage-Griebenow and others 1993, 2001b). Within CD14hi monocytes, the pattern of expression for several markers (CD2, CD13, CD62L, CD38, and CD163) was suggestive of a maturational series, as there is a steady change in expression of these markers between the highly proliferative CD14hiCD64hi and the less proliferative CD14hi CD64lo (Fig. 1).
The correlation between CD64 expression and CD2 expression lead to the delineation of monocyte subsets based upon FcɛRIα, which is expressed on CD2+ monocytes (Cheng and others 2006). BDCA-1 (CD1c) was previously found to be expressed at low levels on CD14lo/hi FcɛRIα+ monocytes, whereas BDCA-2 and BDCA-3 was expressed only on CD14− cells (Cheng and others 2006). A small proportion of classical monocytes express FcɛRIα compared to CD64, which is expressed by the majority of classical monocytes. In contrast, the intermediate monocyte subset expressed no FcɛRIα and much lower levels of CD64. Based on the expression of CD14, CD16, and FcɛRIα, 5 monocyte subsets were defined. The following additional features of these subsets were analyzed: expression of common myeloid markers, size, granularity, and density.
Differences in cell surface markers demonstrated a particular profile for each subset and, on CD14hi monocytes, the cells expressing FcɛRIα or CD16 had the same changes in expression of CD2, CD13, CD62L, CD38, and CD163 as was observed for CD64 defined subsets. Of interest was the surface profile of CD14hiFcɛRIα+ monocyte subset as, relative to all of the CD14hi monocytes, this subset expressed the lowest amount of CD11b and CD13 and somewhat higher levels of CD33 and CD64; it has previously been shown that circulating PM has this cell surface phenotype—CD14hiCD11bloCD13loCD33hiCD64hi (Clanchy and others 2006).
The expression and function of CD62L is necessary for the sequestration of monocytes within the circulation (Xu and others 2008), and murine and human monocyte subsets that are most strongly associated with the endothelium have the lowest expression (Kamei and Carman 2010). The reduction in CD62L expression, concurrent with a reduction in CD64 expression, is consistent with a process of maturation before egress from the circulation. Likewise, Fig. 1B indicates a stepwise reduction in CD163 concordant with a reduction in CD64 expression. CD163 is present on cells of the monocyte/macrophage lineage, from progenitors through to terminally differentiated macrophages (Buechler and others 2000). While in the pig (Fairbairn and others 2013) and mouse (Bain and others 2013), higher CD163 expression is found on more mature monocytes, expression of CD163 on human monocytes is lowest on the intermediate and nonclassical subsets and comparatively higher on FcɛRIα+ monocytes.
HLA-DR was expressed in a bi-modal fashion, with the lowest expression on monocytes defined as CD14hiCD64med or CD14hi(FcɛRIα/CD16)−, but higher on the other subsets. An increase in CD74 expression in the intermediate subset (Zawada and others 2011), compared to the classical subset, may account for the increased surface expression of HLA-DR, due to greater trafficking to the cell surface through this chaperone (Borghese and Clanchy 2011). Monocytes which have increased potential to acquire a DC phenotype (Crawford and others 1999), are found in the subpopulation of monocytes that express CD2 at low to intermediate levels. Although CD2+ cells have been described as DC precursors based upon this differentiation potential, another interpretation of the data supporting this is that they are merely less mature monocytes with greater differentiation potential than CD2− monocytes. CD16− monocytes, which include CD2+ monocytes, have higher osteoclastic potential than CD16+ monocytes (Komano and others 2006), which uniformly lack CD2 (Fagnoni and others 2001), which suggests that CD2+ monocytes have higher differentiation potential rather than DC-specific potential.
When comparing subsets defined by CD64/CD14 or FcɛRIα/CD14/CD16, it is evident from the surface marker profiles that some CD14loFcɛRIα+ cells would be present in the CD14loCD64lo gate, indicating that the delineation of monocyte subsets by the expression of FcɛRIα, CD14, and CD16 achieves comparatively superior deconvolution of monocyte heterogeneity. The incremental changes in surface markers were paralleled by changes in physical properties. A putative transition of CD14lo/hiFcɛRIα+ monocytes to CD14hiCD16+ monocytes appears concurrent with an increase in size and granularity; the differences in buoyancy between subsets were similar and may influence the phenotype of monocytes isolated by elutriation or aphaeresis. A substantial overlap between one or more subsets in buoyancy, size, and granularity would strengthen the hypothesis that there is more than one monocyte lineage [potentially derived from different precursors (Hanna and others 2011)] maturing in the circulation. The nonclassical subset did not uniformly follow the trends of the other subsets in surface marker profile or physical properties, nor did it increase in proportion during short-term culture, indicating that it may not be immediately contiguous with the intermediate subset; other researchers have identified features that support a developmental link between these subsets (Wong and others 2011). By comparison, the physical properties and surface profile of the CD14loFcɛRIα+ monocyte subset appeared more congruous with a monocytic developmental continuum.
PM activity and high-affinity Fc defined subsets
The correlation of circulating PM prevalence and classical monocytes with the highest CD64 expression confirmed a previous finding of this subset's increased proliferative potential (Clanchy and others 2006), and provides further evidence that CD64 expression is indicative of monocyte maturity on CD14hi monocytes. The proportion of CD14hiFcɛRIα+ monocytes was more strongly associated with circulating PM prevalence. This is a refinement to the means by which circulating PM could be estimated, as the measurement of CD14hiFcɛRIα+ monocytes offers more precision than the relative expression of CD64 on the classical monocyte subset, compared to an isotype control. The use of an FACS-based metric for circulating PM also offers practical advantages over an 8 day proliferation assay that requires monocyte isolation, although further testing in the steady state and disease would be necessary to confirm this association.
Monocyte subset transition in vitro
The expression of CD64, the high-affinity receptor for IgG, has been used to classify monocytes with increased antiviral and antigen presentation potential (Grage-Griebenow and others 1993, 2001b); monocyte subsets defined by CD64 also have differences in phagocytosis and the generation of reactive oxygen species (Grage-Griebenow and others 2000). An increase in CD64 expression on monocytes has been measured in autoimmune disease (Li and others 2009), and has also been suggested as indicative of enhanced innate immunity during pregnancy (Neuteboom and others 2011). The increase in CD64 expression during disease may be due, in part, to increased concentrations of circulating IFN-γ, as this cytokine is known to upregulate CD64 expression (Li and others 2009). Increased circulating levels of IFN-γ have been associated with increased CD64 expression in AIDS (Capsoni and others 1994) and the effects of IFN-γ, IL-6, and IL-10 on total monocyte expression of CD64 have been evaluated in vitro (Chapuis and others 1997; Li and others 2009), with broadly similar effects to those described here.
In this study, 2 short-term culture systems were used to measure monocyte subset transition from the classical to the intermediate subset. An increase in nonclassical monocytes has been reported in WBC (Skinner and others 2005); however, this was not observed in the current study, despite the presence of a toll-like receptor ligand (Fig. 6E–G). The presence of GM-CSF and IFN-γ appeared to attenuate the transition from classical to intermediate subset compared to cells cultured in medium alone. Although the changes in monocyte phenotype in response to IFN-γ and other mediators have been reported (Firestein and Zvaifler 1987; Jayaram and others 1989; Buckle and others 1990), changes in phenotype with regard to monocyte subsets have not been adequately detailed.
CD64 expression on each canonical subset differed after culture according to the presence of each cytokine. It was determined that GM-CSF and IFN-γ caused an attenuation of monocyte subset transition, although IFN-γ (and IL-10, to lesser extent) appeared to maintain or increase CD64 expression on monocytes with a classical subset phenotype after culture, whereas GM-CSF did not. After culture, monocytes with the intermediate and nonclassical subset phenotype had increased expression of CD64 in the presence of IFN-γ and IL-10. Stimulation with IFN-γ (and to a lesser extent IL-10) permitted supraphysiological CD64 expression on the monocytes with a CD16+ intermediate and nonclassical subset phenotype after culture.
Monocytes with a classical subset phenotype after culture in medium alone express CD64 at a level that is equivalent to circulating intermediate monocytes (Fig. 5E), whereas intermediate monocyte CD64 expression was unchanged by culture in medium alone. The transition from classical to intermediate monocyte phenotype was also observed in WBC without coexpression of FcɛRIα and CD16; a definitive change was also observed using sorted CD14hiFcɛRIα+ monocytes, which became CD16+. Although monocyte subsets in culture appear to progress from the classical subset to the intermediate subset, further analyses will be required to confirm whether freshly isolated intermediate subset monocytes are phenotypically similar to those derived in vitro, as has been demonstrated by other studies (Calzada-Wack and others 1996; Randolph and others 2002). Nevertheless, these data suggest, for classical monocytes that the reduction in the expression of the high-affinity Fc receptors CD64 and FcɛRIα is a condition precedent to subset transition in the steady state.
Conclusion
It was determined that several key myeloid surface markers are coexpressed at similar levels depending on CD64 expression. Five subsets defined by the expression of CD14, FcɛRIα, and CD16 were characterized and found to differ in surface phenotype and physical properties (size, granularity, and buoyancy). The expression of CD64 on the classical subset and the prevalence of the CD14hiFcɛRIα+ monocyte subset correlated with the prevalence of circulating PM. These findings indicate that the decline in the expression of high-affinity Fc receptors on monocytes is accompanied by changes in cell surface expression and a degree of maturation, which is evidenced by a reduction in proliferative potential. Further characterization of FcɛRIα+ monocytes may reveal a mechanism for increased proliferative potential. Although these data indicate that FcɛRIα+ monocytes are antecedent to the canonical classical and intermediate monocyte subsets, further functional analyses are required to determine whether these “preclassical” monocytes require separate classification within the current nomenclature.
Authorship
F.I.L.C. conceived, designed, and performed research, analyzed data, and wrote the article.
Footnotes
Acknowledgments
The author thanks A. Holloway, D. Hewish, and C. McIssacc for technical assistance and Prof. J.A. Hamilton for helpful comments on the article. This work was supported by the Cooperative Research Center for Chronic Inflammatory Diseases.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.