
Abstract
Mevalonate Kinase Deficiency (MKD) is an autoinflammatory disease caused by mutations in the mevalonate kinase gene, which produces an enzyme responsible for the production of isoprenoids in the mevalonate pathway. Patient data indicate that MKD is a multicytokine disease with increased plasma levels of cytokines such as tumor necrosis factor-α, interleukin (IL)-6, and interferon (IFN)-γ. To study the mechanisms responsible for these changes, the mevalonate pathway was inhibited with lovastatin in peripheral blood mononuclear cells (PBMCs) and monocytes isolated from the blood of healthy donors followed by stimulation with lipopolysaccharide (LPS) to induce an inflammatory response. Lovastatin treatment resulted in increased levels of IL-6, IL-12p40, and IFN-γ mRNA in both PBMCs and monocytes following LPS stimulation compared with control cells. An IL-12 neutralizing antibody blocked the increased levels of IFN-γ mRNA following lovastatin treatment in PBMCs indicating that this effect is dependent on IL-12. Flow cytometry experiments indicated that monocytes, not lymphocytes or granulocytes, are the source of increased IFN-γ and that both classical and nonclassical/intermediate monocytes express IFN-γ. These results indicate that blocking IL-12 or IFN- γ may be therapeutic options for MKD patients.
Introduction
Mevalonate kinase deficiency (MKD) is a rare autoinflammatory disorder characterized by recurrent episodes of symptoms such as fever, headache, joint pain, skin rashes, vomiting, and diarrhea. It can be subdivided into the less severe form of hyperimmunoglobulinemia D and periodic fever syndrome and the more severe form of mevalonic aciduria (MA). In addition to the symptoms listed above, MA patients also suffer from neurological symptoms and developmental abnormalities (Favier and Schulert 2016). During episodes, patients have increased circulating and ex vivo production of cytokines and acute phase protein levels, including C-reactive protein, phospholipase A2, interleukin (IL)-1β, tumor necrosis factor (TNF)-α, IL-6, and interferon (IFN)-γ (Drenth and others 1995a, 1995b, 1996).
MKD is caused by mutations in the mevalonate kinase (MVK) gene, which produces an enzyme in the mevalonate pathway responsible for the production of sterol isoprenoid compounds such as cholesterol and nonsterol isoprenoid compounds such as farnesyl- and geranylgeranylpyrophosphate (FPP and GGPP) (Fig. 1). Alterations to cytokine levels and inflammatory symptoms of the disease have been attributed to depletion of GGPP in monocytes (Frenkel and others 2002; Mandey and others 2006), and GGPP or geraniol (GOH, a more soluble form of GGPP) is able to rescue inflammatory phenotypes in the MKD mouse models (Marcuzzi and others 2008, 2011), RAW 264.7 macrophages (Frey and De Maio 2007; Marcuzzi and others 2011), and human peripheral blood mononuclear cells (PBMCs) (Mandey and others 2006). GGPP is responsible for the prenylation and subsequent membrane localization of the Rho and Rab families of small GTP-binding proteins (Wang and Casey 2016).
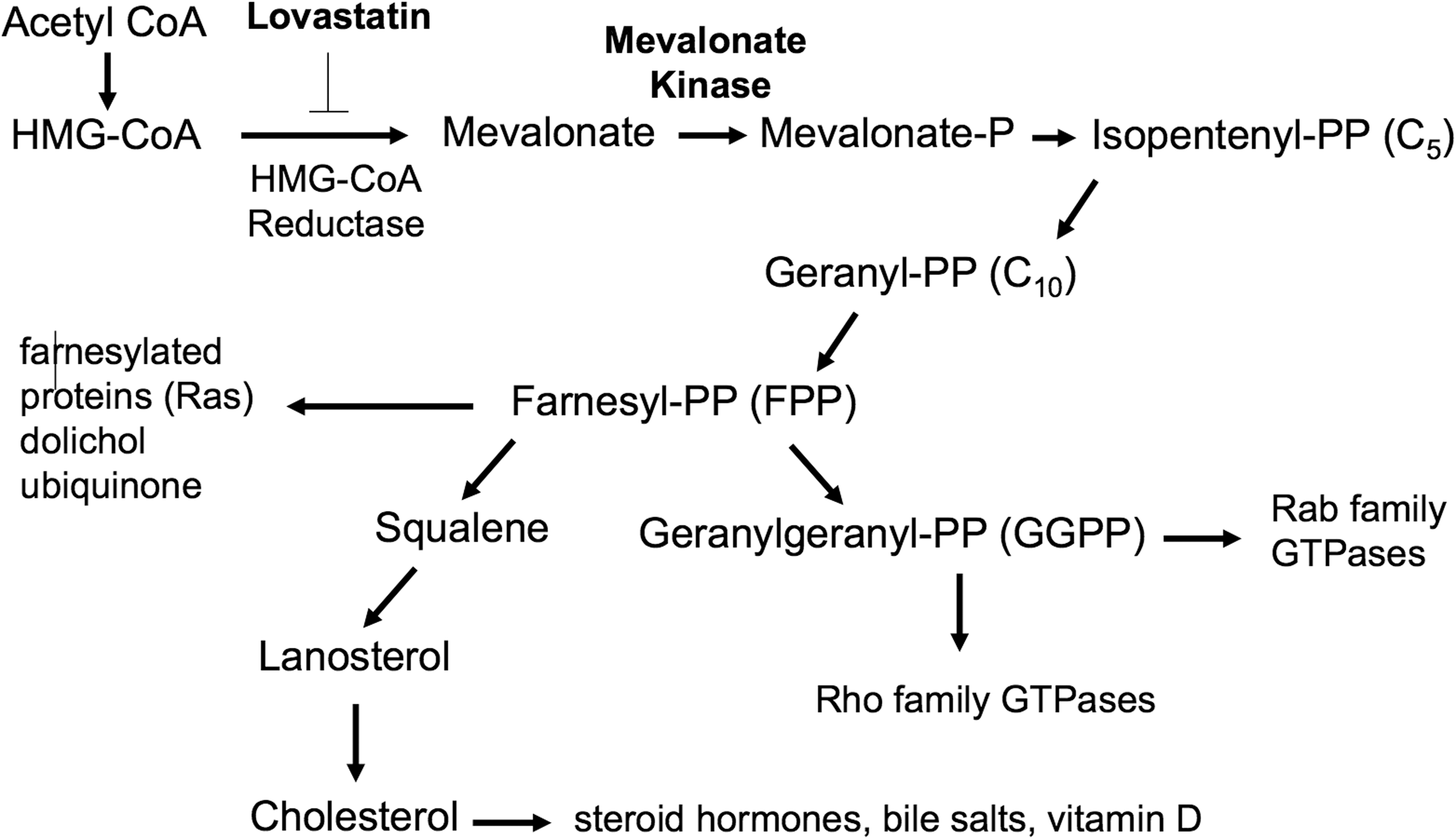
The mevalonate pathway. The mevalonate pathway produces both nonsterol isoprenoids such as FPP and GGPP as well as sterol isoprenoids such as cholesterol. MKD patients have mutations in mevalonate kinase. Lovastatin blocks HMG-CoA reductase, which is one step upstream of mevalonate kinase and is widely used to model MKD in culture. An abbreviated version of the pathway is shown illustrating only relevant molecules. MKD, mevalonate kinase deficiency; FPP and GGPP, farnesyl- and geranylgeranylpyrophosphate.
Most of the mechanistic research surrounding MKD involves the increased secretion of IL-1β by monocytes. The amount of IL-1β released by cells depends on expression levels, processing, and secretion. Inflammatory stimuli such as lipopolysaccharide (LPS) induce increased expression of pro-IL-1β through the activation of inflammatory signaling pathways (Oviedo-Boyso and others 2014). Pro-IL-1β is inactive and requires cleavage by caspase 1, which is activated by inflammasome assembly (Broz and Dixit 2016). Blocking the mevalonate pathway in the THP-1 human monocyte-like cell line has been reported to synergize with LPS to induce the release of IL-1β (Kuijk and others 2008b). This effect was due to activation of caspase 1 and increased processing of pro-IL-1β. It was subsequently reported in the same model that isoprenoid depletion by treatment with simvastatin caused activation of the Rho GTPase Rac1 leading to the phosphoinositide 3-kinase (PI3K)-dependent phosphorylation of protein kinase B and caspase-1 activation (Kuijk and others 2008a). Later, roles for unprenylated RhoA in inflammasome activation were discovered. Lack of RhoA prenylation resulted in decreased activation of RhoA (van der Burgh and others 2014; Park and others 2016), which activated Rac1 (van der Burgh and others 2014) and also resulted in direct activation of the pyrin inflammasome through reduced pyrin phosphorylation leading to increased IL-1β release (Park and others 2016).
The central role of IL-1β in MKD pathogenesis is highlighted by the effectiveness of IL-1 blockade in treating patients (De Benedetti and others 2018). However, IL-1β blockade is not always successful in treating MKD, and it has been reported to be a multicytokine disease with elevated secreted levels of the non-IL-1 family cytokines TNF-α, IL-6, and IFN-γ in plasma during attacks (Drenth and others 1995a, 1995b) and TNF-α and IL-6 following ex vivo stimulation of patient PBMCs with the toll-like receptor (TLR)2/TLR4 agonists (Stoffels and others 2015). In addition, blocking geranylgeranyl transferase I (and therefore the prenylation of cellular targets by GGPP) in mouse bone marrow-derived macrophages (BMDMs) resulted in increased protein and mRNA levels of IL-1β, TNF-α, IL-6, and IL-12p40 levels following LPS stimulation (Akula and others 2016). Despite these interesting findings, a thorough investigation of cytokine mRNA levels in human cells, particularly IFN-γ, is lacking. Therefore, we examined a panel of cytokine mRNA levels in both primary PBMCs and monocytes from healthy donors following isoprenoid depletion with a statin and stimulation of inflammation with LPS. Isoprenoid depletion in PBMCs and monocytes caused increased mRNA levels of TNF-α, IL-6, IL-12p40, and IFN-γ following LPS stimulation. In addition, monocytes were responsible for the increased IFN-γ levels following isoprenoid depletion and increased IFN- γ mRNA levels were blocked by an IL-12 neutralizing antibody indicating that increased IL-12 is necessary for this effect.
Materials and Methods
Cells
THP-1 cells were obtained from ATCC (Manassas, VA) and cultured in Roswell Park Memorial Institute containing 10% fetal bovine serum in a 37°C, 5% CO2 incubator. PBMCs were isolated from the blood of healthy donors obtained from the Holy Spirit Blood Bank (Camp Hill, PA) or the Penn State College of Medicine Blood Donor Center (Hershey, PA). Filter packs were flushed with 150 mL sterile, room temperature phosphate buffered saline (PBS) and 20 mL was layered onto 15 mL Ficoll-Paque PLUS (GE Health care, Chicago, IL). The tubes were centrifuged at 630g for 30 min, and the buffy coat layer was transferred to new tubes, and PBS was added to 160 mL to wash the lymphocytes. The tubes were centrifuged at 375g for 10 min and the cell pellets were resuspended in a total volume of 10 mL AIM-V media (Life Technologies, Carlsbad, CA) and counted. Primary monocytes were isolated from PBMCs using the Pan Monocyte Isolation Kit (Miltenyi Biotec, Bergisch Gladbach, Germany). Cells were cultured with various treatments at a concentration of 0.5 × 106 cells/mL (THP-1 cells) or 1 × 106 cells/mL (PBMCs and primary monocytes).
Treatments
Cells were treated with 10 μM lovastatin (Millipore, Burlington, MA) for 16–18 h followed by stimulation with 100 ng/mL Escherichia coli LPS (026:B6; Sigma-Aldrich, St. Louis, MO) for varying amounts of time. At times, 100 μM mevalonate was added at the same time as lovastatin. For the IL-12 neutralizing experiments, 1,000 ng/mL IL-12 or isotype control antibodies (R&D Systems, Minneapolis, MN) were added after lovastatin treatment at the same time as LPS stimulation.
Real-time polymerase chain reaction
Following treatment, cells were homogenized in TRIzol reagent (Life Technologies, Carlsbad, CA), and RNA was isolated using the Direct-zol RNA Miniprep (Zymo Research, Irvine, CA). RNA was converted to cDNA using the High Capacity cDNA Reverse Transcription Kit (Life Technologies, Carlsbad, CA) and ∼10 ng of converted RNA was used in a real-time polymerase chain reaction (RT-PCR) with iTaq Universal SYBR Green Supermix (Bio-Rad, Hercules, CA) and 250 nM forward and reverse primers (see Table 1 for sequences) on the CFX384 real-time PCR machine (Bio-Rad, Hercules, CA). Samples were quantified using the delta delta Ct relative quantitation method with GAPDH as the endogenous control gene and set relative to carrier-treated cells.
Real-Time Polymerase Chain Reaction Human Primer Sequences
TNF, tumor necrosis factor; IL, interleukin; IFN, interferon.
Flow cytometry
All flow cytometry protocols and reagents were obtained from Miltenyi Biotec (Bergisch Gladbach, Germany). In brief, if intracellular IFN-γ was examined, cells were treated with 3 μg/mL brefeldin A for 4 h before harvest. Upon harvest, cells were collected by centrifugation at 300g and washed with Miltenyi FACs buffer (phosphate-buffered saline, pH 7.2, 0.5% bovine serum albumin, and 2 mM ethylene-diaminetetra-acetic acid). Cell surface antigens (CD14, CD16, and CD45) were then stained according to Miltenyi protocols followed by fixation and permeabilization using the Inside Stain Kit. Cells were then stained for IFN-γ using the Miltenyi protocol. Stained cells were stored at 4°C for up to 24 h before analysis using the 16-color LSR Fortessa Flow Cytometer in the Penn State College of Medicine Flow Cytometry Core Facility. For compensation controls, UltraComp eBeads from Thermo Fisher Scientific (Waltham, MA) were stained with antibodies according the manufacturer's protocol. Data were analyzed using FlowJo software.
Enzyme-linked immunosorbent assay
IFN-γ protein levels were analyzed in cell culture supernatants using the Human IFN-gamma Quantikine enzyme-linked immunosorbent assay (ELISA) kit from R&D Systems (Minneapolis, MN).
Results
Lovastatin treatment increases inflammatory gene expression following LPS stimulation in PBMCs
Statins are competitive inhibitors of HMG-CoA reductase and block the production of mevalonate just 1 step upstream of MVK in the mevalonate pathway (Fig. 1) and are therefore widely used to model MKD in cells (Mandey and others 2006; Frey and De Maio 2007; Kuijk and others 2008a, 2008b; van der Burgh and others 2014; Stoffels and others 2015; Akula and others 2016). In addition, inflammatory stimuli such as the TLR4 ligand LPS amplify MKD phenotypes (Stoffels and others 2015). PBMCs isolated from the blood of healthy donors (n = 3–5) were therefore treated with lovastatin to deplete isoprenoids followed by stimulation with LPS for varying amounts of time. Inflammatory gene expression was measured by RT-PCR. Lovastatin treatment had no effect on mRNA levels in the absence of LPS stimulation (time point 0) for any of the genes examined (Fig. 2). Following LPS stimulation, the mRNA levels of all genes increased at all time points. In addition, lovastatin treatment resulted in significantly higher levels of TNF-α (2.15-fold at 4 h, P = 0.004 and 2.29-fold at 8 h, P = 0.02), IFN-γ (203.4-fold at 24 h, P = 0.005), and IL-6 (3.46-fold at 8 h, P = 0.044) compared with carrier-treated samples (Fig. 2A–C). Lovastatin treatment had no effect on the mRNA levels of IL-1β, IL-10, or CD14 compared with carrier-treated samples at any of the time points examined (Fig. 2D–F).
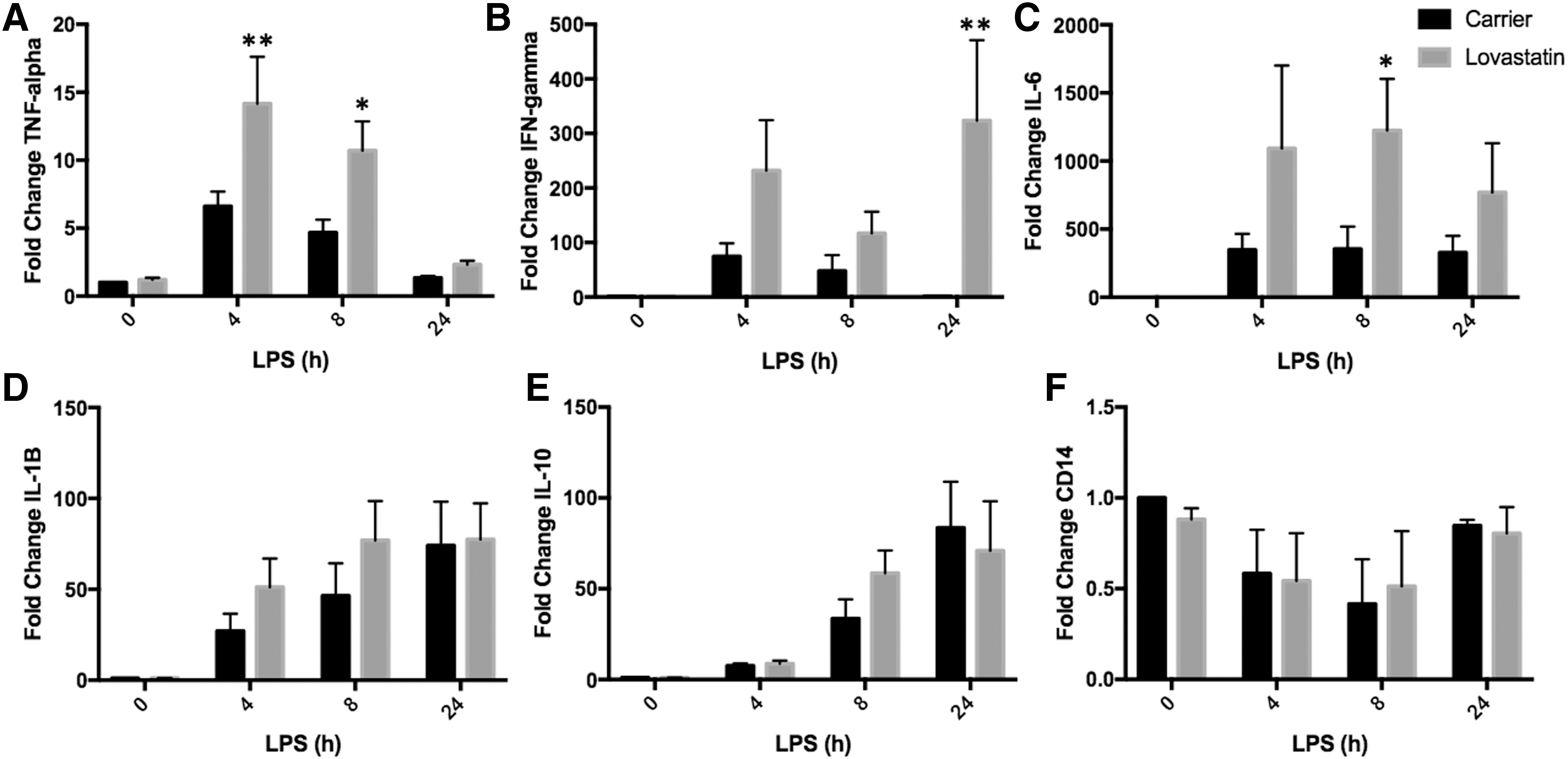
Lovastatin treatment increases inflammatory gene expression following LPS stimulation in PBMCs. PBMCs (n = 3–5) were isolated from the blood of healthy donors and treated with lovastatin for 16–18 h followed by LPS stimulation for varying amounts of time. mRNA levels were measured by RT-PCR. TNF-α mRNA levels were significantly higher in lovastatin compared with carrier-treated cells 4 h (2.15-fold, **P = 0.004) and 8 h (2.29-fold, *P = 0.02) following LPS stimulation
Isoprenoid depletion increases cytokine mRNA levels following LPS stimulation
To determine if the changes in inflammatory gene expression following LPS stimulation were due to isoprenoid depletion and not an off target effect of lovastatin, lovastatin-treated PBMCs (n = 8–10) were coincubated with mevalonate. Mevalonate is the immediate downstream product of the enzyme inhibited by lovastatin (Fig. 1), and therefore, addition of mevalonate to lovastatin-treated samples will prevent any changes in gene expression induced by lovastatin that are caused by depletion of the mevalonate pathway. Cytokine mRNA levels after the addition of mevalonate were only measured at the LPS stimulation time points that showed significant differences between carrier and lovastatin-treated cells (8 h for IL-6 and TNF-α or 24 h for IFN-γ). As observed in the first set of donors (Fig. 2), both IL-6 (2.44-fold, P = 0.05) and IFN-γ (5.32-fold, P = 0.04) mRNA levels were increased in lovastatin-treated samples compared with carrier-treated samples following LPS stimulation for 8 and 24 h, respectively (Fig. 3A, B). Moreover, coincubation with mevalonate prevented the increased mRNA levels observed in the presence of lovastatin alone (Fig. 3A, B). TNF-α mRNA levels were increased in the lovastatin compared with carrier-treated cells (1.65-fold), but unlike the first data set, this difference was not statistically significant (Fig. 3C).
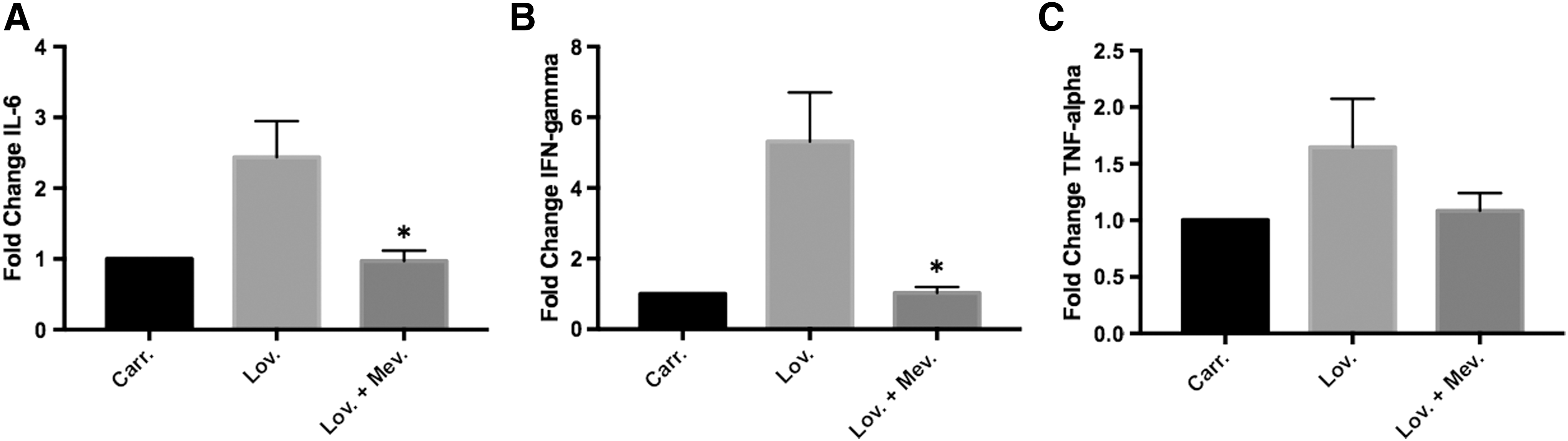
Isoprenoid depletion increases cytokine mRNA levels after LPS stimulation. PBMCs (n = 8–10) were isolated from the blood of healthy donors and treated with lovastatin ± mevalonate followed by LPS stimulation for 8 h (IL-6 and TNF) or 24 h (IFN-γ). mRNA levels were measured by RT-PCR. The addition of mevalonate prevented the increased IL-6 mRNA levels observed in the presence of lovastatin alone
Isoprenoid depletion increases IL-12p40 mRNA levels following LPS stimulation
Both IL-12 and IL-18 are known to cause increases in IFN-γ mRNA levels in target cells (Coward and others 2006). Therefore, both IL-12p40 and IL-18 mRNA levels were measured in lovastatin-treated PBMCs (n = 4) following LPS stimulation. Lovastatin treatment had no effect on IL-18 or IL-12p40 mRNA levels compared with carrier-treated cells in the absence of LPS stimulation (Fig. 4A, B). Both IL-18 and IL-12p40 mRNA levels increased following LPS stimulation at all time points examined (Fig. 4A, B). In addition, IL-12p40 mRNA levels were significantly increased in lovastatin-treated cells compared with carrier-treated cells (12.58-fold, P = 0.037) 8 h following LPS stimulation (Fig. 4B). Lovastatin had no effect on IL-18 mRNA levels following LPS stimulation at any of the time points examined (Fig. 4A). Moreover, coincubation of lovastatin-treated cells with mevalonate (n = 8) prevented increased levels of IL-12p40 8 h following LPS stimulation (Fig. 4C, D), indicating that this effect is due to isoprenoid depletion.
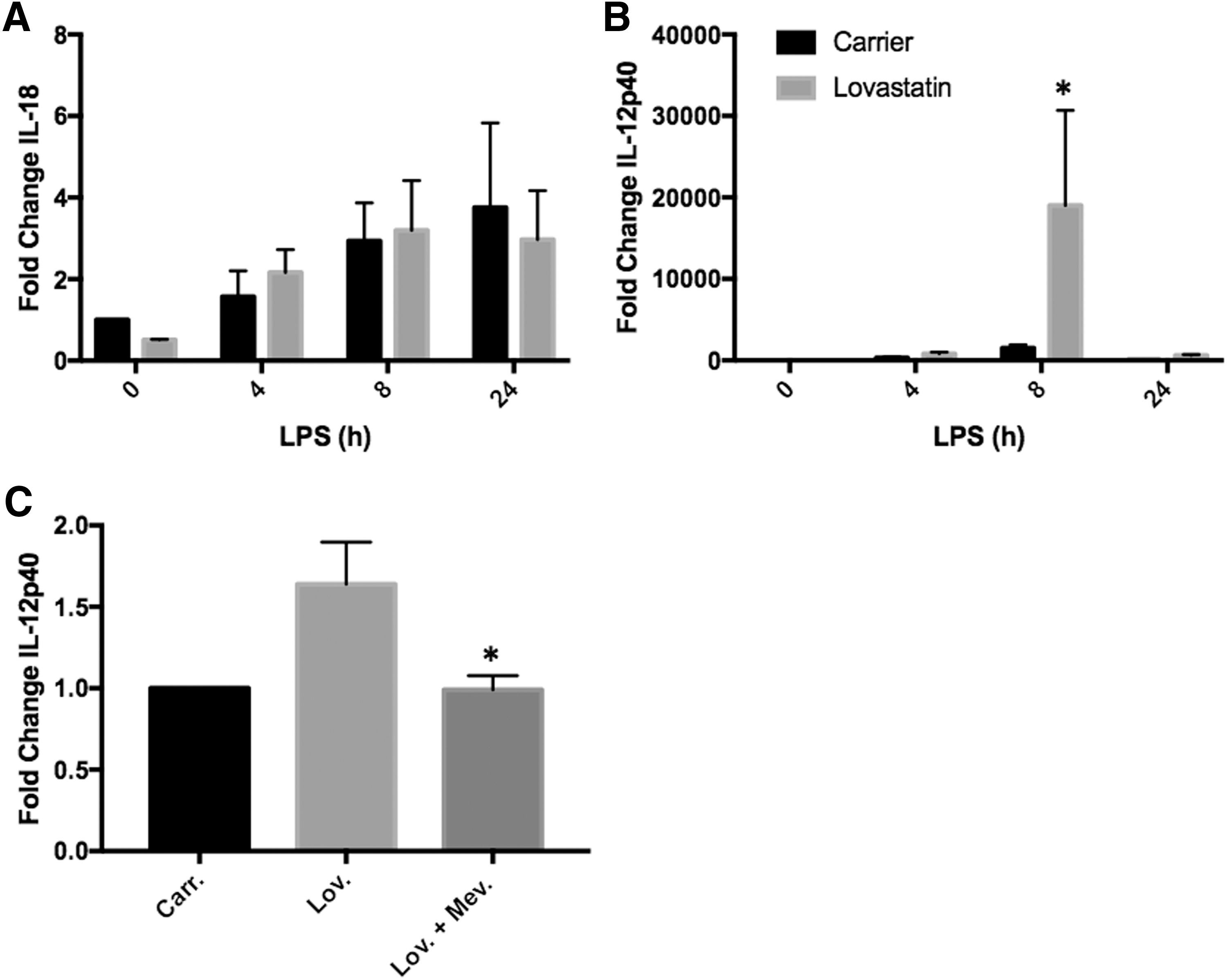
Isoprenoid depletion increases IL-12p40 mRNA levels following LPS stimulation. PBMCs (n = 4) were isolated from the blood of healthy donors and treated with lovastatin for 16–18 h followed by LPS stimulation for varying amounts of time. mRNA was measured by RT-PCR. IL-18 mRNA levels were not significantly different between carrier and lovastatin treated cells at all time points examined
IL-12 causes increased IFN-γ mRNA levels following LPS stimulation
To determine if increased IL-12 levels are responsible for increased IFN-γ mRNA levels in lovastatin-treated PBMCs following LPS stimulation, PBMCs (n = 4) were treated with an IL-12 neutralizing antibody or isotype-matched control antibody. As previously observed, IFN-γ mRNA levels were increased in lovastatin-treated PBMCs compared with carrier-treated PBMCs 24 h following LPS stimulation (6.36-fold, P = 0.015, Fig. 5). Addition of an isotype control antibody at the time of LPS stimulation had no effect on the increased IFN-γ mRNA levels observed in lovastatin-treated cells (4.54-fold, P = 0.045, Fig. 5). However, addition of an IL-12-neutralizing antibody at the time of LPS stimulation resulted in an overall decrease in IFN-γ mRNA levels in all samples and prevented the increase in IFN-γ mRNA levels in lovastatin-treated cells compared with carrier-treated cells (Fig. 5). However, addition of an IL-12-neutralizing antibody had no significant effect on IL-1β, TNF-α, or IL-6 mRNA levels in carrier or lovastatin-treated PBMCs compared to isotype control or untreated cells (n = 4, data not shown).
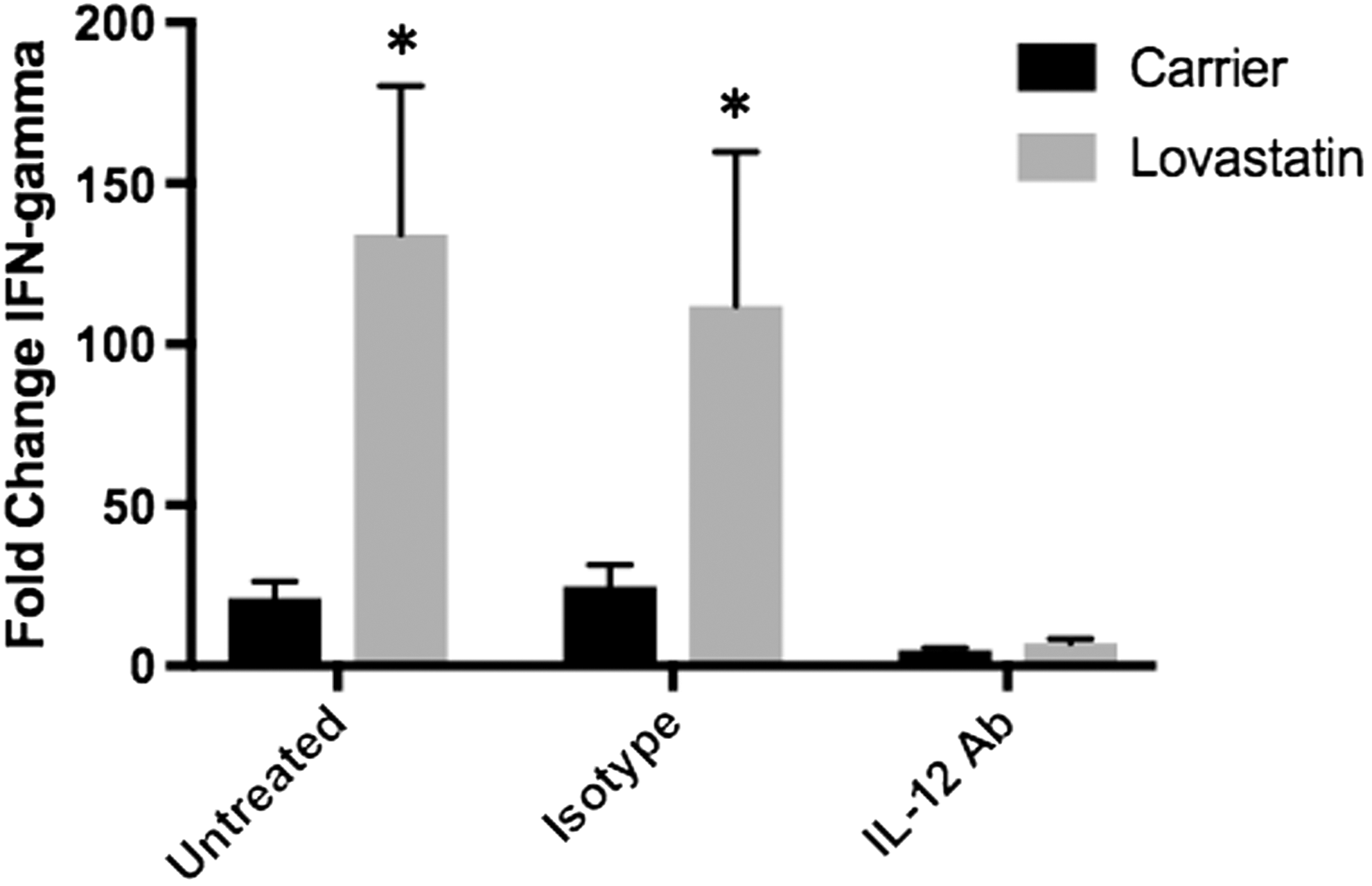
IL-12 causes increased IFN-γ mRNA levels following LPS stimulation. PBMCs were isolated from the blood of healthy donors and treated with lovastatin for 16–18 h followed by LPS stimulation for 24 h. At the time of LPS addition, an IL-12 neutralizing or isotype-matched control antibody was added to the cells. IFN-γ mRNA levels were significantly increased in lovastatin compared with carrier-treated cells (*P = 0.015) and this increase was also observed after the addition of an isotype-matched control antibody (*P = 0.045). No difference in IFN-γ mRNA levels were observed between lovastatin and carrier-treated samples after the addition of an IL-12 neutralizing antibody. Statistical analysis was done by 2-way ANOVA.
Monocytes produce IFN-γ
Since PBMCs are a mixture of mononuclear cells (monocytes, lymphocytes, and some granulocytes), the source of increased cytokine levels following lovastatin treatment was examined. Monocytes were purified from PBMCs by negative selection using the Pan Monocyte Isolation Kit (Miltenyi). Following purification, monocytes from 1 donor were subjected to flow cytometry to assess the purity of the population. CD45 is a cell surface marker that is expressed on all leukocytes. Along with side scatter (SSC-A), CD45 staining (PE label) can be used to separate monocytes from lymphocytes and granulocytes. Before purification, the PBMC mixture contained 3 identifiable populations of cells with ∼64.1% lymphocytes, 16.5% granulocytes, and 7.1% monocytes (Fig. 6A). Of the monocytes, CD14 staining (FITC label) indicated that 89.2% were CD14 positive (Fig. 6A). Following purification, only one clear population of cells was identifiable using CD45 versus SSC-A, which fell in the monocyte region and contained 87.9% of the cells (Fig. 6B). Of those cells, 88.8% were CD14 positive (Fig. 6B). The purified monocytes (n = 4) were treated with lovastatin for 16–18 h followed by LPS stimulation and inflammatory gene expression was measured by RT-PCR. Similar to PBMCs, lovastatin treatment increased IL-6 (2.13-fold, P = 0.064) and IL-12p40 mRNA levels (3.05-fold, P = 0.0045) in monocytes compared with carrier-treated cells 8 h following LPS stimulation (Fig. 6C, D). In addition, IFN-γ mRNA levels were significantly higher in lovastatin compared to carrier-treated monocytes 24 h following LPS stimulation (34.18-fold, P = 0.0028, Fig. 6E).
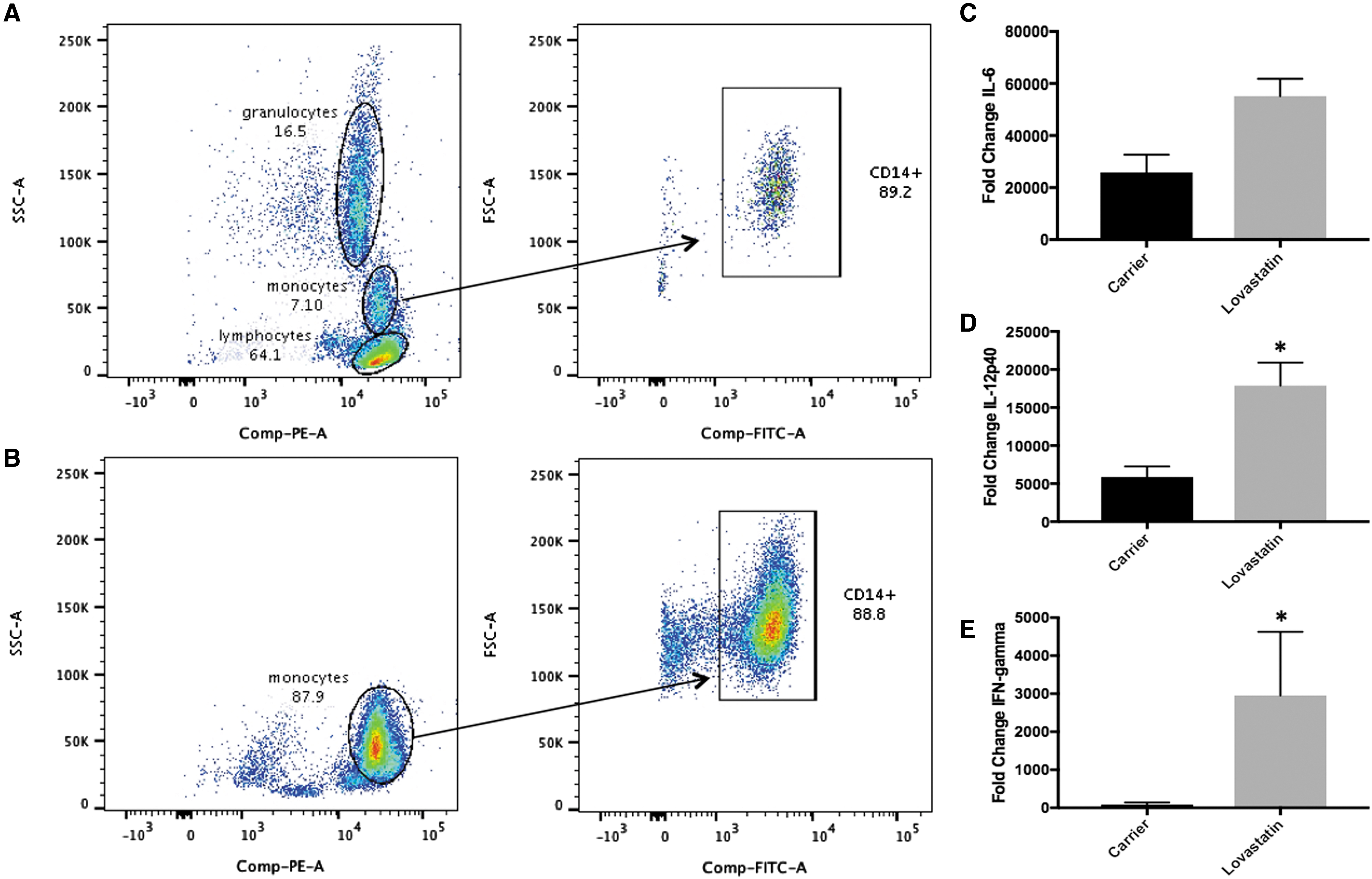
Monocytes express IFN-γ. Monocytes were isolated from PBMCs obtained from the blood of healthy donors (n = 4) and the purity of the population was analyzed by flow cytometry
Since the THP-1 human monocyte cell line is often used as a cell culture model system to study the MKD cytokine response, we examined relevant cytokine mRNA levels following lovastatin treatment and LPS stimulation in THP-1 cells. Of the cytokines that had increased mRNA levels after lovastatin treatment in primary PBMCs and monocytes, only TNF-α and IL-12p40 exhibited a similar increase in THP-1 cells following LPS stimulation with a 3.48-fold (P = 0.04) and 2.44-fold (P = 0.07) increase in mRNA levels compared with carrier-treated cells 6 h after LPS stimulation for TNF-α and IL-12p40, respectively (Fig. 7A, B). Unlike primary monocytes, no difference in IL-6 mRNA levels was observed in lovastatin-treated THP-1 cells compared to carrier following LPS stimulation (Fig. 7C). As previously reported, IL-1β mRNA levels were not significantly increased in lovastatin-treated THP-1 cells compared to carrier following LPS stimulation (Fig. 7D). IFN-γ levels were not detectable in THP-1 cells under any treatment conditions or time points.
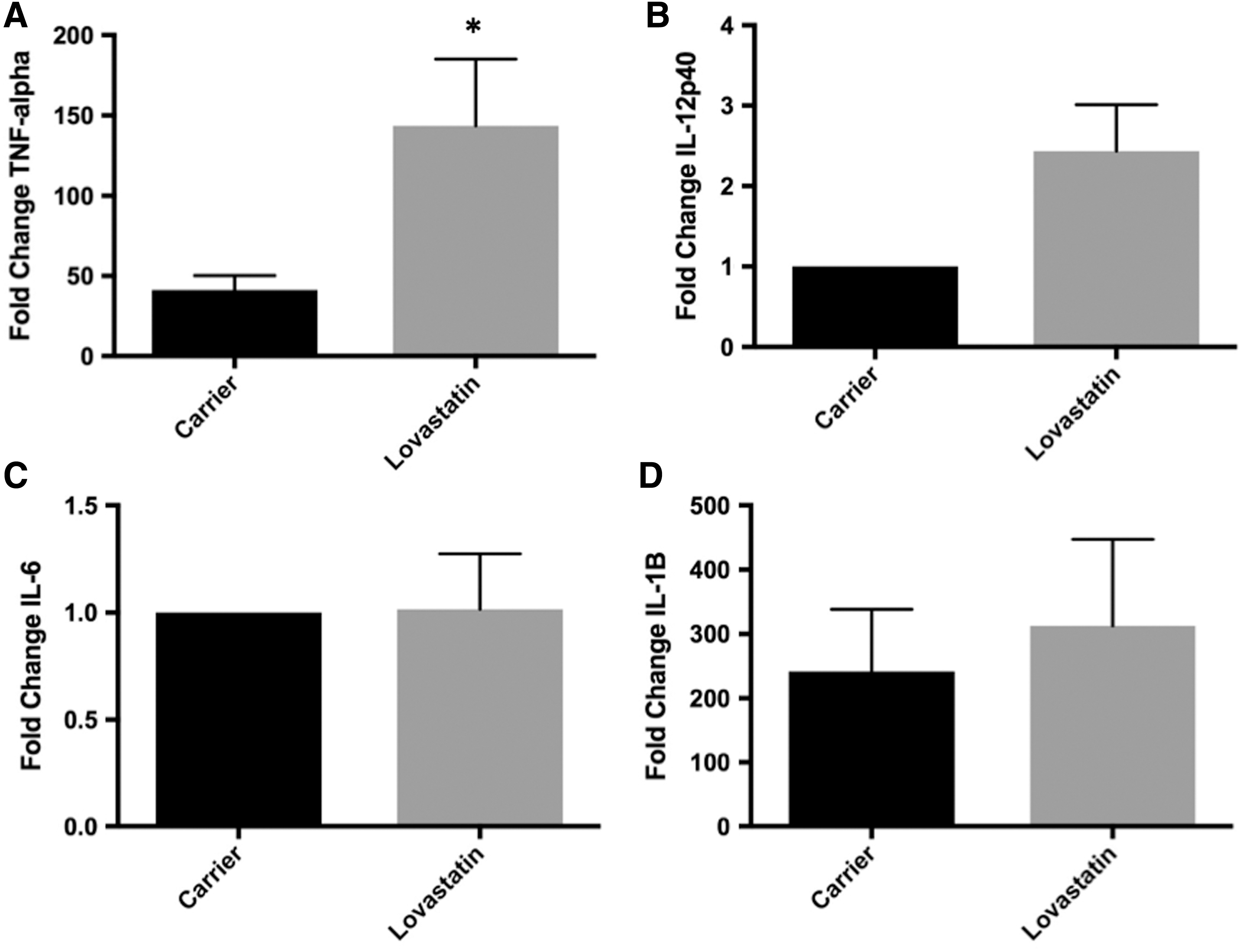
Effect of lovastatin treatment on cytokine mRNA levels in the THP-1 human monocyte cell line. THP-1 monocytes (n = 3–5) were treated with lovastatin for 16–18 h followed by LPS stimulation for 6 h. mRNA levels were measured by RT-PCR. TNF-α mRNA levels were significantly higher in lovastatin compared with carrier-treated cells following LPS stimulation
THP-1 cells are a monocyte-like cell line derived from a patient with acute monocytic leukemia (Tsuchiya and others 1980). However, they are not fully differentiated monocytes and have low expression of CD14, which is a monocyte differentiation marker and LPS receptor, leading to reduced responsiveness to LPS stimulation compared to primary monocytes (Bosshart and Heinzelmann 2016). In contrast, primary human monocytes can be divided into 3 subsets based on the expression of cell surface markers and how they respond to stimulation. Classical monocytes express high levels of CD14 and no CD16 (CD14++CD16−), while nonclassical monocytes express CD16 and lower levels of CD14 (CD14dimCD16++). In addition, an intermediate population has been identified that express high levels of CD14 and low levels of CD16 (CD14++CD16+) (Boyette and others 2017). Therefore, THP-1 cells do not represent the range of primary human monocyte phenotypes. To determine how lovastatin treatment affects primary human monocyte populations and if a particular population is responsible for IFN-γ expression, PBMCs (n = 4) were stained with fluorescently labeled antibodies against CD14 (FITC), CD16 (PE), CD45 (APC-Vio770), and IFN-γ (APC) and subjected to flow cytometry. To determine if lovastatin treatment affects monocyte populations, all CD14+ cells were analyzed for CD16 expression and monocytes were classified as classical (CD14+CD16−) or nonclassical/intermediate (CD14+CD16+). Lovastatin treatment significantly increased the classical population (95.8% versus 90.9%, P = 0.011) with a corresponding reduction in the nonclassical/intermediate population (4.2% versus 9.1%, P = 0.011) compared with carrier-treated cells (Fig. 8).
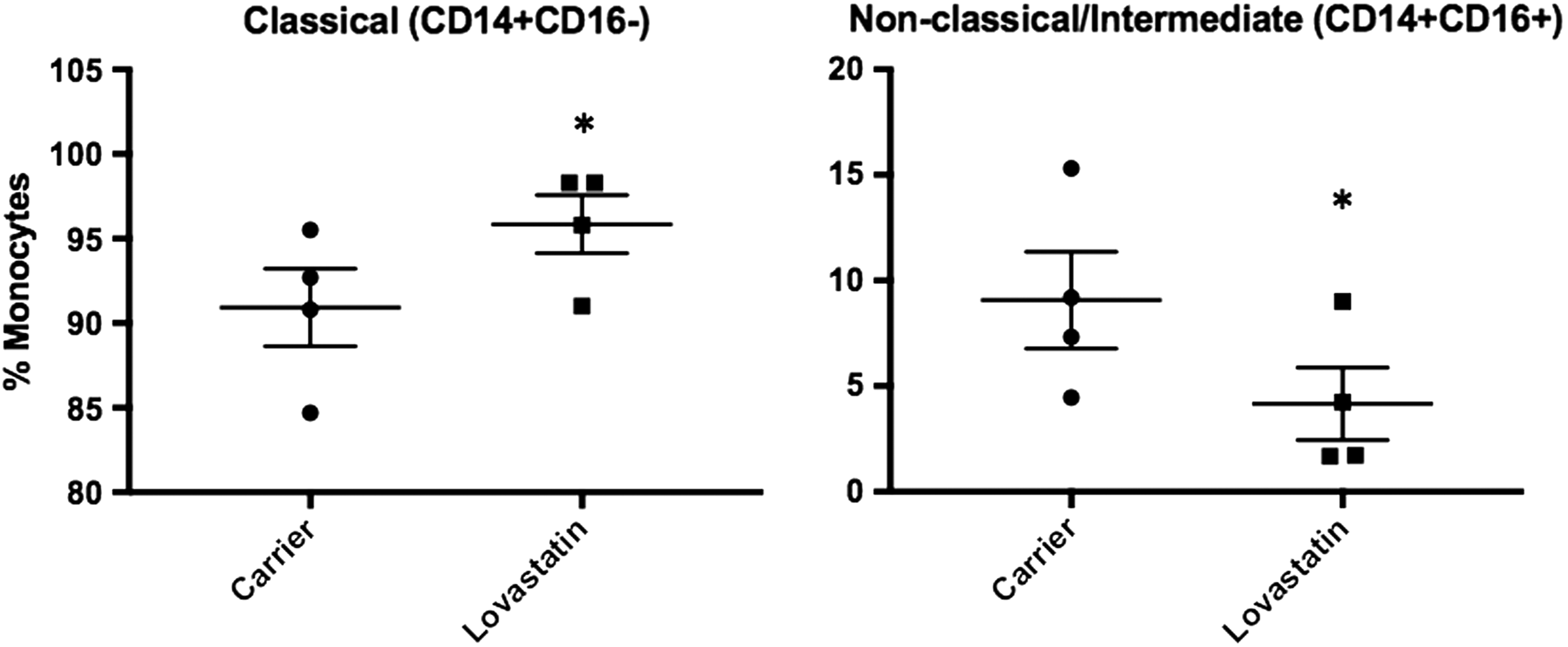
Lovastatin shifts the monocyte population toward classical. PBMCs (n = 4) were treated with lovastatin for 16–18 h followed by staining for CD14 (FITC) and CD16 (PE) and analysis by flow cytometry. CD14-positive cells were examined for the expression of CD16, and monocytes were divided into classical (CD14++CD16−) and nonclassical/intermediate (CD14++CD16+, CD14dimCD16++) subsets. Lovastatin treatment significantly increased the percentage of classical monocytes (*P = 0.011) and decreased the percentage of nonclassical/intermediate monocytes (*P = 0.011) compared with carrier treated cells. Statistical analysis was done by 2-way ANOVA.
Next, to determine which monocyte populations express IFN-γ following LPS stimulation, CD14 expression and CD16 expression were examined within the IFN-γ-positive population. The gating strategy for this experiment is shown for 1 donor in Fig. 9A (carrier) and 9B (lovastatin). First, IFN-γ (APC labeled)-positive cells were selected from the entire PBMC population. The IFN-γ-positive cells were then analyzed by CD45 (APC-Vio770 labeled) versus SSC-A, and lymphocyte, granulocyte, and monocyte populations were selected. Finally, the monocyte population was analyzed for CD14 (FITC label) and CD16 (PE label) expression. Lovastatin treatment significantly increased the percent of IFN-γ-positive monocytes following LPS stimulation (24 h) from 31% in carrier-treated cells to 67% in lovastatin-treated cells (Fig. 9C, n = 3, P = 0.034). In contrast, lovastatin treatment had no effect on the percentage of lymphocytes or granulocytes that were IFN-γ positive (Fig. 9C). Both classical and nonclassical/intermediate monocytes were IFN-γ positive with the percentages of each reflecting the shifts observed in the entire monocyte population (Fig. 8, data for 1 donor shown in Fig. 9A, B). These results indicate that lovastatin treatment causes an increased percentage of primarily classical monocytes to express IFN-γ protein compared with carrier-treated cells. Finally, the secretion of increased IFN-γ protein levels following lovastatin treatment was confirmed by ELISA. Lovastatin treatment resulted in significantly higher levels of IFN-γ protein in the supernatant 24 h following LPS stimulation compared with carrier-treated PBMCs (Fig. 9D, n = 3, P = 0.036).
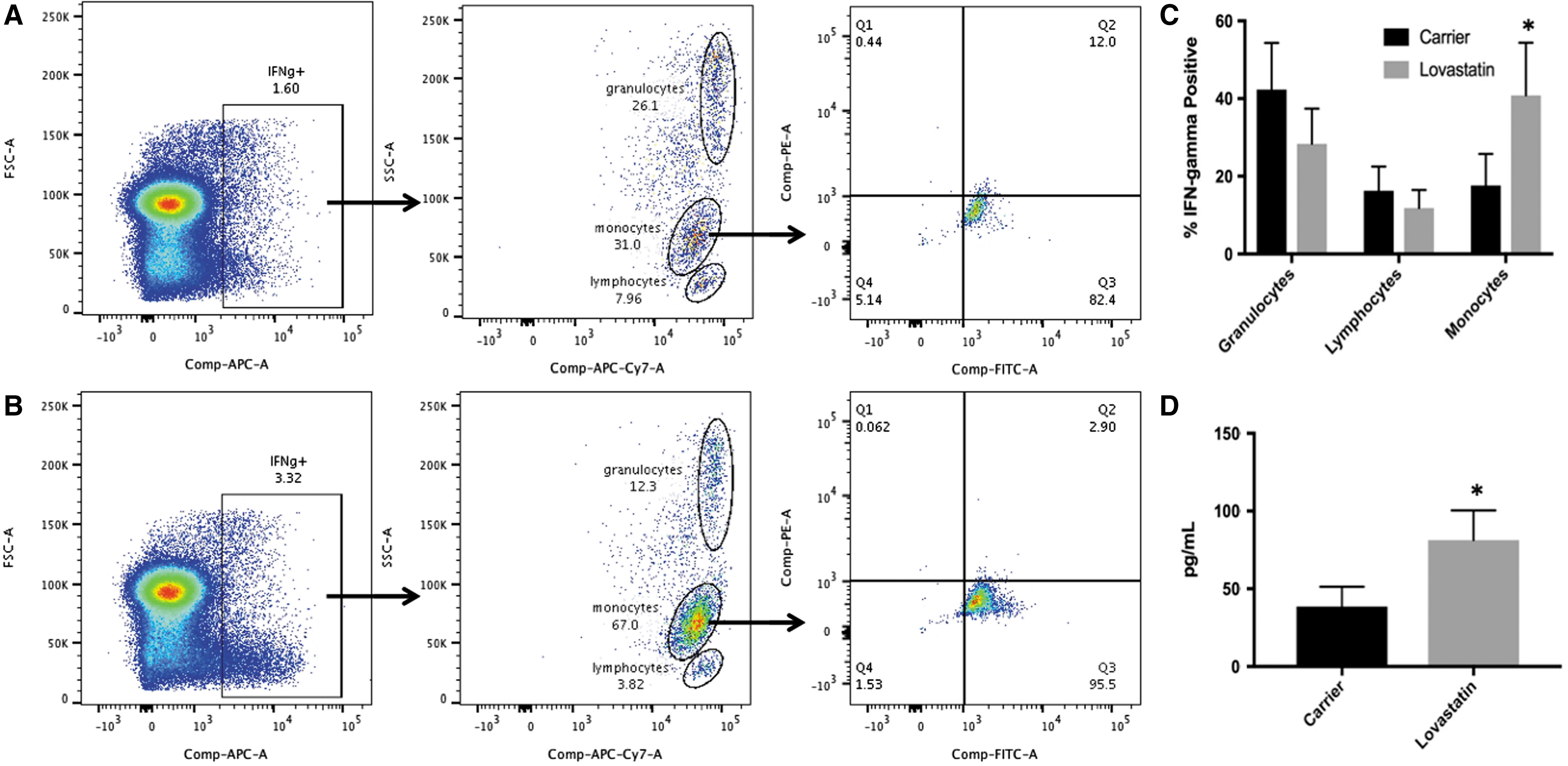
Lovastatin increases the percentage of IFN-γ-positive monocytes. PBMCs (n = 3) were treated with lovastatin for 16–18 h followed by stimulation with LPS for an additional 24 h. Cells were stained for IFN-γ (APC), CD14 (APC-Vio770), CD14 (FITC), and CD16 (PE) and analyzed by flow cytometry. The percentage of monocytes (comprised classical and nonclassical/intermediate populations) expressing IFN-γ was significantly higher in lovastatin compared with carrier-treated cells
Discussion
Patient data indicate that MKD is a multicytokine disease, and previous work has shown that blocking the mevalonate pathway leads to increased expression levels of proinflammatory cytokines, including IL-1β, TNF-α, IL-6, and IL-12p40 in mouse BMDMs and primary PBMCs from healthy donors (Akula and others 2016). The experiments presented in this article confirm results that isoprenoid depletion increases TNF-α, IL-6, and IL-12p40 mRNA levels after LPS stimulation in primary PBMCs and expand upon those findings by showing that isoprenoid depletion leads to increased IL-12 production by monocytes after LPS stimulation leading to increased IFN-γ release from monocytes. To our knowledge, increased IFN-γ levels have not been previously reported in a cell culture-based MKD model system. This is likely due, in part, to the fact that common cell lines used to study MKD, such as THP-1 cells, do not produce IFN-γ under similar conditions. It has also been reported that primary BMDMs express IFN-γ, while macrophage cell lines such as RAW 264.7 and J774 do not express IFN-γ after stimulation with IL-12 and IL-18 (Munder and others 1998). It should also be noted that previous work has found differences in how the RAW 264.7 murine macrophage cell line responds to isoprenoid depletion compared to primary PBMCs. CD14 mRNA and protein levels are increased in RAW 264.7 macrophages following isoprenoid depletion and LPS stimulation (Frey and De Maio 2007), whereas no differences in CD14 mRNA levels were observed in the primary PBMCs used in these experiments. This difference may be due to the fact that RAW 264.7 cells are differentiated macrophages as opposed to monocytes. Additional experiments with primary human monocyte-derived macrophages may resolve this contradiction. In addition, it is important to note that there was a high level of donor-to-donor variability in IFN-γ mRNA levels following lovastatin treatment and LPS stimulation. IFN-γ mRNA levels were measured on a total of 15 lovastatin-treated PBMC samples throughout this article with an average fold change of 104.5 ± 176.95 compared with carrier-treated cells 24 h following LPS stimulation. Among these samples, only 1 sample did not exhibit increased IFN-γ in lovastatin-treated cells. The remaining samples can be divided into high and low responders with 8 samples falling below a 50-fold change (range 1.9–33.4-fold) and 6 samples above a 50-fold change (range 56.5–620.2-fold).
IFN-γ is typically classified as a Th1 cytokine primarily expressed by T and NK cells in response to IL-12 and IL-18. However, there is an accumulating body of evidence that both mouse and human monocytes and macrophages express IFN-γ under various conditions (Munder and others 1998; Darwich and others 2009; Robinson and others 2010; Kraaij and others 2014; Metcalf and others 2017), including following LPS stimulation of primary human monocytes (Kraaij and others 2014; Metcalf and others 2017). Our results also indicate that human primary monocytes express IFN-γ following LPS stimulation. In addition, staining of monocytes with CD14 and CD16 to differentiate subsets by flow cytometry indicated that both CD16− and CD16+ monocytes were positive for IFN-γ expression. Furthermore, isoprenoid depletion caused by blocking the mevalonate pathway with lovastatin augmented IFN-γ mRNA levels in monocytes following LPS stimulation in an IL-12-dependent manner. IL-12 and IL-18 act together to increase IFN-γ mRNA levels in T cells and macrophages (Munder and others 1998; Darwich and others 2009; Robinson and others 2010). Statins have been previously reported to increase IFN-γ levels in CD4 and CD8 T cells following mitogen activation, which was dependent on IL-18 and IL-12 (Coward and others 2006). Our results indicate that IL-12p40, but not IL-18 mRNA, levels are increased in primary human monocytes following LPS stimulation and that an IL-12 neutralizing antibody completely blocks the increased IFN-γ levels observed after lovastatin treatment. While this does not rule out a role for IL-18, it does indicate that the increased IL-12 expression following lovastatin treatment is necessary for the subsequent increase in IFN-γ mRNA levels.
The exact mechanism by which isoprenoid depletion causes increased mRNA levels of IL-12p40 and other cytokines has yet to be determined. Our work indicates that the lovastatin-induced changes in gene expression are specific to blocking the mevalonate pathway since coincubation of lovastatin with mevalonate completely prevents the observed gene expression changes. Previous work indicates that that blocking geranylgeranylation of the Ras family member Kras in mouse BMDMs leads to reduced activation of the p110δ subunit of PI3K with a subsequent reduction in the phosphorylation of downstream targets, including Akt and GSK3β, following TLR4 activation by LPS (Akula and others 2016). These events result in increased transcription of the proinflammatory cytokines TNF-α, IL-12p40, and IL-6. Future work must be done to determine if GGPP depletion and subsequent effects on protein prenylation and PI3K signal transduction are responsible for our results in PBMCs.
We propose a model in which monocyte isoprenoid depletion increases LPS signal transduction leading to increased IL-12p40 mRNA (8 h following LPS stimulation) and subsequent IL-12 release from monocytes, which then acts on all monocyte subsets to increase IFN-γ mRNA levels and subsequent protein secretion at a later time point (24 h following LPS stimulation). These results may be impactful for MKD patients who do not respond to IL-1β blockade therapy. A recent study indicated that only 35% of MKD patients had a complete response to the anti-IL-1β monoclonal antibody canakinumab (versus 6% for placebo) after 16 weeks of treatment with only 23% maintaining disease control on an extended dosing regimen after 16 weeks (De Benedetti and others 2018). These results highlight the need for alternative MKD treatments. In addition, this work sets the stage for additional studies that examine the underlying mechanisms responsible for increased expression of multiple cytokines, including IFN-γ, TNF-α, and IL-6, which could lead to the development of therapies that target multiple cytokines.
Footnotes
Acknowledgments
RT-PCR primer sequences were provided by Faoud Ishmael, MD, PhD and the majority of work on this project was completed in his laboratory at Penn State College of Medicine. We thank Dr. Ishmael for the use of laboratory space, equipment, and expertise related to this project. We also acknowledge the contribution of students in the Fall 2017 Biology 433 class at Dickinson College for generating THP-1 data included in this article. The acquisition of the flow cytometry data was supported by the Penn State College of Medicine Flow Cytometry Core Facility.
Author Disclosure Statement
No competing financial interests exist.