
Abstract
Interferon beta (IFNβ) is used as a first-line treatment for multiple sclerosis (MS) and is injected intramuscularly or subcutaneously (s.c.). The subcutaneous route is considered more immunogenic as it is associated with increased antidrug antibody-positive patients. The skin contains dendritic cells (DCs) and it is unclear whether these contribute to immunogenicity. To assess the effect of IFNβ on skin-resident cells, IFNβ was injected intradermally (i.d.) ex vivo using a human skin explant model or s.c. in vivo in MS patients. Ex vivo, intradermal IFNβ injections reduced migration and enhanced surface CD86 expression of dermal DCs, and an increased expression of HLA-DR+ was observed in skin biopsies taken after subcutaneous IFNβ injection (in vivo). In both models, IFNβ elevated the expression of several inflammatory cytokines when compared to the control biopsies. Our results show that 3 different IFNβ preparations, normalized in dose and injection site, induce similar immune responses, suggesting that the differences in immunogenicity are likely due to the route and frequency of administration.
Introduction
Recombinant interferon beta (IFNβ) preparations are used as an initial treatment regime for relapsing remitting multiple sclerosis (RRMS), a disease of the central nervous system. Two types of recombinant IFNβ preparations are available for the treatment of multiple sclerosis (MS): IFNβ-1a (Avonex®/Rebif®/Plegridy®) and IFNβ-1b (Betaferon®/Extavia®). The differences between these preparations lie in their amino acid sequence (Haji Abdolvahab and others 2016) and glycosylation pattern. IFNβ-1a is produced in Chinese hamster ovary cells (Secondary Progressive Efficacy Clinical Trial of Recombinant Interferon-Beta-1a in MSSG 2001) and has the same amino acid sequence and glycosylation structure as the endogenous protein (Radhakrishnan and others 1996). IFNβ-1a is administrated either intramuscularly (i.m.) as 6 million units (30 μg) once a week (Avonex) or subcutaneously (s.c.) with a dose escalation from 2.4, 6, to 12 million units (8.8, 22, or 44 μg) thrice a week (Rebif). The most recently approved IFNβ-1a formulation is PEGylated (Plegridy) and is administrated s.c. as 12 million units (125 μg) every 2 weeks. IFNβ-1b is produced in Escherichia coli (Knobler and others 1993) and is therefore not glycosylated, and has amino acid modifications to increase the yield (Karpusas and others 1998; Runkel and others 1998; van Beers and others 2010). IFNβ-1b (Betaferon/Extavia) is administrated s.c. as 8 million units (250 μg) every other day.
In some individuals, administration of IFNβ results in a break of immune tolerance leading to a humoral response against the protein and development of neutralizing antidrug antibodies (NAb) (Group 1993; Tovey and Lallemand 2011). The efficacy of IFNβ is adversely affected as NAb develop by reduction or abrogation of the bioavailability of the drug (Polman and others 2010), and therefore negatively affect clinical outcomes (van der Voort and others 2009). The frequency of NAb positivity varies between different IFNβ products and develops in up to 47% of IFNβ-1b recipients: in 6% for i.m. administered IFNβ-1a and in 28% for s.c. delivered IFNβ-1a (Deisenhammer 2009). Factors that influence the immunogenicity of IFNβ are both patient and drug related, including the molecular structure of the drug, as well as the frequency of administration and the injection route. The subcutaneous route has been demonstrated to be more immunogenic than the intramuscular route (Schellekens 2002; Deisenhammer 2009).
The skin is an important immunological barrier and provides local immunity by complex cell networks, including epidermal Langerhans cells (LCs) and dermal dendritic cells (DCs). Several subsets of DCs are resident in the dermis, including CD1a+ DCs and CD14+ DCs. Once the LCs and dermal DCs are activated, they will migrate to the peripheral lymph nodes where the LCs and CD1a+ DCs stimulate T cells to become efficient helpers for the activation of cytotoxic T cell responses (Nestle and others 1994; Yuan and others 2005; van der Vlist and others 2011). Dermal CD14+ DCs are more prone to prime CD4+ T cells to the follicular helper T cell phenotype (Klechevsky and others 2008), and a study by Yao and others (2015) showed that presentation of foreign antigen to LCs also promoted an expansion of follicular helper T cells. The follicular helper T cells can promote the differentiation of activated B cells into immunoglobulin (Ig)-secreting plasma cells and induce naive B cells to switch isotype toward IgG and IgA (Klechevsky 2013), and it has been suggested that targeting dermal CD14+ DCs could induce a strong antibody response (Klechevsky and others 2008; Ueno and others 2010). Indeed, cutaneous vaccine delivery to target skin-resident antigen-presenting cells is associated with a greater immune response compared to other delivery routes (Romani and others 2012; Klechevsky 2013; Smed-Sorensen and Lore 2013). The mechanisms of induction of immunological responses against biological drugs administered through the skin are still relatively unexplored.
In this study, we monitored migration status of skin cells and surrounding matrix after administration with IFNβ preparations in an ex vivo skin explant model (Schneider and others 2012; Bakdash and others 2013), and after subcutaneous IFNβ administration to people with MS (in vivo). To the best of our knowledge, this is the first study investigating the local biological effect of IFNβ administration using a human ex vivo skin model.
Materials and Methods
Ethics
The research study was approved by the regional ethics board in Stockholm, Sweden, and Innsbruck, Austria. Written informed consent was obtained from all participating patients.
Human skin explants
Human skin specimens were obtained from abdominal reduction surgery. The skin was kept at 4°C before processing (between 3 to 12 h postsurgery). The skin was injected intradermally (i.d.) with 50 μL of 105 or 102 U/mL of IFNβ-1a (Biogen Idec, Inc. and Merck Serono) or IFNβ-1b (Bayer) diluted in phosphate-buffered saline (PBS), or 50 μL PBS alone, which was used as a control. Insulin needles (0.6 × 25 mm Microlance; BD Biosciences) were used for the injections placed at a 5- to 15-degree angle. The technique allows for a bollow to identify the exact location of the intradermal injection. A 6 mm biopsy (Kai Europe GmbH, Stille) was sampled directly after each intradermal injection. As a washing step, each biopsy was transferred with the epidermal side up to 0.5 mL Iscove's Modified Dulbecco's Medium (Lonza) containing 1% fetal bovine serum (FBS) (Thermo Scientific) and 0.086 mg/mL gentamicin (Sigma) in a 48-well plate (Costar; Corning Life Sciences) for 1 h in room temperature. Then, the biopsies were transferred with the epidermal side up to a new 48-well plate (Costar; Corning Life Sciences) containing 1 mL Iscove's Modified Dulbecco's Medium (Lonza) supplemented with 10% FBS (Thermo Scientific), 500 U/mL of recombinant human granulocyte-macrophage colony-stimulating factor (PeproTech), and 0.086 mg/mL gentamicin (Sigma), and cultured at 37°C, 5% carbon dioxide (CO2). After 3 days, the biopsies were discarded, and the medium containing migrating crawl-out cells was harvested and pooled for each test condition, and a hemocytometer was used for cell counting. Some biopsies were collected for immunohistochemistry analysis after 90 min and 24 h after the intradermal IFNβ-1a (Rebif; Merck Serono) (105 U/mL) or PBS injections, or at an uninjected site.
Phenotypical analysis of crawl-out cells
Cells that migrated from the skin biopsies were stained with fluorescent-conjugated antibodies in PBS, 1% bovine serum albumin (Sigma), 2% FBS (Thermo Scientific), and 1% human serum (Invitrogen) for 30 min at 4°C. To identify the different skin DC subsets, the following monoclonal mouse anti-human antibodies were used: HLA-DR, CD1a, CD14 (BD Biosciences), CD11c (eBioscience), and langerin (Beckman Coulter). CD86 was used as maturation marker (BD Biosciences). The samples were acquired by CyAn Dako flow cytometer (Beckman Coulter) and FlowJo software version 7 (Tree Star, Inc.) was used for the analysis.
Isolation of naive CD4+ T cells
Human peripheral blood mononuclear cells from 1 healthy donor were isolated from buffy coat by Ficoll-Paque (GE HealthCare) density gradient centrifugation. As a first separation step, naive CD4+ T cells were isolated by negative magnetic separation with the autoMACS Pro Separator (Miltenyi Biotec) using the Naive CD4+ T cell isolation kit II (Miltenyi Biotec) according to the manufacturer's instructions. As a second step, the collected naive CD4+ T cells were further cell sorted by flow cytometry (MoFlo™ XDP; Beckman Coulter). The cells were stained for 30 min at 4°C with monoclonal mouse anti-human fluorescent-conjugated antibodies CD45RA and CD4 (BD Biosciences). The sorted cells reached a purity of 99% and were cryopreserved until use.
Stimulation of naive CD4+ T cells
The total population of migrated DCs was irradiated with 25 Gy of gamma radiation to avoid potential cross-reactivity of T cells in the skin. The DCs were then washed and immediately cocultured (4 × 104 cells/well) with allogeneic naive CD4+ T cells (4 × 104 cells/well) in a total volume of 200 μL IMDM supplemented with 10% FBS in a 96-well flat-bottom plate and incubated at 37°C, 5% CO2. As controls, naive CD4+ T cells and DCs were cultured separately. T cell proliferation was measured after 5 to 6 days by ATPlite (PerkinElmer) according to the manufacturer's protocol using GloMax® 96 microplate luminometer (Promega).
Multiple sclerosis patients
Two females and 1 male with RRMS took part in the skin biopsy study. The patients had no history of other diseases. All patients were treatment naive to IFNβ when included in this study. Biopsy samples from the MS patients were obtained at 2 time points. At the baseline study visit, 2 patients received their first subcutaneous injection containing 8.8 or 44 μg IFNβ-1a (Rebif), and 1 patient had been on IFNβ treatment for 1 month and received 44 μg. The drug was given in the abdomen or thigh and the injection site was marked with a pen. After 4 to 5 h, following local anesthesia, one 4 mm punch biopsy was taken from the subcutaneous IFNβ-1a injection site (marked with a pen) and 1 biopsy from the collateral site of the body at an uninjected skin site. After 11 to 12 months on full-dose (44 μg) IFNβ-1a (Rebif) treatment, additional 4 mm punch biopsies were sampled at the subcutaneous IFNβ-1a injection site and 1 on the collateral site of the body at an uninjected skin site 4 h and 40 min to 18 h after receiving the treatment. A blood sample was collected simultaneously for NAb analysis. Potential injection site reactions were assessed.
Immunohistochemical analysis and image analysis
For both the MS patients and the skin explant model, skin punch biopsies were mounted in 15 × 15 × 5 mm cryomolds (Sakura, Tissue-Tek) with optimum cutting temperature (O.C.T.) (Sakura, Tissue-Tek) and frozen on dry ice before being stored at −80°C. Cryosections of 8 μm thickness were prepared on slides (Superfrost Plus, Menzel-Gläser) and fixated in 2% paraformaldehyde in PBS. Sections were stained at 4°C overnight with rabbit monoclonal HMGB1, rabbit polyclonal IL-1β, IL-6, COX-2, and PGE2 (Abcam), and mouse monoclonal TLR4 (Abcam), HLA-DR (BD Biosciences, L243), and MxA (Flohr and others 1999) (Professor Haller and Kochs, University of Freiburg, Germany, M143). Appropriate isotype-specific control antibodies were included to determine specificity (Abcam). Biotinylated rabbit anti-mouse (Dako) or goat-anti-rabbit (Dako) was used as secondary antibodies. To label the secondary antibodies, Vectastain Elite ABC kit together with substrate diaminobenzidine tetrahydrochloride (Vector Laboratories) were used. Endogenous peroxidase and avidin/biotin were blocked with 3% hydrogen peroxidase (Sigma-Aldrich) and avidin/biotin blocking kit (Vector Laboratories), respectively. Rabbit and goat serum (Sigma-Aldrich) was applied to block before application of the secondary antibodies. Hematoxylin (1:1 in PBS) (Mayers, Histolab) staining was performed to define the skin structure. All scale bars represent 100 μm. 3,3′-Diaminobenzidine staining was evaluated by optical inspection and Fiji Software (Schindelin and others 2012). Rating performed were I = no staining/weak, II = moderate, and III = strong.
Statistical analysis
Data are presented as the mean and standard deviation. Statistical calculations were performed with GraphPad Prism 6. Shapiro–Wilk normality test was used and since not all data groups were normally distributed, we choose to only use nonparametric paired Wilcoxon test for analysis. Significance was defined as a P value <0.05.
Results
IFNβ reduces the migration of tissue-resident DCs
To evaluate the impact of IFNβ administration through the skin, we investigated the migration pattern of skin-resident DCs after intradermal injection with IFNβ-1a (i.m. or s.c.) and IFNβ-1b (s.c.) compared to only injecting PBS by flow cytometry using 10 skin donors (ex vivo). Absolute cell counts with hemocytometer showed that treatment with the high-dose IFNβ (105) resulted in lower numbers of cells that migrated out of the tissue biopsies compared to the low-dose IFNβ (102) and PBS (Fig. 1a). Migration of 3 major skin DC subsets was investigated; LCs (CD207+), CD1a+ dermal DCs (DDCs), and CD14+ DDC. DC subsets were identified by their forward and side scatter (FSC/SSC) properties followed by expression of HLA-DR+ and CD11c+ (parent gate) (Fig. 1b), and categorized according to their expression of HLA-DR+/CD11c+/CD1a+/CD207+, HLA-DR+/CD11c+/CD1a+, and HLA-DR+/CD11c+/CD14+ (see Fig. 1c for a representative sample). The high-dose IFNβ (105 U/mL) significantly reduced the migration of HLA-DR+/CD11c+/CD14+ DDCs compared to the PBS control (IFNβ-1a [i.m.] P = 0.0039; IFNβ-1b [s.c.] P = 0.0020; and IFNβ-1a [s.c.] P = 0.0020) (Fig. 1d). HLA-DR+/CD11c+/CD1a+ DDCs first showed a trend that the high-dose IFNβ (105 U/mL) induced the migration compared to the PBS control, but the finding was not significant when replicated (Fig. 1e). HLA-DR+/CD11c+/CD1a+/CD207+ LC had significantly reduced migration pattern when encountered with the high-dose IFNβ (105 U/mL) compared to the PBS control (s.c. IFNβ-1b P = 0.0156 and s.c. IFNβ-1a P = 0.0078) (Fig. 1f).
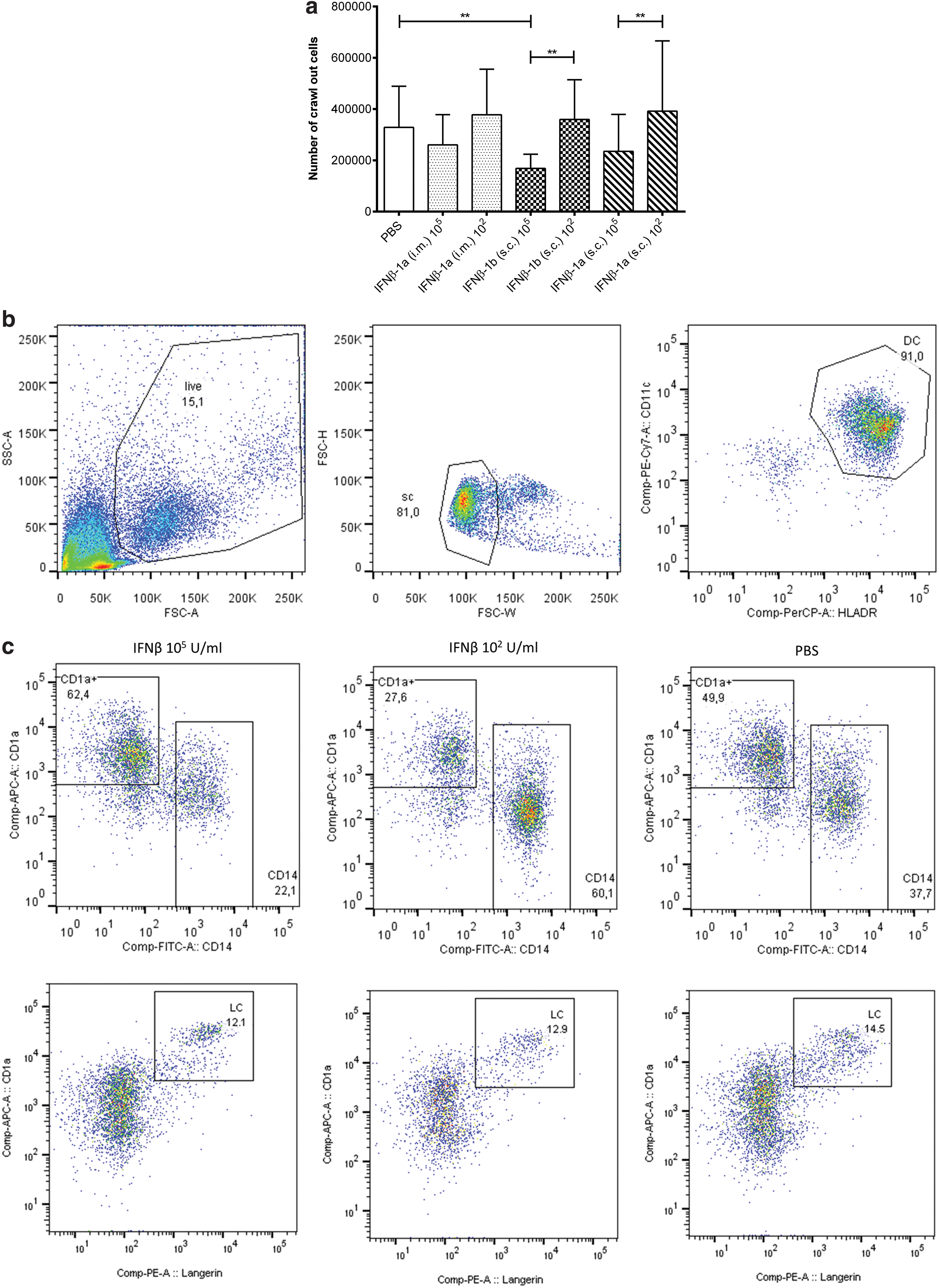
Human skin punch biopsies were obtained after intradermal injection of IFNβ-1a (i.m.), IFNβ-1a (s.c.), IFNβ-1b (s.c.), or PBS followed by a 3-day culture.
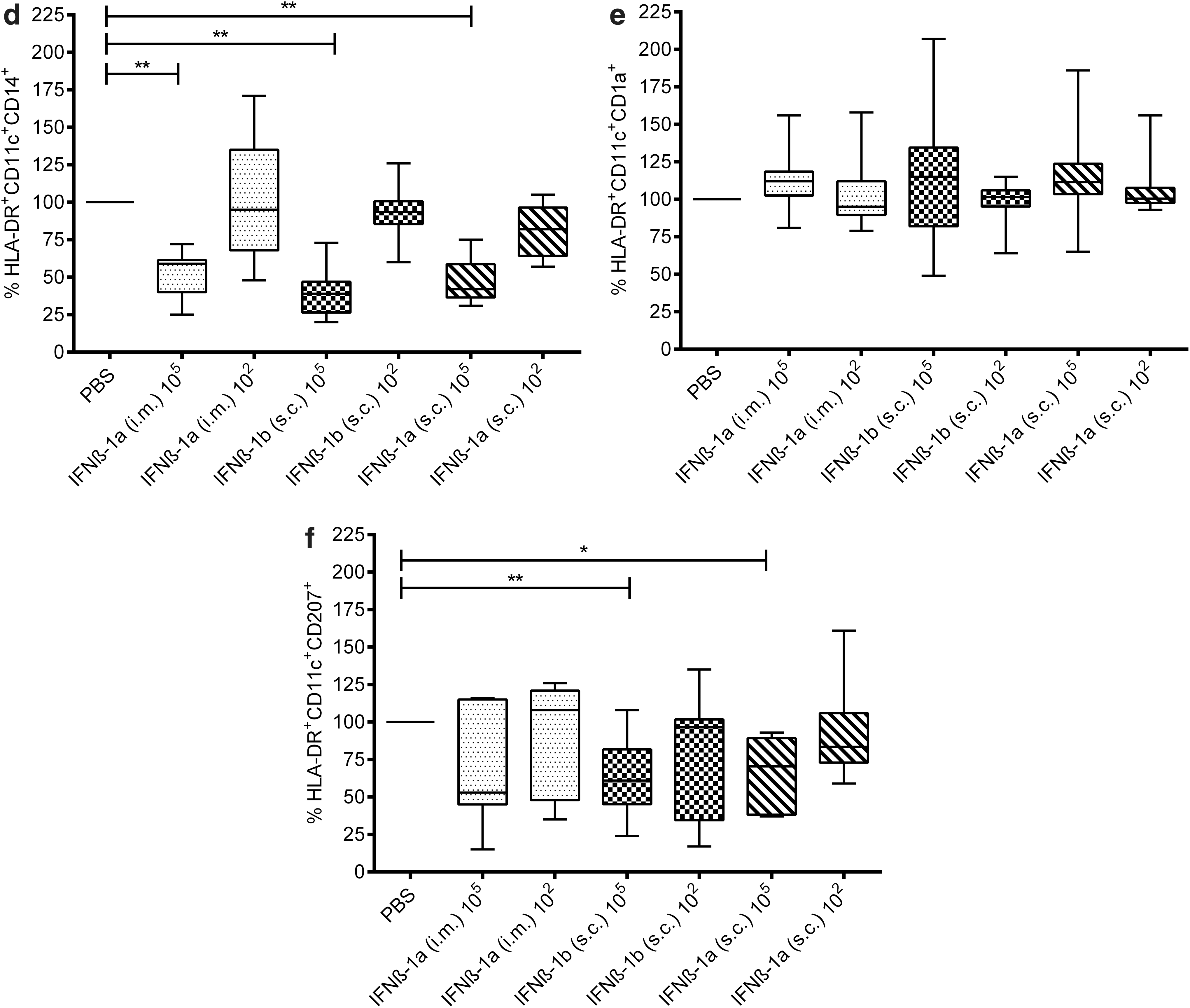
IFNβ upregulates cell surface expression of CD86
Next, we investigated whether the IFNβ-treated migratory DCs had an affected maturation phenotype. This was tested by the expression of the activation marker CD86 by flow cytometry using 10 skin donors (ex vivo). DC activation was phenotypically assessed on 2 DDC subsets: HLA-DR+/CD11c+/CD1a+ and HLA-DR+/CD11c+/CD14+. DC subsets were identified by their FSC/SSC properties followed by expression of HLA-DR+ and CD11c+ (parent gate) and categorized according to their expression of HLA-DR+/CD11c+/CD1a+/CD86+ and HLA-DR+/CD11c+/CD14+/CD86+. Mean fluorescent intensity histogram plots show that the expression of CD86 was shifted after the high-dose IFNβ compared to PBS injections (Fig. 2a). Furthermore, geometric means of CD86 expression were used to quantify fluorescence intensity. Compared to PBS, the high dose of IFNβ (105 U/mL) significantly upregulated the cell surface expression of CD86 on HLA-DR+/CD11c+/CD14+ DDCs (i.m. IFNβ-1a P = 0.0234; s.c. IFNβ-1b P = 0.0020; and s.c. IFNβ-1a P = 0.0039) (Fig. 2b) and HLA-DR+/CD11c+/CD1a+ DDCs (i.m. IFNβ-1a P = 0.0430; s.c. IFNβ-1b P = 0.0020; and s.c. IFNβ-1a P = 0.0059) (Fig. 2c).
Maturation of the crawl-out DCs was investigated for the expression of CD86 following the injection of IFNβ-1a (i.m), IFNβ-1a (s.c), IFNβ-1b (s.c), or PBS.
IFNβ-treated DCs reduced the proliferation capacity of naive CD4+ T cells
Since the IFNβ-treated dermal DCs (ex vivo) had a more cell surface expression of CD86 than PBS-treated DCs, it was of interest to further investigate DC function by measuring their stimulatory capacity of naive CD4+ T cells using 6 skin donors (ex vivo). Overall, intradermal administration of IFNβ hampered the ability of the crawl-out DDCs to promote proliferation of allogenic naive CD4+ T cells compared to PBS-treated DCs (P = 0.0313) (Fig. 3) when using luminescence detection of intracellular adenosine triphosphate as a measure of cell numbers.
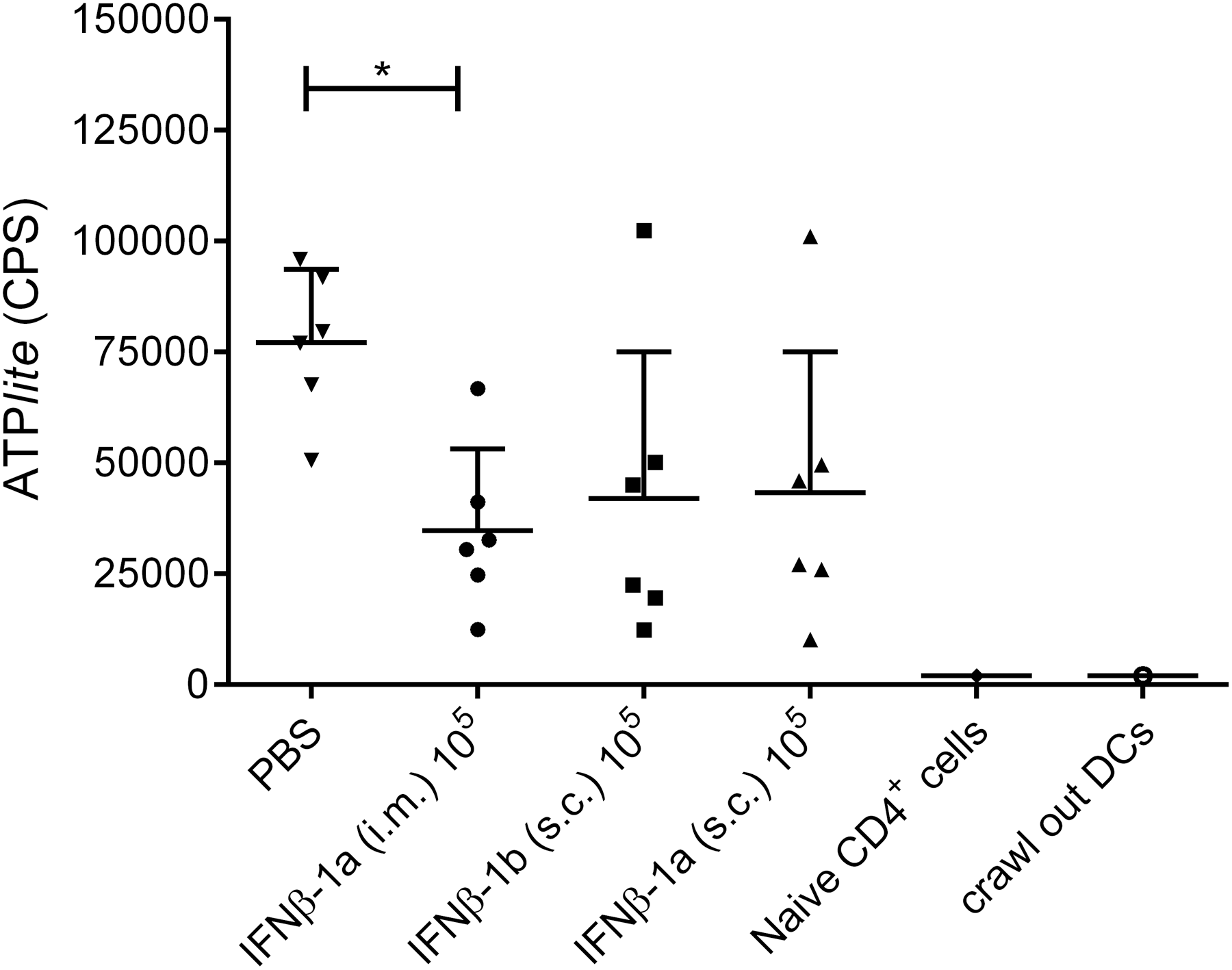
Intradermal injection of the high-dose (105) IFNβ reduced the induced T cell stimulatory capacity of the skin crawl-out DCs. The proliferation of allogenic naive CD4+ T cells stimulated with crawl-out cells derived from IFNβ injected skin was determined by ATPlite. Data are representative of six independent experiments. *P ≤ 0.05.
Inflammatory marker expression is elevated in biopsies after IFNβ injection ex vivo
To determine the impact of the IFNβ injection at the skin site, immunohistochemical staining for different inflammatory markers was performed using the human ex vivo skin model using 4 donors. Microscopic examination showed elevated expression of HMGB1, IL-1β, IL-6, and COX-2 as seen by increased stained area in the epidermis and dermis in IFNβ-injected skin compared to the saline control and the uninjected biopsies sampled after 90 min, and PGE2 and TLR4 had similar staining intensity in IFNβ-injected skin and the saline control site. The uninjected sites showed negative or week staining (Fig. 4a) (Supplementary Figs. S1 and S2). After 24 h, the IFNβ-injected biopsies showed stronger expression of HMGB1, IL-6, COX-2, PGE2, and TLR4 compared to saline and uninjected biopsies. The uninjected sites showed continued weaker staining compared to IFNβ and saline (Fig. 4b) (Supplementary Figs. S1 and S2). MxA was used as a positive control for IFNβ and was increased first after 24 h after the IFNβ injection compared to the saline and uninjected biopsies (Fig. 4c) (Supplementary Figs. S1 and S2).

Cryosections of skin biopsies from abdominal reduction surgery sampled
HLA-DR+ is increased after IFNβ injection in MS patients
Immunohistochemical staining was performed on tissue biopsies from 3 MS donors on IFNβ-1a (s.c.) treatment (Table 1). At the baseline visit, 2 patients received their first IFNβ injection and 1 patient had been on IFNβ treatment for 1 month. No cutaneous reactions were noted after the IFNβ injection at the first visit in any of the subjects. After 11 to 12 months on full-dose IFNβ (44 μg), a skin examination revealed a mild reaction at the IFNβ injection site for 1 MS patient (Table 1). Two out of the 3 patients developed NAb to IFNβ within the first year of treatment. For all 3 MS patients, microscopic examination showed that there was an increased HLA-DR+ staining in the biopsies sampled from the IFNβ injection site compared to the uninjected site at both the baseline visit and visit 2 (Fig. 5a–c). Figure 5d depicts the NAb-negative patient and Figure 5e depicts one of the NAb-positive patients with a high NAb titer (736 ten reduction units [TRU]/mL). At visit 1, the donors' IFNβ-injected skin biopsies showed elevated expression of inflammatory markers HMGB1, IL-1β, IL-6, COX-2, and PGE2 as seen by an increased percentage of stained area compared to the uninjected control sites (Fig. 5d, e). At visit 2, the NAb-negative patient had similar staining pattern as at the baseline visit (Fig. 5d), whereas the NAb-positive patient had decreased staining intensity in the IFNβ-injected biopsy compared to the baseline visit (Fig. 5e). The other NAb-positive patient with a low NAb titer (52 TRU/mL) had similar staining pattern as the NAb-negative patient at visit 1. However, at visit 2, the low NAb-positive patient had increased staining intensity, which more resembled the staining pattern for the high NAb-positive patient at visit 1 (Supplementary Fig. S3). MxA was used as a positive control for IFNβ and was increased after the IFNβ injection compared to the uninjected skin site for the 3 patients at the first visit (Fig. 5f–h). At the second visit, MxA was upregulated in 2 of the patients at the IFNβ injection site (1 NAb negative and 1 NAb positive [52 TRU/mL]) (Fig. 5f, h), whereas no difference was seen in staining intensity between the IFNβ injection site and the uninjected site for one of the patients (NAb positive [736 TRU/mL]) (Fig. 5g).

Punch biopsies were sampled from three multiple sclerosis patients after subcutaneous injection of IFNβ-1a and at an unaffected skin site at 2 time points (visit 1: 4 to 5 h; visit 2: 4 h 40 min to 18 h). Specific staining is shown in brown (DAB) and counterstained with hematoxylin (purple). Immunohistochemical staining was performed with
Patient Treatment Data
IFNβ, interferon beta; NAb, neutralizing antidrug antibodies; TRU/mL, ten reduction units per milliliter.
Discussion
Our study investigated the impact of IFNβ injection on skin-resident cells and 3 main findings were observed. First, inflammatory cytokine release at the site of the intradermal and subcutaneous IFNβ injections suggests that the drug initiates a local innate immune activation. Second, IFNβ enhanced cell maturation markers on skin-resident cells in vivo and ex vivo. Third, no immunological response differences were seen ex vivo between the 3 IFNβ preparations when normalized in dose and administration route.
Immunogenicity is a key limitation for the clinical use of interferons since NAb abrogate the biological efficacy of the drug (Schellekens 2002; Kappos and others 2005; Sorensen and others 2006). The mechanisms that contribute to the break of tolerance are not fully understood, but i.m. administration of IFNβ gives less NAb formation than if the drug is administrated s.c. (Ross and others 2000; Perini and others 2001; Minagara and others 2008), and immunogenicity increases with dosing frequency (Ross and others 2000), indicating that the repetitive breaching of skin might increase the immunogenicity of the drug (Farrell and Giovannoni 2007). Our results show that the local immune response in the skin is affected not only by IFNβ but also when only PBS is injected. Thus, we have concluded that the mere violation of the integrity of the skin evokes an immune reaction.
Studies have shown that a high percentage of the s.c. or i.m. delivered IFNβ never leaves the injection site (Chiang and others 1993; Buchwalder and others 2000). It is therefore not surprising that local inflammatory injection site reactions, in different degrees, are very common adverse events during IFNβ treatment. In a review, Balak and others (2012) describe that injection site reactions occurred in up to 96% of the MS patients using the subcutaneous application route and in up to 33% of MS patients using i.m. injected IFNβ. The mechanisms triggering injection site reactions are thought to be caused by an inflammatory response to IFNβ. Indeed, a study showed that MS skin biopsies sampled 24 h after subcutaneous IFNβ injection had a strong CCL2 and CXCL10 chemokine expression that initiated T cell trafficking from the circulation into the skin tissue (Buttmann and others 2005). These factors might be associated with the inflammatory skin reactions experienced by some of the treated MS patients. In our study, the immunohistochemistry panel is based on mediators that are released upon inflammatory responses that can occur when the tissue is damaged. We observed that the IFNβ injection induced the release of innate cytokines such as HMGB1, IL-1β, and IL-6, which play a key role in the initial phase of inflammation (Eckert 1989). HMGB1 is an alarmin that is expressed in all nucleated cells and is released from necrotic cells upon danger signals, following tissue damage to activate an immune response (Lotze and Tracey 2005). When released, HMGB1 signals through various receptors, including TLR4 (Park and others 2004), which induce secretion of proinflammatory cytokines such as IL-1β, IL-6, or TNF-α (Park and others 2003), which further promote COX-2 production (Zarghi and Arfaei 2011). Thus, we believe that injection of IFNβ triggers translocation of HMGB1 from the nucleus to the cytoplasm and extracellular space, and thus stimulates a further release of secondary proinflammatory mediators, contributing to local inflammation in the tissue. In our ex vivo model, we found less cell migration from the biopsies injected with the high-dose IFNβ, which partly could be explained by potential cell death in the tissue. Another potential mechanism for lack of cell migration was showed by Yen and others (2010), which demonstrated that in vivo treatment with IFNβ inhibited the migration capacity of DCs.
We observed that IFNβ enhanced the maturation markers on skin-resident cells in vivo and ex vivo. Mature DCs have enhanced expression to enable efficient activation and differentiation of naive T cells (Guermonprez and others 2002). Unexpectedly, increased naive CD4+ T cell proliferation was not observed after coculture with IFNβ-stimulated DCs. These data seem to conflict with our results, showing an enhanced expression of costimulatory molecules HLA-DR+ and CD86+, which indicates a mature phenotype. However, several studies have shown that IFNβ alters the function of antigen-presenting cells by downregulating their antigen presentation capacity and thus decreasing the ability to stimulate T cell responses (Jiang and others 1995). Another study has shown that human T cell proliferation is markedly inhibited if the cells are preexposed to IFN-alpha (IFN-α) (Zella and others 2000). Even though such antiproliferative activity appears inconsistent with the ability of IFN-α/β to stimulate T cell expansion, the effects on cell proliferation appear to depend on the timing. For instance, treatment of T cells with IFN-α/β for 18 h or more has an antiproliferative effect, whereas a 3-h preexposure was shown to enhance T cell proliferation (Feng and others 2005). We addressed this by running the supernatant from 4 ex vivo donor biopsy conditions (IFNβ and PBS) using the iLite® bioassay (Euro Diagnostica). The results showed that the IFNβ-injected biopsy supernatant had an IFN concentration (IU/mL) that was equivalent to 32–80 IU/mL of the IFNβ standard curve. Interestingly, also, the PBS-injected skin biopsy supernatant had some IFN present equivalent to 0.33–0.82 IU/mL of the IFNβ standard curve (data not shown). Our data suggest that the DCs are triggered to produce their own IFN since also the PBS condition has traces of IFN. Since small amounts of the cell media from the ex vivo experiment will be transferred to the T cell assay, there will indeed be IFN present during the 5-day incubation. These data collectively suggest that IFN were present in the T cell stimulatory assay and thus could affect the proliferation capacity of the naive CD4+ T cells in this experiment. It should, however, be considered that the IFNβ concentration reaching the cells ex vivo might not reflect the in vivo physiological condition. Moreover, certain stimuli can promote DC activation and maturation and yet induce tolerogenic T cells. This was demonstrated by Waithman and others (2007), where skin-resident DCs expressing a mature phenotype, as measured by the expression level of MHC class II and CD86, still resulted in T cell tolerance. It is also possible that there is a promoted polarization toward follicular helper T cells. Therefore, the lack of T cell proliferation does not confirm the downstream capacity of these T cells to stimulate B cells to produce NAb, and this has to be studied further. Interestingly, a study by Yen and Ganea (2009) showed in a murine model that IFNβ-treated DCs increased the expression of CD86 and at the same time had a significant reduction on T cell proliferation. They discuss that this contradictory effect might be due to the fact that IFNβ also upregulates program death ligand 1 and program death ligand 2 on the DCs, which could cancel the effect of CD86.
This study is coupled to a few limitations. Primarily the study includes individual variations of how the skin cells (ex vivo) reacted to the IFNβ preparations. For example, for some individuals, IFNβ treatment of the CD1a+ dermal DCs induced their migration capacity, whereas it was the opposite effect that was observed for other individuals. Increasing the study population might be beneficial to achieve greater consistency, or showing 2 distinct groups. However, Cole and others (2001) showed that complementary DNA microarray analysis of normal human skin only identified 1.7%, out of the ≈4400 genes tested, as variable in their expression. Thus, even though variability is expected between skin donors, the majority of genes are equally expressed. Another limitation using the ex vivo model is that the therapeutic dose of IFNβ could not be used because the saturation of IFNβ is around 104 U/mL (Buttmann and others 2005), and a control, with the same fluid composition without the active form of IFNβ, could be beneficial. Another aspect to consider in vivo is that before the subcutaneous IFNβ injection, the skin site was prepared with a local anesthetic that has a vasodilating effect. If patients' control site biopsy would have been injected with PBS after a local anesthetic, the control site would have more closely mimicked the event of the IFNβ injection.
Using human skin as a model system for drug screening has the great advantage to maintain the cells in their natural environment and at the same time observe how the cells and extracellular space are affected. Screening of several drugs at an early stage during preclinical development could allow for the least immunogenic to be selected with the aim to improve therapeutic efficacy. A key question yet to be answered is whether the skin DCs take up IFNβ and home the lymph node and initiate an immune response toward the drug. Interestingly, we noted that the skin biopsies sampled after IFNβ injection from one of the MS patient that became NAb positive had reduced immunostaining at the second visit (12 months) compared to the first visit (1 month on treatment). We speculate that it might be due to the fact that the NAb bind to the drug in close proximity to the skin site and limit the drugs' immunomodulatory properties, or that the patient had become more tolerant to the drug throughout the treatment year. If we find that the immunology of the skin has a considerable impact on NAb development, there are preventative regimes that could be implemented. The skin is readily available for different stimulations; for example, the use of a topical anti-inflammatory cream such as hydrocortisone before IFNβ administration (Pozzilli and others 2002) or UV light (Loser and Beissert 2009) could potentially be used to minimize the risk of local immune activation and hence NAb formation.
Footnotes
Acknowledgments
The authors wish to thank Bibbi Svensson for providing us with skin explants. We thank all patients for their participation in this study. We also thank Ingegerd Löfving-Arvholm for excellent technical assistance in cell culture work. The research leading to these results has received support from the Innovative Medicines Initiative Joint Undertaking under grant agreement no. 115303, resources of which are composed of financial contribution from the European Union's Seventh Framework Programme (FP7/2007–2013) and EFPIA companies' in kind contribution.
Author Disclosure Statement
C.H. has no conflict of interests to report for this study. T.M.M.v.C. has no conflict of interests to report for this study. M.A. has no conflict of interests to report for this study. V.K. has no conflict of interests to report for this study. F.D. has participated in meetings sponsored by or received honoraria for acting as an advisor/speaker for Biogen Idec, Genzyme-Sanofi, Merck, Novartis Pharma, Roche, Shire, and TEVA ratiopharm. He received research funding from Biogen and Genzyme-Sanofi. He is section editor of the MSARD Journal (Multiple Sclerosis and Related Disorders). E.C.d.J. has no conflict of interests to report for this study. A.F.H. has no conflict of interests to report for this study.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3