
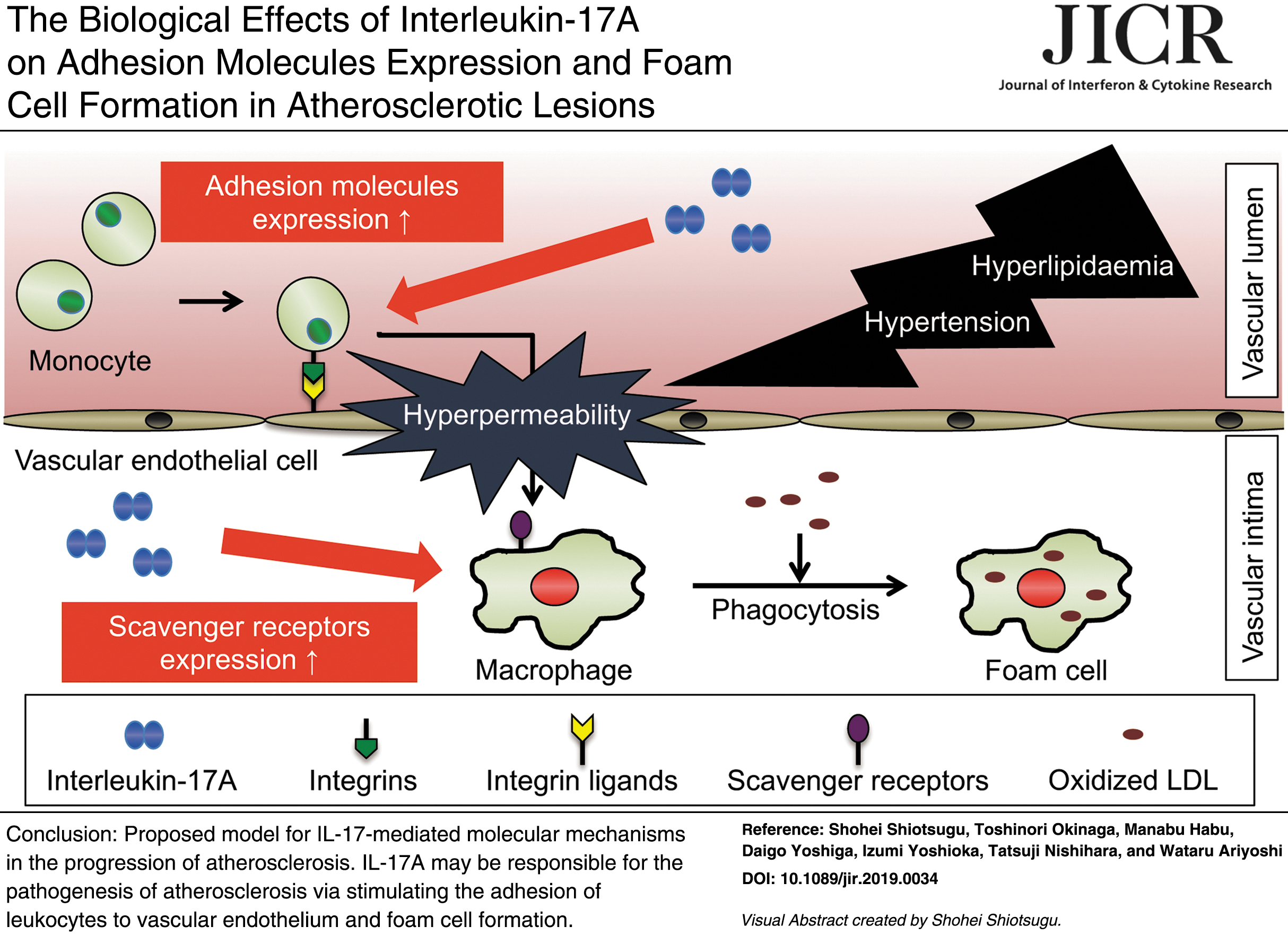
Abstract
Interleukin-17A (IL-17A), a major effector cytokine secreted by T helper 17 (Th17) cells, is elevated in atherosclerosis lesions. The purpose of this study was to assess the role of IL-17A in the pathogenesis of atherosclerosis. To measure the expression of adhesion molecules, human umbilical vein endothelial cells (HUVECs) and U937 cells were stimulated with IL-17A. Western blot and real-time polymerase chain reaction analyses revealed that intercellular adhesion molecule-1 (ICAM-1) and vascular cell adhesion molecule-1 (VCAM-1) expression in HUVECs, and very late antigen-4 (VLA-4), lymphocyte function-associated antigen-1 (LFA-1) and macrophage-1 antigen (MAC-1) expression in U937 cells was upregulated by IL-17A. Furthermore, IL-17A stimulation resulted in mRNA and protein expression of scavenger receptor (LOX-1) in phorbol 12-myristate 13-acetate (PMA)-activated U937 cells. Oil Red O also demonstrated that IL-17A enhanced foam cell formation by PMA-activated U937 cells induced by oxidized low-density lipoprotein (ox-LDL), and this enhancement of ox-LDL-induced foam cell formation in IL-17A-treated U937 cells was downregulated by transfection of LOX-1 siRNA. These results indicated that IL-17A induced the expression of adhesion molecules, promoted the adherence of monocytes to vascular endothelial cells. IL-17A also stimulated ox-LDL-induced foam cell formation via upregulation of LOX-1 in activated macrophages. IL-17A may be responsible for the pathogenesis of atherosclerosis by inducing the adhesion of leukocytes to vascular endothelium and foam cell formation.
Introduction
Atherosclerosis is a progressive vessel disease that is characterized by the accumulation of lipids and formation of fibrous plaques in the artery (Lusis 2000), and is responsible for a large proportion of cardiovascular-related morbidity and mortality worldwide (Liu and others 2016). Vascular endothelial cells are critical for cardiovascular homeostasis by maintaining an antithrombotic, anti-inflammatory, and antiatherogenic state within the vessel wall (Thomas and others 2008). Many studies have indicated that chronic inflammatory process is critical for the development of atherosclerosis and suggests that understanding the role of specific inflammatory factors involved in atherosclerosis will lead to more effective treatments and guidelines for suppressing the risk of cardiovascular events (Koh and Park 2018).
Atherosclerotic plaque formation is initiated by the interaction between adhesion molecules on leukocytes and on the endothelium, which leads to the accumulation of leukocytes on the artery wall (Ley and others 2007). Among the identified endothelial adhesion molecules, the biological properties and circulating expression levels of E-selectin, P-selectin, intercellular adhesion molecule-1 (ICAM-1), and vascular cell adhesion molecule-1 (VCAM-1) have been well described (Hansson and Libby 2006). Selectins are involved in the adherence of circulating monocytes to the endothelial wall (capturing) and their movement along the endothelium (rolling), which are the first 2 steps in the adhesion cascade during endothelial inflammation. However, transmembrane glycoproteins, ICAM-1 and VCAM-1 are responsible for transmigrating monocytes into the vascular intima.
In the process of atherosclerosis progression, formation and accumulation of lipid-laden macrophages (foam cells) within the lipid-rich subendothelial space of the affected artery play an important role in the inflammatory response (Angelovich and others 2015). Foam cell formation is associated with hyperlipidemia and abnormal accumulation of oxidized low-density lipoprotein (ox-LDL). Several scavenger receptors for ox-LDL have been identified, such as SR-A (Kodama and others 1990) and CD36 (Endemann and others 1993), both of which function primarily in macrophages, and lectin-like oxidized LDL receptor (LOX-1) (Sawamura and others 1997). Scavenger receptors bind multiple ligands and have been implicated in a variety of physiological and pathological processes, including endocytosis, phagocytosis, adhesion, and signaling (Prabhudas and others 2014).
The atherosclerotic plaque formation process is mediated by a variety of factors including inflammatory cytokines. Interleukin-17A (IL-17A) is a member of the IL-17 family and is produced mainly by T helper 17 (Th17) cells (Kolls and Linden 2004). IL-17A is a potent proinflammatory cytokine and modulates immune cell trafficking, mainly by inducing chemokines, thereby initiating inflammation and cytokine production in several diseases. The elevated expression of IL-17A leads to excessive inflammation and plays a pathological role in several inflammatory, infectious, and autoimmune diseases (Fossiez and others 1996). In addition, accumulating data showed increased IL-17A expression in atherosclerotic tissues (Erbel and others 2011), suggesting that IL-17A may be linked to the vulnerability of human atherosclerotic lesions.
Although several studies have assessed the association between IL-17 and susceptibility to many cardiovascular diseases (Robert and Miossec 2017), the involvement of IL-17A in the pathogenesis of atherosclerosis has not been clarified. In this study, we used an in vitro culture system to assess the role of IL-17A in the expression of adhesion molecules and in foam cell formation in the atherosclerotic plaque formation process.
Materials and Methods
Reagents and antibodies
Recombinant human IL-17A and anti-human LOX-1/OLR1 monoclonal antibodies were purchased from R&D Systems (Minneapolis, MN). Anti-human VCAM-1 and anti-human ICAM-1 monoclonal antibodies were obtained from Santa Cruz Biotechnology (Dallas, TX). Anti-human CD11b monoclonal, anti-human CD11a polyclonal and anti-human integrin alpha 4 polyclonal antibodies were purchased from Abcam (Cambridge, UK). Human ox-LDL was purchased from Biomedical Technologies, Inc. (Stoughton, MA).
Cell culture
Human umbilical vein endothelial cells (HUVECs) were purchased from Lonza, Inc. (Walkersville, MD) and expanded in endothelial growth medium-2 (EGM-2; Lonza, Inc., Allendale, NJ) supplemented with human epidermal growth factor, vascular endothelial growth factor, insulin-like growth factor, ascorbic acid, hydrocortisone, human fibroblast growth factor β, heparin, and fetal bovine serum (FBS) at 37°C in an atmosphere of 5% CO2. Cells below passage 3 were used for further experiments.
The human monocytic cell line, U937 (JCRB Cell Bank, Osaka, Japan), was maintained in RPMI 1640 (Gibco Laboratories, Grand Island, NY) supplemented with 10% heat-inactivated FBS (Corning, Corning, NY), 100 U/mL penicillin G potassium salt (Nacalai Tesque, Kyoto, Japan), and 100 μg/mL streptomycin (Wako Pure Chemical Industries, Osaka, Japan) at 37°C in an atmosphere of 5% CO2. For some experiments, U937 cells were seeded at 1.0 × 106 cells/mL in 6-well plates (Iwaki, Chiba, Japan) and cultured in RPMI 1640 containing 5% FBS in the presence of 100 ng/mL phorbol 12-myristate 13-acetate (PMA; Sigma-Aldrich, St. Louis, MO) overnight. Cultured cells were washed with RPMI 1640 without PMA, incubated in RPMI 1640 for 24 h, and then stimulated with IL-17A for the indicated durations.
Quantitative real-time reverse transcriptase–polymerase chain reaction analysis
Total RNA was extracted from cultured cells using Cica Geneus RNA Prep Kit (Kanto Chemical, Tokyo, Japan) according to the manufacturer's instructions. Extracted RNA was reverse transcribed with ReverTra Ace qPCR RT Master Mix (Toyobo, Osaka, Japan) and cDNA was amplified using a Thermal Cycler GeneAtlas (Astec, Tokyo, Japan). Quantitative polymerase chain reaction (PCR) was performed using an AriaMx Real-Time PCR System (Agilent Technologies) using Brilliant III Ultra-Fast SYBR Green QPCR Master Mix With Low ROX (Agilent Technologies, Santa Clara, CA). Relative changes in gene expression were calculated by the comparative CT (ΔΔCT) method. Total cDNA abundance in samples was normalized to β-actin mRNA expression.
The primers used for real-time reverse transcriptase (RT)-PCR were as follows: human VCAM-1, forward 5′-GATTGGTGACTCCGTCTCATT-3′ and reverse 5′-CCTTCCCATTCAGTGGACTATC-3′; human ICAM-1, forward 5′-GTAGCAGCCGCAGTCATAAT-3′ and reverse 5′-GGGCCTGTTGTAGTCTGTATTT-3′; human CD11b, forward 5′-GGAGTCATTCGCTACGTCATT-3′ and reverse 5′-TAAGCTCTTGGCGGGATTTC-3′; human Integrin alpha 4, forward 5′-ACTACCCGTGGGTCTTTATTTC-3′ and reverse 5′-TTGTACCACGCCAGAGTTATC-3′; human CD11a, forward 5′-ATT CCTGCCTGTCTCCTTTG-3′ and reverse 5′-TGGTGAACCGTCTGGTAATG-3′; human LOX-1/OLR1, forward 5′-GTCCTTTGCCTGGGATTAGTAG-3′ and reverse 5′-GGTTTGCTTGCTCTTGTGT TAG-3′; human SR-A1, forward 5′-TTTGCTTCCTCCGAATCCTAAA-3′ and reverse 5′-CCAATGAGAGGGATGAGAACTG-3′; human SR-B1, forward 5′-CATCTACCCACCCAACGAAG-3′ and reverse 5′-GAAGTGAGGATGGGAGAGAAAC-3′; human CD36, forward 5′-CATTGGTGATGAGAAGGCAAAC-3′ and reverse 5′-CACCACACCAACACTGAGTAA-3′; human CXCL16/SR-PSOX, forward 5′-CTG TGGCACCTGACTCTAATAC-3′ and reverse 5′-CTTGACTCAGGAGTTCCATAACA-3′; and human β-actin, forward 5′-GCGCGGCTACAGCTTCA-3′ and reverse 5′-CTTAATGTCACGCACGATTTCC-3′.
Western blot analysis
After treatments, total protein was extracted using sodium dodecyl sulfate (SDS) lysis buffer (50 mM Tris-HCl and 2% SDS, pH 6.8) supplemented with a protease inhibitor mixture (Nacalai Tesque). Protein content was measured using the DC protein assay reagent (Bio-Rad, Hercules, CA). Twenty micrograms of total protein were subjected to electrophoresis on SDS-polyacrylamide gels (Kishida Chemical, Osaka, Japan), followed by electroblotting onto polyvinylidene difluoride membranes (PALL Corp., Port Washington, NY). The membranes were blocked for 1 h with 5% skim milk at room temperature. The membranes were subjected to overnight incubation with diluted primary antibodies at 4°C. After washing with Tris-buffered saline containing 0.1% Tween 20 (TBS-T), they were incubated with horseradish-peroxidase (HRP)-conjugated anti-mouse and anti-rabbit IgG (GE Health Care, Little Chalfont, UK) antibodies for 1 h at room temperature. After washing with TBS-T, chemiluminescence was produced using the ECL Prime Western Blotting Detection Reagent (GE Health Care) and detected digitally with the GelDoc XRS Plus system (Bio-Rad).
Adhesion assay
HUVECs (5.0 × 105 cells/mL) were seeded on a cell culture insert (pore size 3.0 μm; Greiner Bio-One, Kremsmünster, Austria) and incubated with or without IL-17A (50 ng/mL) for 6 h. Meanwhile, U937 cells (1.0 × 106 cells/mL) were also stimulated with or without IL-17A (50 ng/mL) for 6 h. Stimulated HUVECs and U937 cells were labeled fluorescently with green and red, respectively, using Cell Explorer™ Fixable Live Cell Tracking Kit (AAT Bioquest, Sunnyvale, CA) according to the manufacturer's protocol. The labeled HUVECs monolayer (culture on cell culture insert) was exposed to the labeled U937 cells and cocultured for 6 h to allow cell–cell adhesion. The insert containing HUVECs and the adherent U937 cells were washed and carefully cut out. Adherent cells were monitored by a BZ-9000 fluorescence microscope (Keyence, Osaka, Japan). Images were captured digitally in real-time and processed using BZ-II imaging software (Keyence).
Silencing of LOX-1 expression by specific small-interfering RNA
Small-interfering RNA (siRNA) was used to knockdown LOX-1 expression in U937 cells. Control siRNA and siRNA against human LOX-1 were obtained from Santa Cruz Biotechnology. PMA-activated U937 cells (5.0 × 105 cells/mL) were plated in 6-well plates. The cells were transfected with 10 nM siRNA using Lipofectamine RNAiMAX (Thermo Fisher Scientific) according to the manufacturer's instructions. The transfected cells were subsequently incubated before stimulation with IL-17A.
Oil Red O staining
PMA-activated U937 cells (5.0 × 105 cells/mL) were grown in 6-well plates and treated with IL-17A and ox-LDL. After treatment, cells were washed with PBS and fixed with 10% formalin for 1 h, gently washed with 60% isopropanol, and stained with 0.5% Oil Red O (Wako Pure Chemical Industries) staining solution (prepared in 60% isopropanol) for 15 min. The percentage of foam cells is expressed as the percentage of Oil Red O-positive cells to total macrophages.
Statistical analysis
All statistical analyses were carried out using JMP® software, version 10.0.2 (SAS Institute, Inc., Cary, NC). Data were expressed as the mean ± SD of 3 individual experiments and analyzed by one-way analysis of variance (ANOVA) followed by a suitable post hoc test (Dunnett's or Tukey's) for multiple comparison P < 0.05 was considered statistically significant.
Results
Effect of IL-17A on adhesion molecules expression in endothelial cells
First, we investigated the effect of IL-17A on cell adhesion in vascular endothelial cells in vitro. HUVECs were incubated in the presence of IL-17A at different concentrations. IL-17A induced a concentration-dependent increase in ICAM-1 and VCAM-1, reaching statistical significance at 20 ng/mL after 9 h of treatment (Fig. 1A). This IL-17A effect was also time dependent with maximum enhancement observed at 3 h (Fig. 1B, ICAM-1, 3.0-fold increase; VCAM-1, 7.5-fold increase). Furthermore, Western blotting showed substantial enhancement of ICAM-1 and VCAM-1 protein expression in HUVECs treated with IL-17A (Fig. 1C).
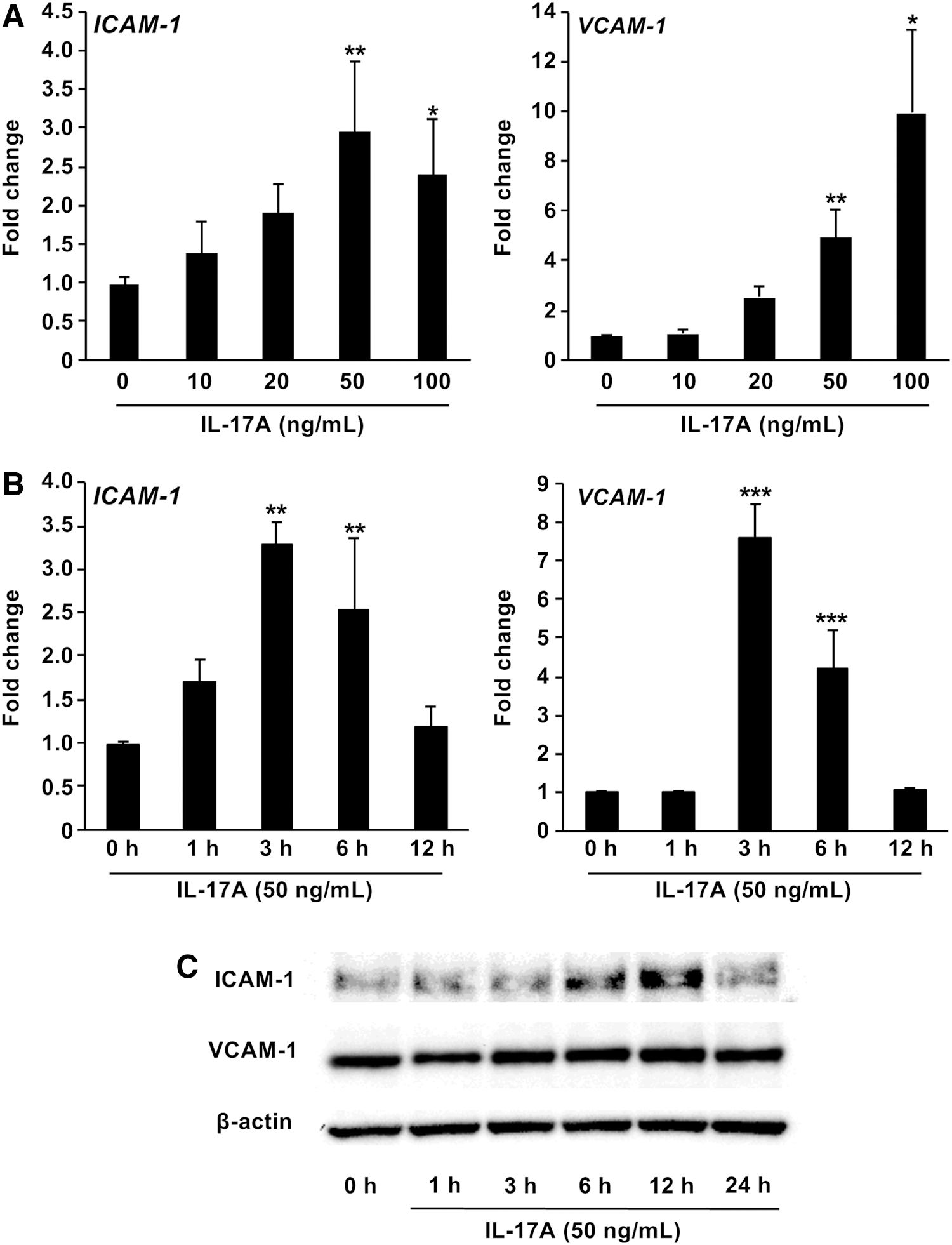
Effect of IL-17A on adhesion molecules expression in HUVECs
Effect of IL-17A on adhesion molecules expression in monocytes
Next, we investigated the effect of IL-17A on cell adhesion in monocytes in vitro. IL-17A stimulation resulted in transient enhancement of the mRNA (Fig. 2A) and protein (Fig. 2B) expression of lymphocyte function-associated antigen-1 (LFA-1), very late antigen-4 (VLA-4), and macrophage-1 antigen (MAC-1) in a time-dependent manner.
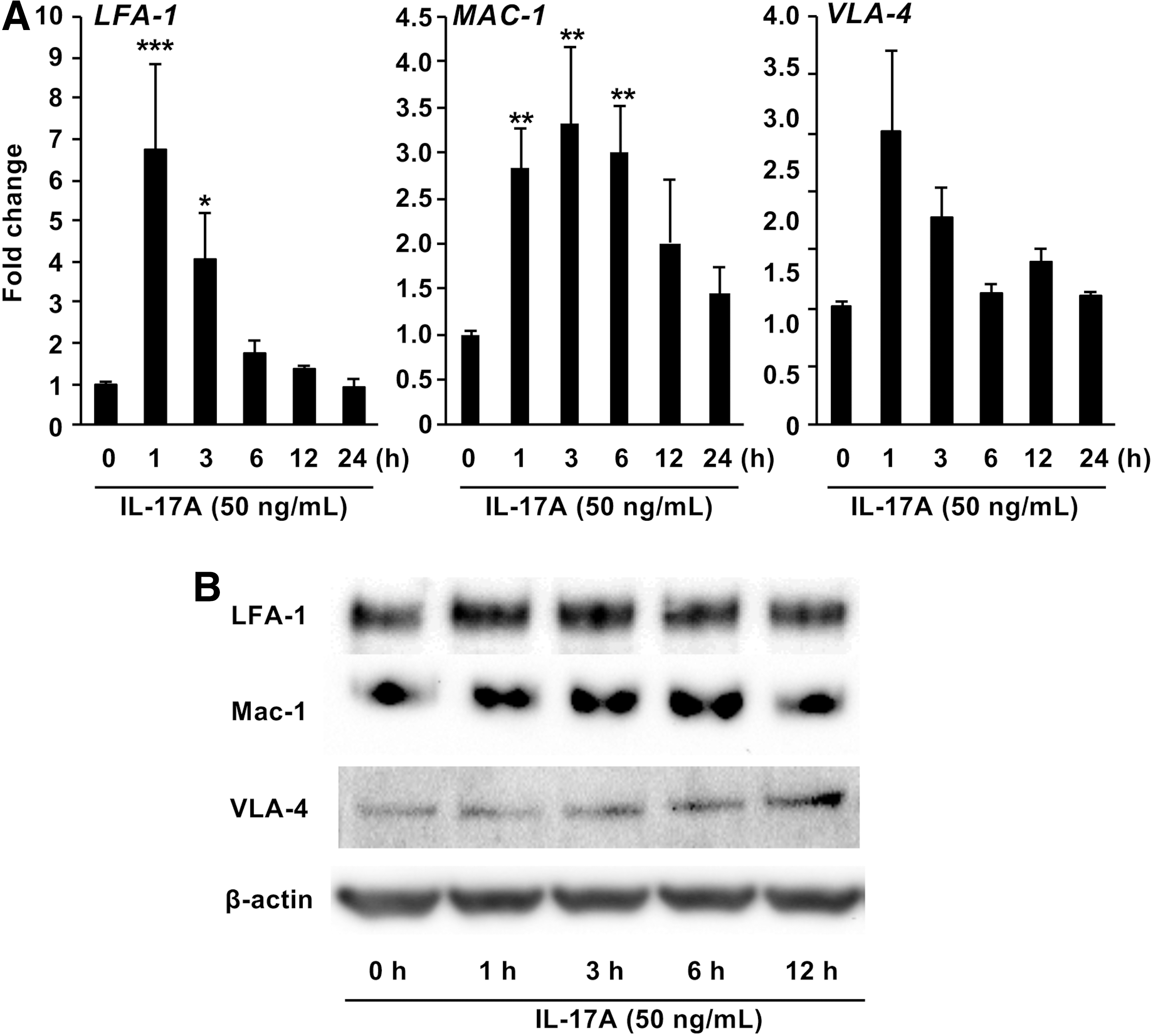
Effect of IL-17A on adhesion molecules expression in U937 cells
Effect of IL-17A on adhesion between HUVECs and U937 cells
To investigate whether IL-17A affects the adherence of monocytes to endothelial cells, we performed an adhesion assay using a coculture system in a cell culture insert (Fig. 3A). The number of U937 cells (red) surrounding HUVECs (green) was obviously increased in the IL-17A-stimulated group compared with the control group (Fig. 3B).
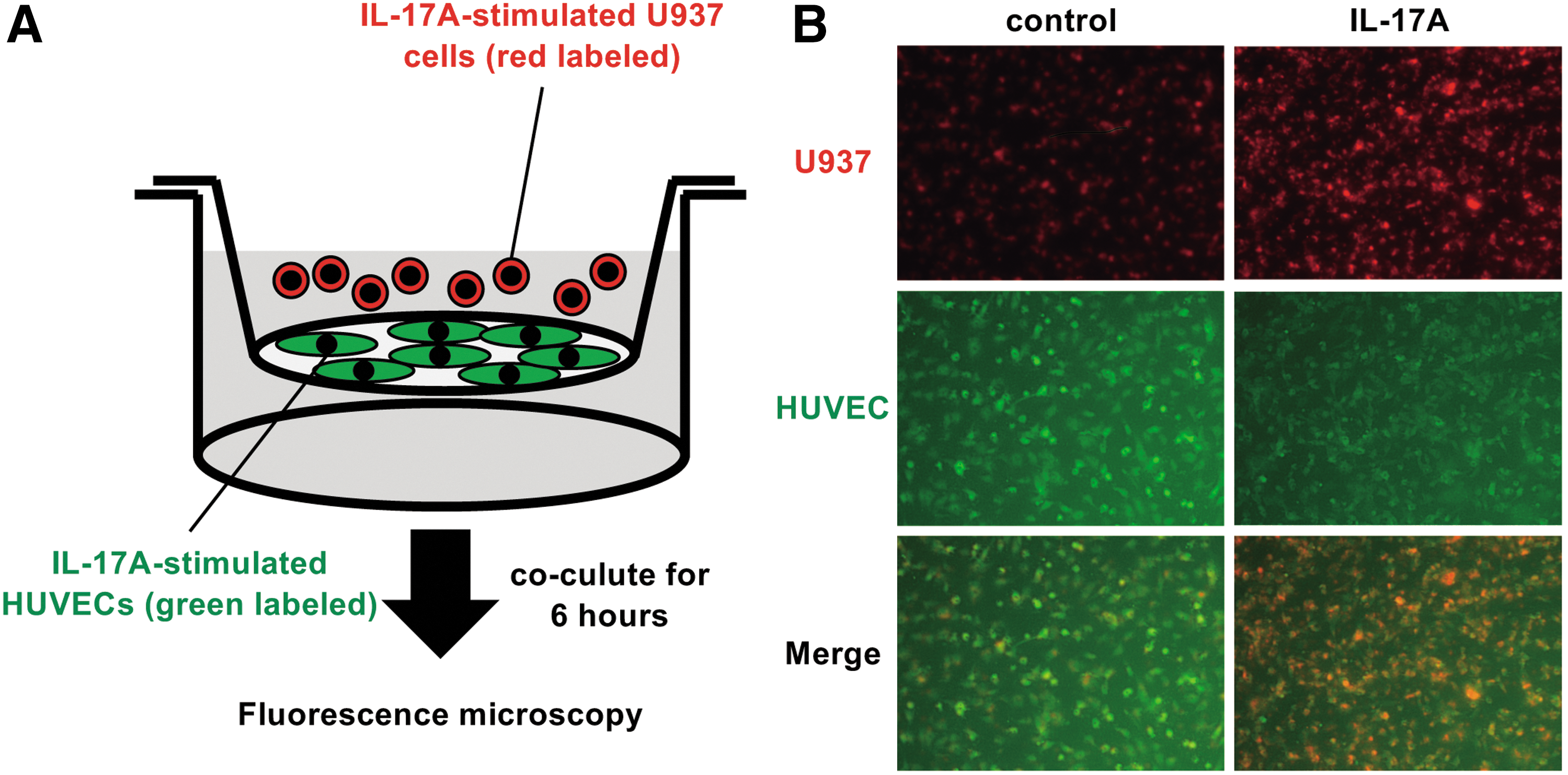
Effect of IL-17A on adhesion between HUVECs and U937 cells
Effect of IL-17A on scavenger receptors expression in macrophages
To investigate the effect of IL-17A on foam cell formation in macrophages in vitro, we determined the expression of scavenger receptors in PMA-activated U937 cells stimulated with IL-17A. IL-17A stimulation resulted in mRNA (Fig. 4A) and protein (Fig. 4B) expression of LOX-1 in a time-dependent manner. However, IL-17A stimulation had no effect on the expression of other scavenger receptors (Fig. 4A).
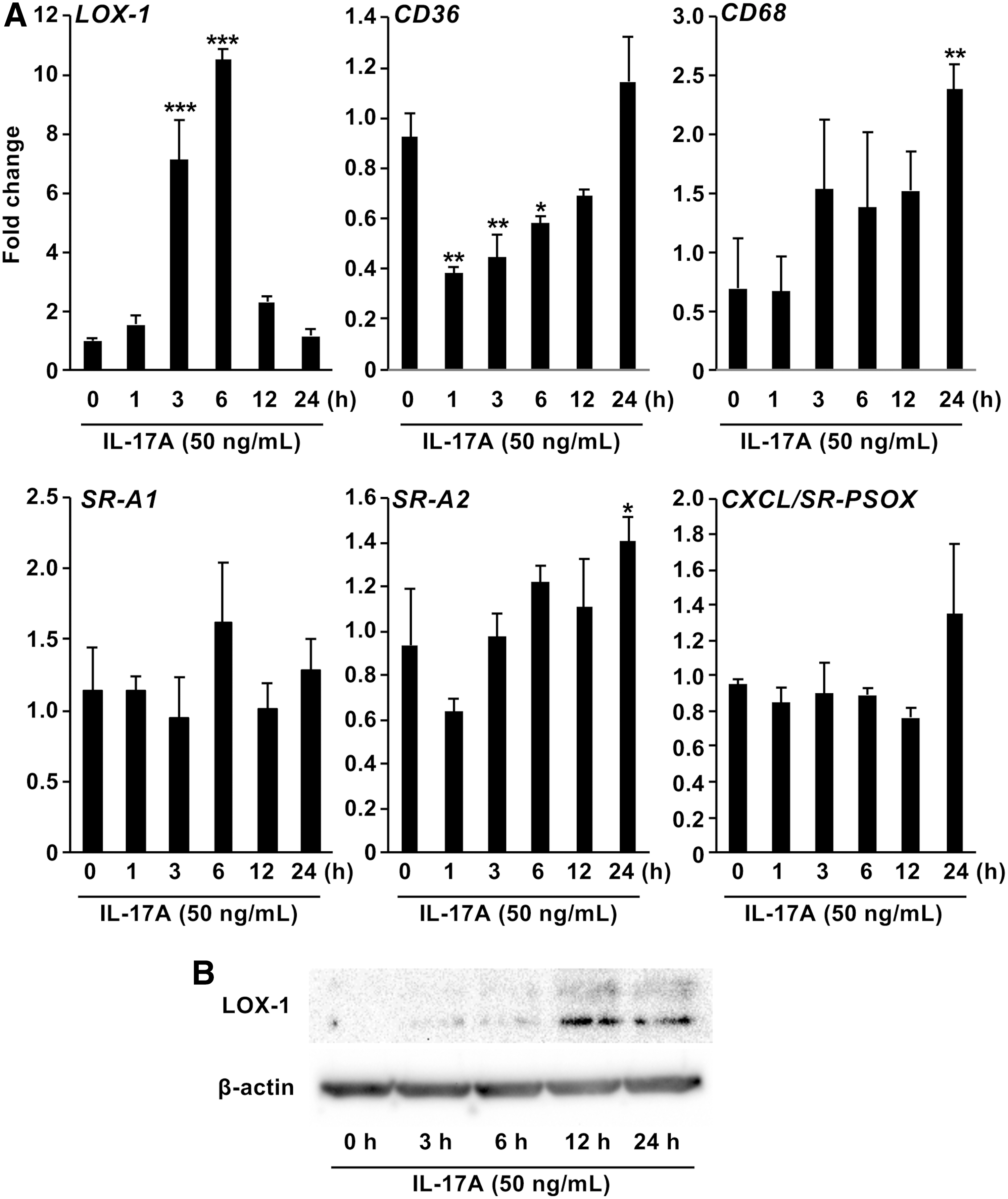
Effect of IL-17A on scavenger receptors expression in macrophages
Effect of IL-17A on foam cell formation mediated by LOX-1
We examined the effect of LDL on foam cell formation by PMA-activated U937 cells using Oil Red O staining. Treatment with ox-LDL clearly increased foam cell formation in a time-dependent manner up to 24 h (Fig. 5A). To elucidate the effect of IL-17A on the LDL-induced foam cell formation, we treated PMA-activated U937 cells with IL-17A and ox-LDL for 48 h. Treatment with IL-17A enhanced the ox-LDL-induced foam cell formation by PMA-activated U937 cells in a dose-dependent manner (Fig. 5B and C). To further examine the role of LOX-1 as a scavenger receptor in the induction of foam cell formation induced by IL-17A, PMA-activated U937 cells were transfected with LOX-1-specific siRNA. Transfection of PMA-activated U937 cells with LOX-1-specific siRNA effectively suppressed mRNA expression LOX-1 compared with a nonspecific control siRNA transfected cells (Fig. 5D). Furthermore, the IL-17A-induced increase in foam cell formation was suppressed in the presence of LOX-1 siRNA (Fig. 5E).
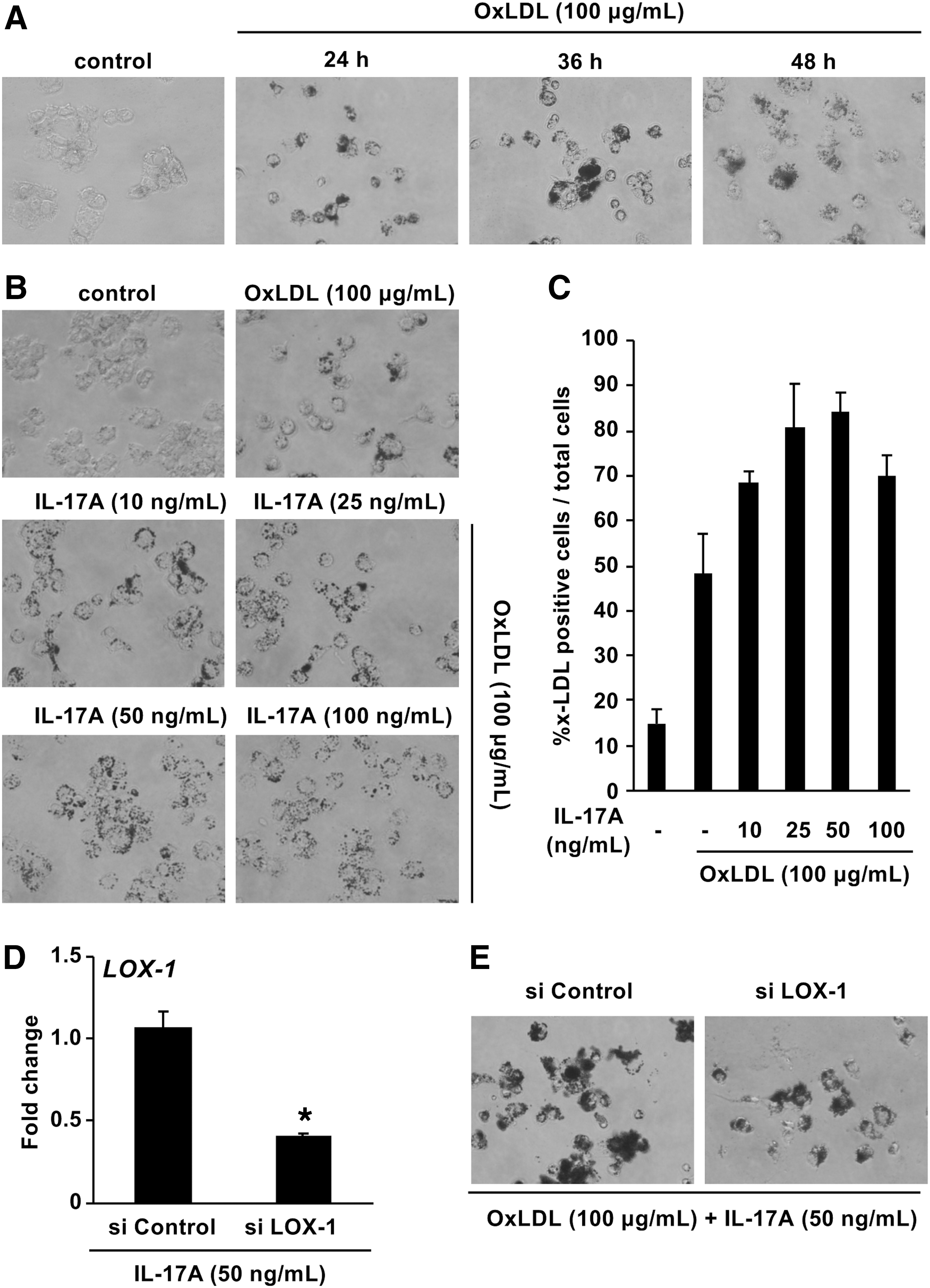
Effect of IL-17A on foam cell formation mediated by LOX-1
Discussion
Inflammation is a pivotal process in all stages of atherosclerosis. A number of inflammatory cytokines are involved in the pathogenesis of atherosclerosis and are considered as immunotherapeutic targets for modulation of atherosclerotic diseases. Inflammatory cytokines act on most of the cells involved in the pathogenesis of atherosclerosis and participate in all stages of the process from endothelial dysfunction to foam cell formation and plaque disruption (Tedgui and Mallat 2006). IL-17A belongs to the IL-17 family, originally thought to be produced by Th17 cells (Korn and others 2009). The major function of IL-17A in humans is being a proinflammatory factor, and numerous target cells of IL-17A including vascular endothelial cells, neutrophils, and macrophages have been shown (Onishi and Gaffen 2010; Numasaki and others 2016). Moreover, blocking IL-17A with monoclonal antibodies has been reported to attenuate cellular infiltration, expression of activation markers on the endothelium and immune cells, and cytokine/chemokine secretion in a mouse model of atherosclerosis (Erbel and others 2009). These findings implicate IL-17A in the pathogenesis of atherosclerosis. In the present study, we explored the role of IL-17A in adhesion molecules expression and foam cell formation in atherosclerosis.
Endothelial dysfunction is characterized by increased expression of vascular adhesion molecules on the endothelial cell surface and leukocyte/endothelium interactions, followed by promotion of atheroma initiation and progression (Pober 2002; Rao and others 2007; Weber and Noels 2011). The results of this study indicated that expression of adhesion molecules was stimulated in HUVECs in response to IL-17A. Consistent with these results, IL-17 stimulated human retinal vascular endothelial cell capillary tube formation via upregulation of ICAM-1 (Liu and others 2017). Furthermore, IL-17A stimulated adhesion molecules expression in U937 cells. From these results, we speculated that IL-17A induces leukocytes accumulation at the endothelial cells by upregulation of adhesion molecules expressed on both cell types. To study this speculation, we used an in vitro coculture model, which mimics the process of leukocyte adhesion in vivo, and showed that IL-17A stimulated the adherence of U937 cells to HUVECs. These data suggest that IL-17A induced leukocytes accumulation at the vessel wall by stimulating interactions between the adhesion molecules expressed on leukocytes and endothelial cells. Furthermore, ox-LDL is an important factor in endothelial injury (Liu and others 2014) and many studies have demonstrated that ox-LDL induces dysfunction of HUVECs via several signaling pathway including endothelial nitric oxide synthase (eNOS), phosphoinositide 3-kinase (PI3K), and protein kinase C-α (PKCα) (Simon and others 2014; Zhang and others 2019). Future studies were needed to determine the effect of IL-17A on ox-LDL-stimulated dysfunction and expression of vascular adhesion molecules.
Next, we focused on the effect of IL-17A on macrophage foam cell formation, which plays an important role in the promotion of plaque development within the arterial intima (Stary and others 1994; Galkina and Ley 2009). Macrophage foam cell formation has been reported to occur only through the uptake of modified forms of LDL, such as oxidized or aggregated LDL (Kruth 2001). PMA-derived macrophages from U937 cells exhibited foam cell formation when incubated with ox-LDL (Yao and others 2016). Ox-LDL in atherosclerotic lesions is mainly taken up by macrophages via scavenger receptor family members such as Class A (SR-A1), Class B (CD36, SR-B1), Class D (CD68), Class E (LOX-1), Class F (SREC-1), and Class G (SR-PSOX/CXCL16) (Febbraio and others 2000; Kunjathoor and others 2002; Moore and others 2005; Aslanian and Charo 2006; Mehta and others 2007; Manning-Tobin and others 2009). Unregulated uptake of OxLDL by scavenger receptors is crucial for the transformation of monocyte-derived macrophages to foam cells in atherosclerotic lesions.(Yamada and others 1998). Our results demonstrated that LOX-1 transcription and protein synthesis are stimulated in PMA-activated U937 cells in response to IL-17A.
LOX-1 is a multi-ligand scavenger receptor that has been identified as a receptor for ox-LDL uptake by vascular endothelial cells (Sawamura and others 1997, 2012). It is produced as a 40 kDa precursor protein with N-linked high mannose carbohydrate, and when further glycosylated it is processed to the mature form of 50 kDa (Kume and Kita 2001). LOX-1 is expressed in cells derived from atherosclerosis and is observed in large amounts in human and animal lesions (Kataoka and others 1999). In the present study, IL-17A induced foam cell formation by macrophages in the presence of exogenous ox-LDL. Based on these findings, we speculated that LOX-1, which is induced by IL-17A, is one of the important scavenger receptors responsible for the formation of foam cells in U937 cells. This speculation was clearly supported by the dramatic decrease in foam cell formation upon LOX-1 knockdown.
The binding of IL-17A via its correspondent receptor is reported to be involved in the biological effects of IL-17A. IL-17 receptor is a heterodimer, each presenting as a single pass transmembrane glycoprotein, which is ubiquitously expressed in a variety of cells, including vascular endothelial cells (Yang and others 2014) and monocytes (Suzuki and others 2014). The binding of IL-17A to its receptor induced the activation of downstream signaling molecules including mitogen-activated protein kinases and nuclear factor kappa B (Iwakura and others 2011). These signaling molecules have also been reported to be associated with the expression of adhesion molecules (Zhang and others 2018) and LOX-1 (Li and others 2004). Furthermore, combination of IL-17A and TNF-α has been reported to have a synergistic effect on chemokine expression by mRNA stabilization (Hartupee and others 2007). Th17 inflammatory process in atherosclerosis was also reported to be activated by Chlamydophila pneumoniae infection (Benagiano and others 2012) and in lupus erythematosus with antiphospholipid syndrome patients (Benagiano and others 2019). To fully elucidate the molecular mechanism of IL-17A function in the pathogenesis of atherosclerosis, the effects of activation and interaction of intracellular signaling by IL-17A on accumulation of leukocytes at the endothelial cells and foam cell formation are currently under investigation in our laboratory.
In summary, our findings provided valuable insight into the mechanism by which IL-17A may contribute to the pathogenesis of atherosclerosis by stimulating the accumulation of leukocytes at the artery wall and foam cell formation. These findings may provide a mechanistic explanation for the pathogenesis of atherosclerosis, and suggest that targeting the IL-17A signaling pathway is a novel promising strategy for treating patients with atherosclerosis.
Footnotes
Acknowledgments
We thank Edanz Group for editing a draft of this article.
Author Disclosure Statement
No competing financial interests exist.