
Abstract
Immunomodulatory effects of mesenchymal stem cells (MSCs) in inflammatory diseases, including psoriasis, are well documented. However, the role of MSC-derived exosomes (MSCs-Exo) in psoriasis-like skin inflammation remains largely unknown. This study aimed to investigate whether MSCs-Exo play a regulatory role in psoriasis-like skin inflammation. We found that subcutaneous injection of human umbilical cord MSCs-Exo (hucMSCs-Exo) significantly suppressed the proliferation of epidermis and reduced Psoriasis Area and Severity Index (PASI) scores in imiquimod (IMQ)-induced mice. hucMSCs-Exo also reduced the expression of interleukin (IL)-17, IL-23, and chemokine C-C-motif ligand 20 (CCL20) and inhibited phosphorylation of signal transducer and activator of transcription 3 (STAT3) in the skin of IMQ-induced mice and in human keratinocyte (HaCaT) cells. In addition, co-cultured with hucMSCs-Exo in vitro, the maturation and activation of dendritic cells (DCs) were suppressed, and the expression level of IL-23 was decreased. These results indicate that hucMSCs-Exo can effectively ameliorate psoriasis-like skin inflammation in mice by regulating the expression of IL-23 and IL-17, and inhibiting the maturation and activation of DCs. Our data offer a promising therapeutic approach for psoriasis by leveraging the immunomodulatory effects of hucMSCs-Exo.
Introduction
Psoriasis is a chronic immune-mediated inflammatory skin disease (Boehncke and Schon 2015; Deng and others 2016). Although the pathogenesis of psoriasis is not fully understood, interleukin (IL)-23 secreted by dendritic cells (DCs) and monocytes/macrophages is recognized to promote the expansion of IL-17-producing T cells, which exert pleiotropic effects on the recruitment and activation of other immune cells (Lowes and others 2014). The recent therapeutic success of anti-IL-23 and anti-IL-17 antibodies has shown that the IL-23/IL-17 axis plays a pivotal role in the inflammatory response in psoriasis, confirming that the IL-23/IL-17 axis is a good target for psoriasis treatment (Hawkes and others 2018).
Currently, psoriasis is mainly treated using topical therapies, phototherapy, acitretin, immunosuppressant drugs, immunomodulators, biologics, and phosphodiesterase 4 inhibitor. However, since the efficacy of conventional treatments for psoriasis is still unsatisfactory, newer approaches are urgently needed. Cell therapies have shown promising effects on the treatment of psoriasis (Chen and others 2016; Sah and others 2016). Mesenchymal stem cells (MSCs), the major stem cells used in cell therapy, have been used in clinical practice for more than 10 years and have been proven to be effective for the treatment of various intractable autoimmune and inflammatory disorders because of their immunomodulatory properties (Le Blanc and others 2008; Karussis and others 2010; Connick and others 2012; Khosrotehrani 2013). A previous preclinical study demonstrated that allogeneic MSC therapy has a beneficial therapeutic effect on psoriasis (Chen and others 2016; De Jesus and others 2016; Sah and others 2016). Recent studies have shown that the therapeutic benefits of MSCs were closely to related their exosomes (Lai and others 2010; Xin and others 2013; Liang and others 2014).
Exosomes are nanovesicles (50–200 nm) derived from multivesicular bodies, released by most living cells, and are considered to play an important role in transmitting information between cells (van Balkom and others 2019). Recent studies have shown that MSC-derived exosomes (MSCs-Exo) can recapitulate the therapeutic efficacy of MSCs, including anti-inflammatory effects (Zhang and others 2014a; Pachler and others 2017). In addition, MSCs-Exo have satisfactory curative effects on the treatment of spinal cord injury, liver fibrosis, and graft versus host disease (GVHD) (Huang and others 2017; Lou and others 2017). However, the role of MSCs-Exo in psoriasis-like skin inflammation remains largely unknown. Hence, this study aimed to investigate whether MSCs-Exo play a regulatory role in psoriasis-like skin inflammation.
Materials and Methods
Culture and identification of human umbilical cord MSCs
Human umbilical cord MSCs (hucMSCs) were kindly donated by the Central Laboratory of Liaocheng People's Hospital; the cell product has been certified by the National Institutes for Food and Drug Control of China (authorization No. SH201900594 and SH201900597). An extraction method was used as described previously (Zhang and others 2020b). The study was approved by the Ethics Committee of the Hospital Liaocheng People's Hospital, China. The cells were cultured in complete Dulbecco's modified Eagle's medium (Corning) containing 10% fetal calf serum (Gibco) at 37°C and 5% CO2. Third-passage hucMSCs were used in subsequent experiments. Morphological characteristics of hucMSCs were examined using an inverted microscope. The immunophenotypes, including CD29-FITC (TS2/16; eBioscience), CD44-FITC (IM7; eBioscience), CD105-PE (SN6; eBioscience), and HLA- DR-PE (TU36; eBioscience) of hucMSCs, were analyzed in a FACSCalibur equipped with Cell Quest software (BD Biosciences). Lipid differentiation and osteogenesis of hucMSCs were performed, described in a previous report (Zhang and others 2020a).
Oil red O staining
hucMSCs were seeded into 6-well plates. After lipid induction for 14 days, the cells were washed with phosphate buffer saline (PBS). Next, the cells were fixed with 10% formaldehyde for 40 min, and after washing with PBS, they were stained using Oil red O (ORO, (G1262; Solarbio) solution for 40 min. The working fluid was discarded, and the plates were washed with 60% isopropanol. The cells were then observed under an inverted microscope (Olympus, Tokyo, Japan).
Alizarin red staining
hucMSCs were seeded in 6-well plates. After induction of osteogenesis for 21 days, the cells were washed with PBS. Next, the cells were fixed with 10% formaldehyde for 15 min, and after washing with double distilled water, they were stained with alizarin red (G1038-100ML; Servicebio) for 20 min. Finally, the cells were washed again with double distilled water and observed under an inverted microscope (Olympus).
Cell culture and exosomes extraction
hucMSCs were cultured in exosome-free medium (C3801-0050; Shanghai Xiaopeng Biological Technology Co., Ltd.) for 48 h. The culture supernatant was collected (∼60 mL), centrifuged at 300g for 10 min, and then at 16,500g for 30 min, and later filtered through a 0.22 μm pore size filter to remove cell debris. At each of these steps, the pellet was discarded, and the supernatant thus obtained was used for the following step. The final supernatant was then ultracentrifuged at 100,000g for 70 min, and recentrifuged at the same speed. The purified exosomes were resuspended in PBS and prepared for identification.
Characterization of hucMSCs-Exo
Morphological characteristics of hucMSCs-Exo were observed employing transmission electron microscopy (TEM). The numbers and diameters of hucMSCs-Exo were detected using nanoparticle tracking analysis (NTA, ZetaView PMX 110; Particle Metrix, Meerbusch, Germany). Phenotypic profiles of the hucMSCs and hucMSCs-Exo were assessed by Western blotting (see Materials and Methods section) using antibodies (obtained from Abcam) specific for CD9 (1:2,000, ab92726), CD63 (1:1,000, ab134045), TSG101 (1:1,000, ab125011), and CD81 (1:1,000, ab109201) (Wang and others 2020). Protein content of hucMSCs-Exo was quantified using a BCA Protein Assay kit (Biyuntian P0010 500T).
Uptake experiment of exosomes
Freshly isolated hucMSCs-Exo were resuspended in PBS and labeled using the PKH26 Red Fluorescent Cell Linker Mini kit (MINI26-1 KT; Sigma-Aldrich, St. Louis, MO). Later, the suspension was centrifuged at 100,000g for 70 min, and the supernatant discarded. The labeled hucMSCs-Exo were resuspended in PBS and centrifuged once again at the same speed for washing the labeled hucMSCs-Exo. Next, the cells were resuspended in PBS and co-cultured with human keratinocytes (HaCaT) cells for 12 h. The hucMSCs-Exo were then separated from the HaCaT cells (ZQ0044; Shanghai Zhongqiao Xinzhou Biotechnology Co., Ltd.) by washing 3 times with PBS and fixed with 4% formaldehyde for 10–30 min. Subsequently, the cells were stained using 1 × phalloidin-iFluor 488 reagent, incubated for 20–90 min, washed with PBS, and finally DAPI was added to counterstain the nuclei. The cells were then observed under a confocal microscope (LSM800; Zeiss).
Animal model
Female C57BL/6 mice (8 weeks old) were randomly divided into 4 groups (n = 6 mice/group): control group (Control), imiquimod (IMQ) group (IMQ), PBS-treated IMQ group (PBS+IMQ), and hucMSCs-Exo-treated IMQ group (hucMSCs-Exo+IMQ). All experimental procedures were conducted in accordance with the Animal Use and Care Committee of Shandong University. To establish the IMQ model, the C57BL/6 mice were treated with a daily topical dose (62.5 mg) of 5% IMQ cream (Aldara; 3M, St Paul, MN) applied on shaved dorsal skin for 6 days as reported previously (Swindell and others 2017). Clinical scores were assessed by 3 independent researchers, who assigned scores of 0–4 (0, none; 1, mild; 2, moderate; 3, severe; and 4, very severe) for erythema, scaling, and thickness. On days 0, 2, and 4 (Fig. 3a), the mice in the hucMSCs-Exo+IMQ and PBS+IMQ groups were subcutaneously injected with exosomes (50 μg/mouse) and PBS (50 μL/mouse), respectively. All the mice were euthanized on day 7 for analysis. The dorsal skin of the mice was sectioned, and a part of the skin was stained with hematoxylin and eosin (H&E) for histological evaluation. The other tissues were used for Western blotting (see Materials and Methods section) to detect the levels signal transducer and activator of transcription (STAT)3/p-STAT3, IL-17, IL-23, and chemokine C-C-motif ligand (CCL20); GAPDH was used as an internal control. All experimental procedures were conducted in accordance with the Animal Use and Care Committee of Shandong University (KYLL-2019(KS)-095). There is no Clinical Trial Registration involved.
Histological evaluation
Skin sections from the dorsal surface of the mice were stained with H&E for histological evaluation. Psoriasiform epidermal hyperplasia was assessed microscopically in the sections as reported earlier (Nadeem and others 2015, 2017).
Generation and differentiation of bone marrow-derived dendritic cells and co-culture with hucMSCs-Exo
DCs were obtained from the bone marrow (BM) of 2 normal 4- to 6-week-old female C57BL/6 mice according to a previously described protocol (Zhang and others 2014b). BM cell pellets were resuspended in RPMI-1640 medium (Gibco) supplemented with 10% fetal bovine serum (Gibco), 1% penicillin-streptomycin, 20 ng/mL rmGM-CSF (PeproTech), and 20 ng/mL rmIL-4 (PeproTech) at 1 × 106 cells/mL and transferred to a 6-well plate. On day 3, nonadherent cells were gently removed, and fresh culture medium was added. Thereafter, fresh culture medium was added every 2 days to induce DC differentiation. For DC maturation, on day 7, nonadherent cells were collected, washed, resuspended at 2 × 105 cells/mL in a culture medium containing 10 ng/mL rmTNF-α (PeproTech), and seeded in a 24-well plate, with or without hucMSCs-Exo (2.5 μg/mL) and co-cultured for another 24 h. The different groups of DC cells were collected to detect their phenotypes by flow cytometry, and the cell supernatants were collected and centrifuged at 1,000g for 20 min at 4°C. The supernatants thus obtained were stored at −80°C and utilized to perform enzyme-linked immunosorbent assays (ELISAs) within 1 month.
HaCaT cells co-cultured with hucMSCs-Exo
HaCaT cells (ZQ0044; Shanghai Zhongqiao Xinzhou Biotechnology Co., Ltd.) were used as a cell model system. When the cells reached ∼90% confluency, they were seeded in 6-well flat-bottom plates (2 × 105 cells/mL). When the cells reached ∼80% confluency, they were starved overnight in serum-free and antibiotic-free medium and were then stimulated with 100 ng/mL IL-17A (PeproTech) with or without the addition of hucMSCs-Exo (2.5 μg/mL) for 24 h. The different groups of HaCaT cells were harvested, and cellular proteins were extracted for Western blot analysis to detect the expression levels of STAT3/p-STAT3, with GAPDH as an internal control. Cell culture supernatants were collected and centrifuged at 1,000g for 20 min at 4°C and stored at −80°C.
Western blot analysis
HaCaT cells and mouse skin tissues from each treatment group were collected and lysed in RIPA buffer containing protease and phosphatase inhibitors. Protein samples (30 μg/lane) were separated using 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to PVDF membranes (Millipore). The membranes were blocked with 5% bovine serum albumin in Tris-buffered saline with Tween solution (TBST) for 2 h at room temperature (RT) and incubated with primary rabbit monoclonal antibodies (procured from Abcam) specific for CD9 (1:2,000, ab92726), CD63 (1:1,000, ab134045), TSG101 (1:1,000, ab125011), CD81 (1:1,000, ab109201), STAT3 (1:1,000, ab68153), p-STAT3 (1:1,000, ab30647), IL-17A (2 μg/mL, ab79056), IL-23 (1 μg/mL, ab45420), and CCL20 (0.2 μg/mL, ab9829) at 4°C overnight. Next day, the membranes were incubated with peroxidase-coupled avidin goat anti-rabbit IgG (Cell Signaling Technology) at RT for 1 h. Then the membranes were scanned and protein levels were normalized to GAPDH (1:1,000, , ab8245; Abcam) as a control. The ChemiDoc Touch Gel Imaging System and Image Lab Touch software (Bio-Rad) were used to record and quantify the signal intensity.
ELISA analysis
Supernatants of cell cultures were analyzed by ELISA. The protein level of IL-23p40 (E-EL-M2451c; Elabscience) was evaluated in the DC culture supernatant. Protein levels of IL-23p19 (E-EL-H0107c; Elabscience) and CCL20 (E-EL-H0027c; Elabscience) were evaluated in the HaCaT cell culture supernatant. Absorbance at 450 nm was measured using an ELISA plate reader (Bio-Rad).
Flow cytometric analysis
Flow cytometric (FCM) analysis of DCs was performed by triple-color staining. Cells were incubated with the following fluorochrome-conjugated antibodies for 30 min at 4°C: CD11c-FITC (N418; eBioscience), CD86-APC (GL1; eBioscience), and MHCII-PE (M5/114.15.2; eBioscience). Next, the cells were centrifuged at 300g for 5 min at RT, washed, resuspended in PBS, and analyzed using a FACSCalibur equipped with Cell Quest software (BD Biosciences).
Statistical analysis
All data are presented as mean ± standard deviation. All experiments were performed at least 3 times. The differences between 2 groups were analyzed using Student's t-test. When ≥3 groups were compared, the differences were analyzed using 1-way analysis of variance with SPSS 17.0 analytical software (IBM, Armonk, NY) or GraphPad Prism 7.00 (GraphPad Software, La Jolla, CA), followed by Tukey's post hoc test. P < 0.05 was considered to be statistically significant.
Results
Identification and labeling of hucMSCs
After 72 h of incubation, hucMSCs showed a spindle-shaped appearance and grew in whorls when examined under an inverted microscope (Fig. 1a). They could be differentiated into multiple lineages, including osteogenic and adipogenic lineages, under lineage-specific culture conditions. Osteogenesis of hucMSCs was observed by alizarin red staining (Fig. 1a) and adipogenesis of MSCs was observed by ORO staining (Fig. 1a). Using FCM analysis, we found that MSCs expressed CD105, CD29, and CD44, but not HLA-DR (Fig. 1b).
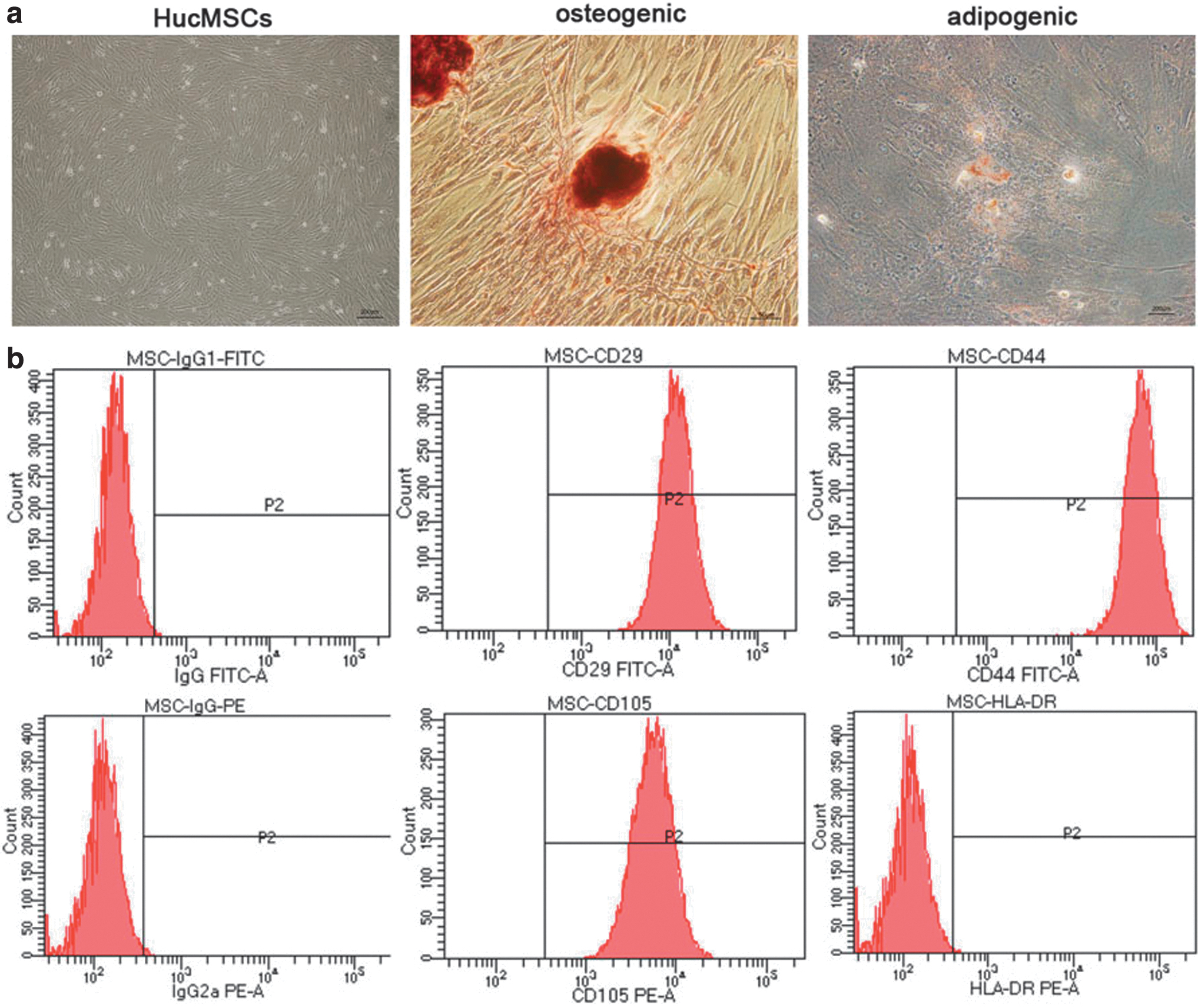
Typical characteristics of hucMSCs.
Characterization of hucMSCs-Exo
Using TEM, hucMSCs-Exo were observed as cup-shaped vesicles with diameters ranging from 50 to 200 nm (Fig. 2a). High expression levels of CD9, CD63, CD81, and TSG101 were detected in hucMSCs-Exo, but not in hucMSCs (Fig. 2b, Supplementary Fig S1–S5). Exosome distribution was performed by NTA (Fig. 2c). In addition, using confocal fluorescence microscopy, we provided qualitative evidence that HaCaT cells took up hucMSCs-Exo after co-culture for 12 h (Fig. 2d). These results indicated that we successfully extracted hucMSCs-Exo, and verified that they could be taken up by HaCaT cells after co-culture.
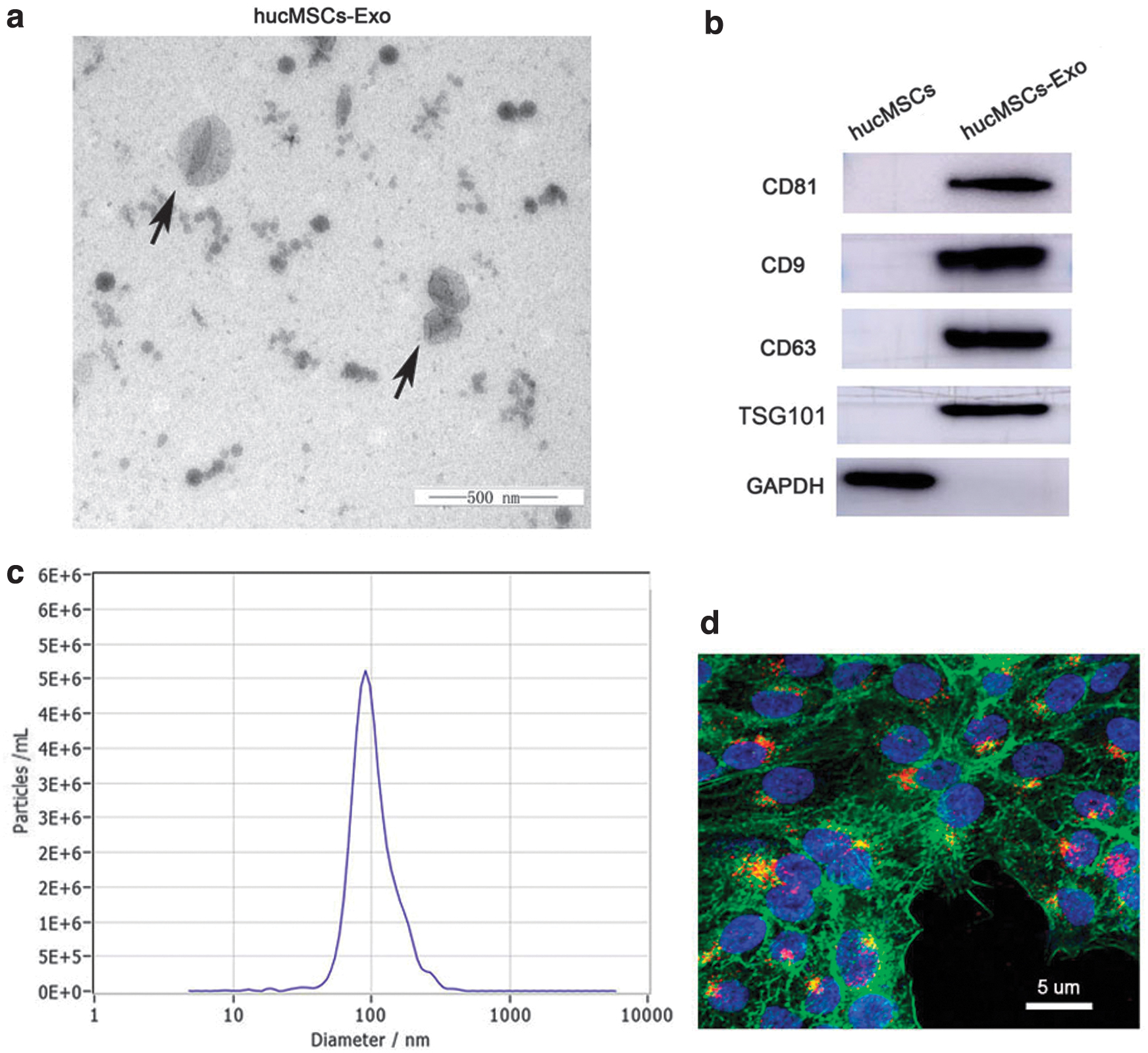
Characterization of hucMSCs-Exo.
hucMSCs-Exo ameliorated the severity of psoriasis-like symptoms in skins of mice
Mice in the IMQ-treated groups appeared to have signs of erythema, scaling, and thickening. H&E staining results showed increased epidermal thickness and inflammatory infiltration in the IMQ-treated group. Compared with the PBS+IMQ group, the hucMSCs-Exo+IMQ group exhibited reduced psoriatic erythema, scaling, and thickening (Fig. 3b). In addition, the Psoriasis Area and Severity Index (PASI) scores of the mice in the IMQ+hucMSCs-Exo group were significantly lower than those in the IMQ+PBS group (Fig. 3c), and H&E staining showed decreased epidermal thickness and inflammatory infiltration (Fig. 3d). These data suggested that hucMSCs-Exo ameliorated IMQ-induced psoriasis-like symptoms in skins of mice.
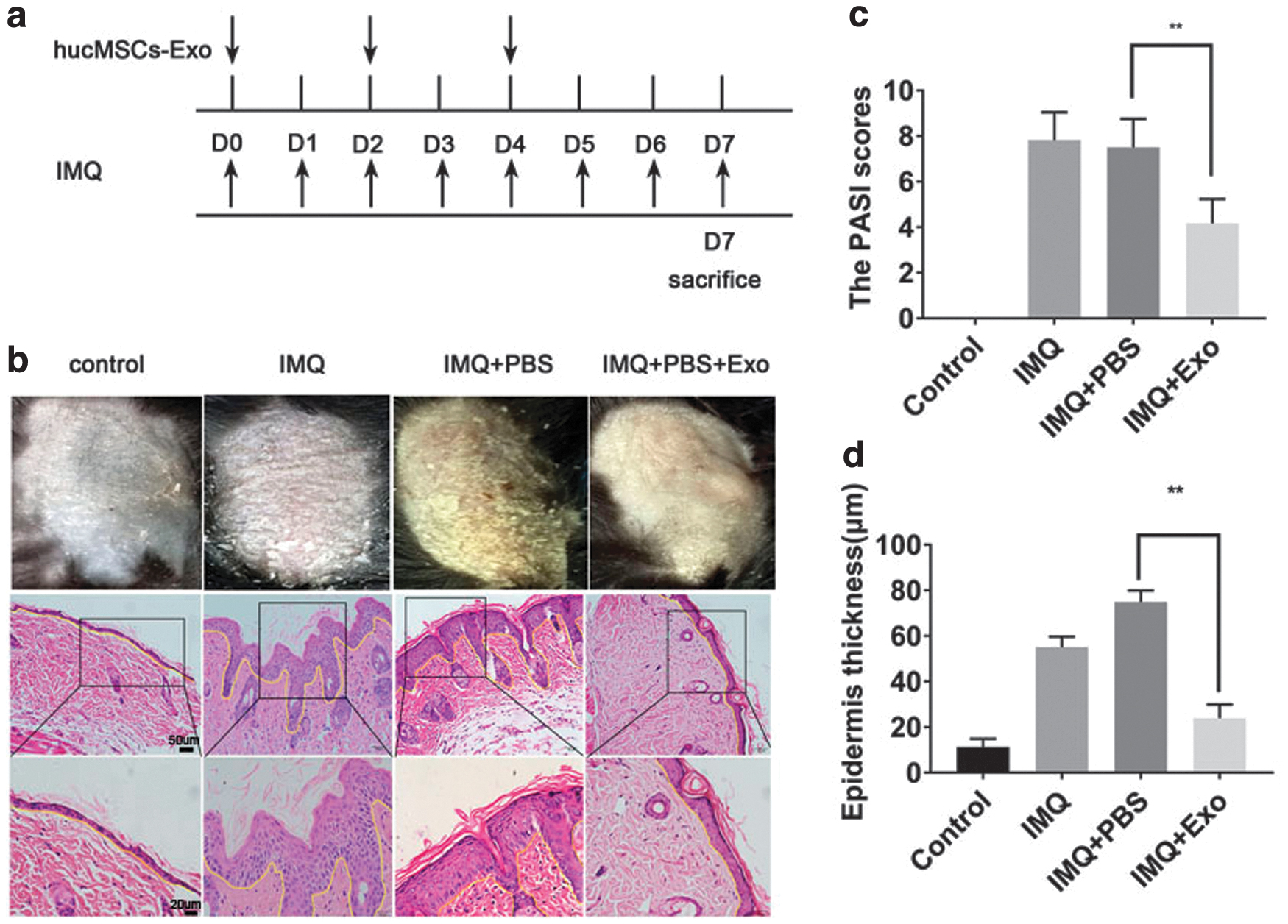
hucMSCs-Exo inhibited epidermal hyperproliferation, decreased acanthosis, and reduced the severity of IMQ-induced psoriatic skin inflammation.
hucMSCs-Exo inhibited the expression of IL-17, IL-23, and CCL20
Psoriasis is a chronic immune-mediated inflammatory skin disease; the IL-23/IL-17 cytokine axis plays a pivotal role in the inflammatory response in psoriasis. STAT3 has recently emerged as a key player in the development and pathogenesis of psoriasis and psoriasis-like inflammatory conditions (Calautti and others 2018). STAT3 is phosphorylated by receptor associated kinases, which later translocates to the cell nucleus where it acts as a transcription activator. Hence, we further tested the effects of hucMSCs-Exo on the expression of such inflammatory cytokines and chemokines. As shown in Fig. 4a–f and Supplementary Fig S9–S14, subcutaneous injection of hucMSCs-Exo effectively reduced their expression in hucMSCs-Exo+IMQ groups, which was consistent with the observed histological changes. These results implied that hucMSCs-Exo could inhibit psoriasis-like skin inflammation by suppressing the expression of IL-17, IL-23, and CCL20, thereby inhibiting the phosphorylation of STAT3.
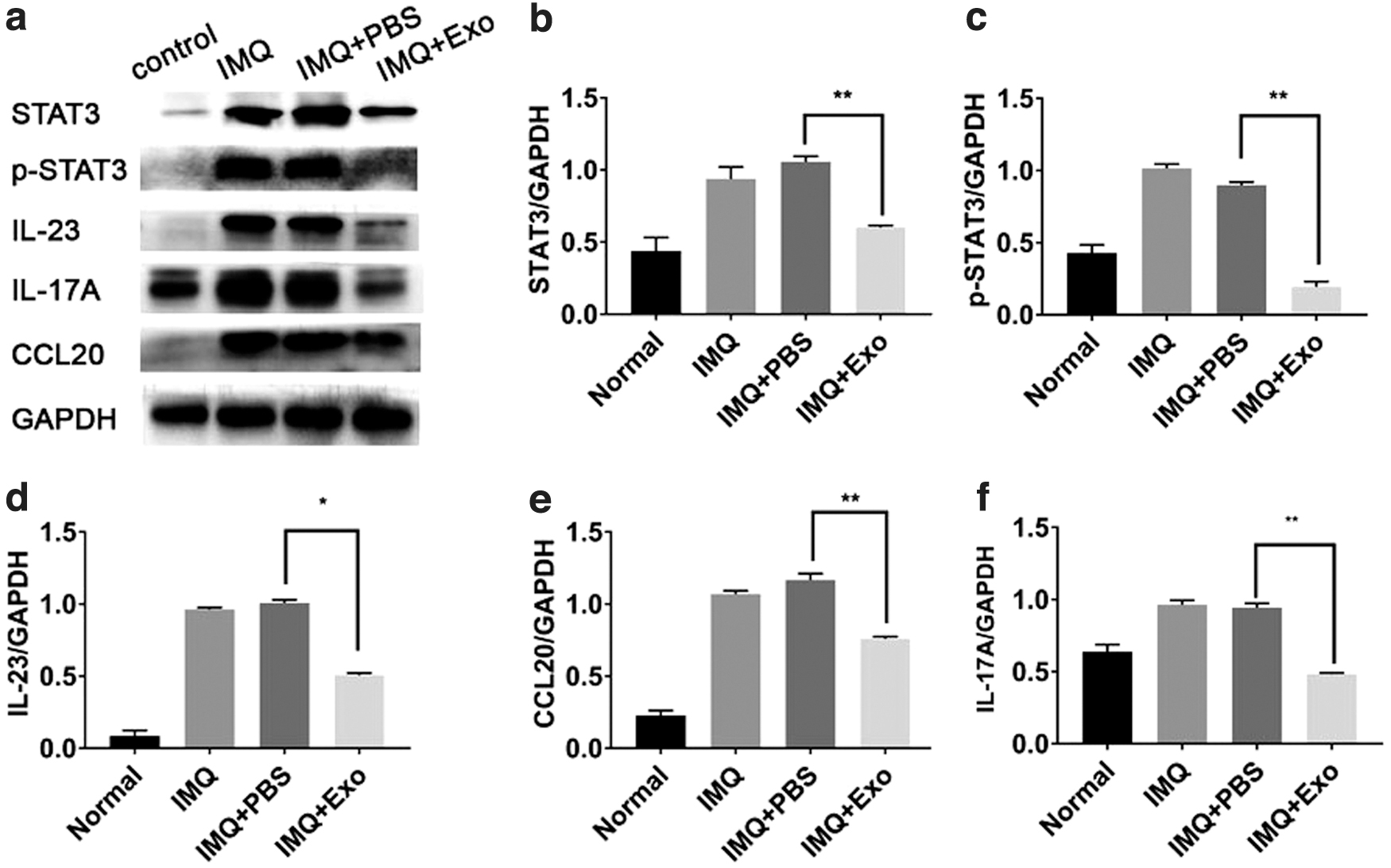
hucMSCs-Exo ameliorated IMQ-induced psoriasis-like skin inflammation in mice.
hucMSCs-Exo inhibited the maturation and activation of DCs
DCs play a pivotal role in the pathogenesis of psoriasis by regulating the differentiation and function of Th17 and Treg cells. Using FCM analysis, we found that co-culturing cells with hucMSCs-Exo versus without co-culturing with hucMSCs-Exo, the percentages of CD11c+MHCII+mDCs (59.15% ± 9.28 versus 64.95% ± 4.38%, respectively), CD11c+CD86+mDCs (47.53% ± 9.86 versus 59.86% ± 9.66%, respectively), and CD11c+MHCII+CD86+mDCs (56.83% ± 8.05 versus 44.70% ± 12.38%, respectively) were downregulated (Fig. 5a, b).
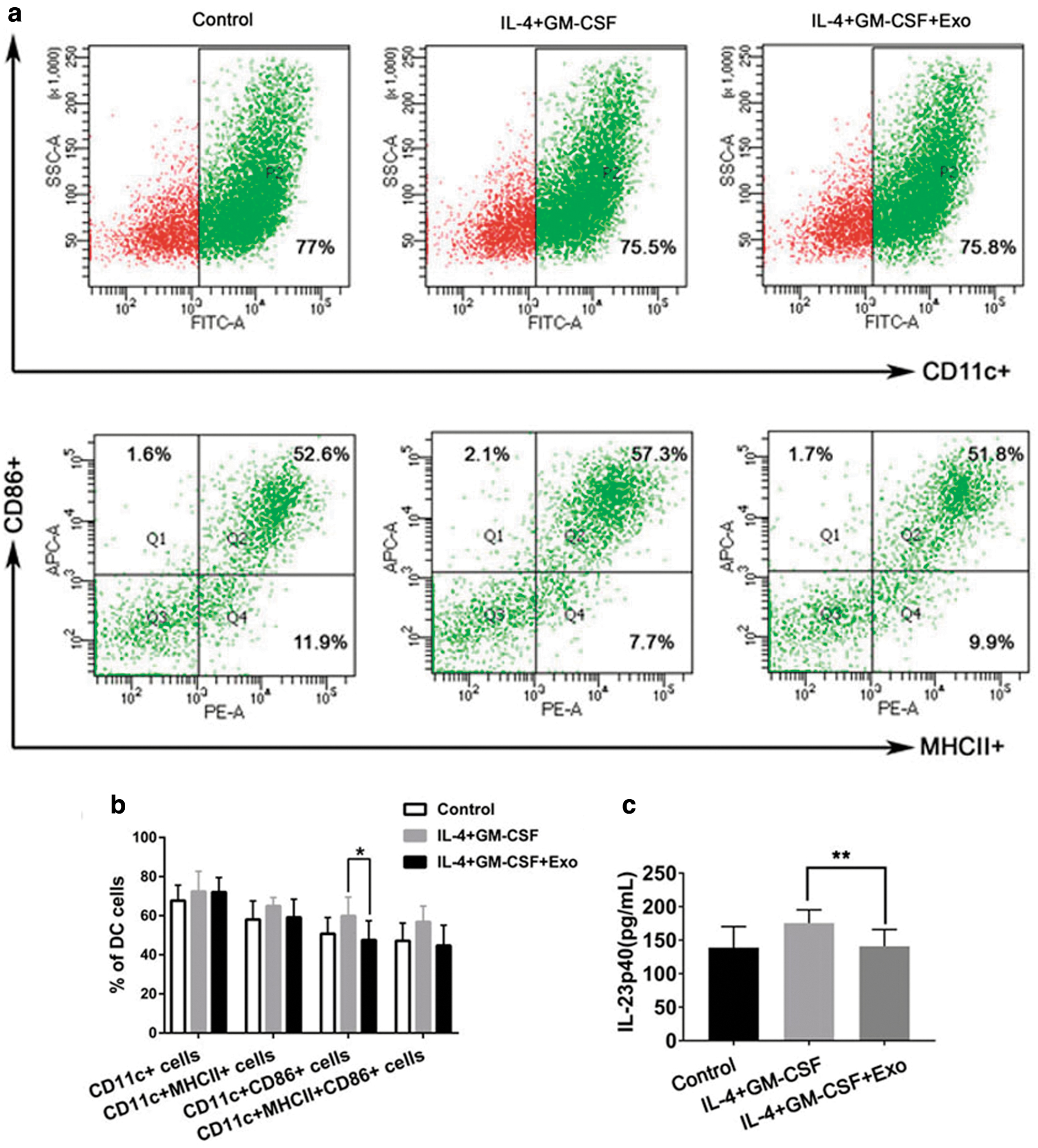
hucMSCs-Exo suppressed maturation and activation of DCs and decreased IL-23 secreted by DCs.
Next, we investigated whether hucMSCs-Exo were involved in regulating the expression of IL-23 secreted by DCs. Our results showed that the expression of IL-23 exhibited a decreasing trend after treatment with hucMSCs-Exo (Fig. 5c). These results indicated that hucMSCs-Exo not only inhibited the maturation and activation of DCs but also decreased the production of IL-23.
hucMSCs-Exo suppress IL-23 and CCL20 secretion of HaCaT cells by inhibiting STAT3 activity
Activated keratinocytes (KCs) play an important role in the inflammatory environment in psoriasis. HaCaT cells were treated with IL-17A and hucMSCs-Exo to assess their effect on cytokine and chemokine secretion. Using Western blot analysis and ELISA, we found that hucMSCs-Exo not only significantly reduced the expression and phosphorylation of STAT3 (Fig. 6a, b, Supplementary Fig. S6–S8) but also inhibited the expression of IL-23 and CCL20 secreted by HaCaT cells (Fig. 6c, d). These data indicated that the immunomodulatory effect of hucMSCs-Exo could be achieved by regulating the expression and phosphorylation of STAT3 and the secretion of IL-23 and CCL20.
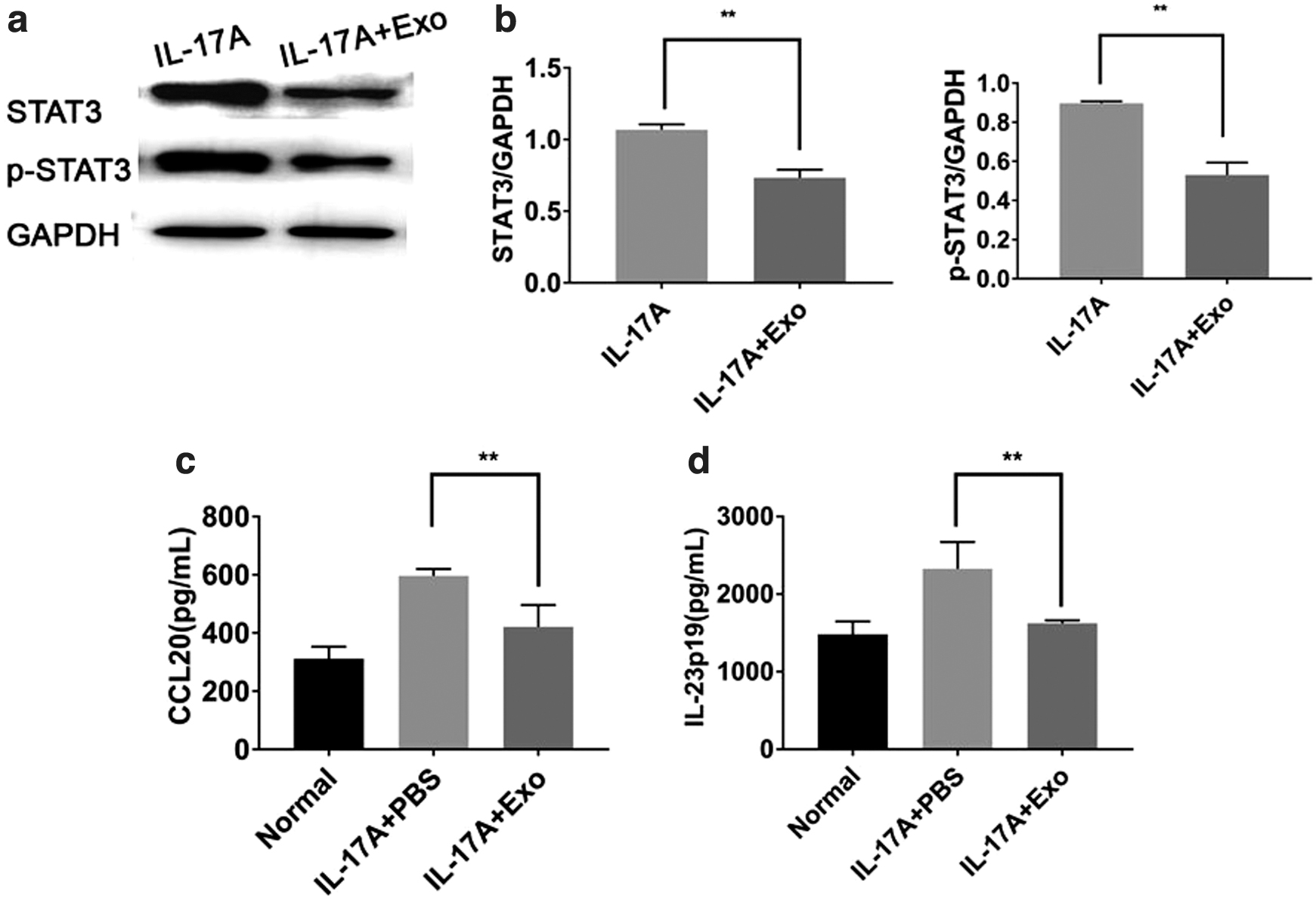
hucMSCs-Exo suppress IL-23 and CCL20 secretion of HaCaT cells by inhibiting STAT3 activity.
Discussion
Therapeutic effects of hucMSCs on psoriasis in experimental studies and clinical cases have been reported previously (Chen and others 2016; Sah and others 2016). HucMSCs have been considered to be a promising candidate for novel cell-free therapy (Ding and others 2015; Li and others 2015). MSCs-Exo have therapeutic effects on various chronic inflammatory disorders, such as inflammatory bowel disease, atopic dermatitis, and chronic GVHD (Mao and others 2017; Cho and others 2018; Lai and others 2018). Accumulating evidence suggest that MSCs-Exo exhibit potent immunomodulatory effects by regulating the activation of immune cells and inhibiting the expression of various inflammatory cytokines (Teng and others 2015; Li and others 2016).
Psoriasis is driven by interactions between activated immunocytes (such as DCs, Th1 and Th17) and KCs (Zaba and others 2009; Hawkes and others 2018). Recently, a new theory regarding the role of the IL-23/IL-17 axis in psoriasis has been increasingly validated (Sakkas and Bogdanos 2017; Hawkes and others 2018; Erichsen and others 2020). It is well known that (mature dendritic cells) can produce IL-23, which plays an important role in stimulating the proliferation and activation of Th17 cells, resulting in abnormal secretion of inflammatory cytokines such as IL-17, IL-21, and IL-22. These cytokines act on KCs, leading to epidermal hyperplasia, acanthosis, and hyperparakeratosis, which are key characteristics of psoriasis. In this study, we found that hucMSCs-Exo significantly alleviated the severity of psoriasis. The skin erythema, scaling, and epidermal thickness were significantly decreased. Our results demonstrated that subcutaneously injected hucMSCs-Exo significantly improved clinical symptoms, decreased inflammatory infiltration, and inhibited hyperplasia of the epidermis in IMQ-induced psoriatic mice.
DCs play a pivotal role in the pathogenesis of psoriasis. Based on a previous study, MSCs inhibited early DC maturation, and either MSCs or MSC-derived extracellular vesicles (MSC-EVs) co-cultured with DCs downregulated the secretion of IL-23 and IL-22 (Favaro and others 2016; Harrell and others 2019). Our results showed that hucMSCs-Exo not only inhibited the maturation and activation of DCs but also decreased the production of IL-23. In addition, treatment with hucMSCs-Exo decreased the expression of IL-23 in the skin of IMQ-induced psoriatic mice, which is consistent with our in vitro experimental results. Taken together, our results suggested that the effects of hucMSCs-Exo ameliorated psoriasis-like skin inflammation by inhibiting the functions of DCs and secretion of IL-23.
Increasing evidence has shown an overactive response of Th17 in psoriasis-like inflammation (Chang and others 2018). Activated KCs play an important role in the inflammatory environment of psoriasis by producing IL-23, which can synergistically mediate cross talk between Th17 lymphocytes and IL-23 secreted by mDCs. Moreover, KCs also secrete chemokines such as CCL20, which efficiently recruit Th17 cells into skin lesions. In this study, we observed that hucMSCs-Exo inhibited the secretion of IL-23 and CCL20 by HaCaT cells. Moreover, we further found that hucMSCs-Exo inhibited the expression of the IL-17, IL-23, and CCL20 in the skin of IMQ-induced psoriatic mice. Our results demonstrated that hucMSCs-Exo played a regulatory role in psoriasis, probably by regulating the secretion of proinflammatory cytokines and chemokines.
As a central regulator of inflammatory and immune responses, STAT3 mediates the IL-23/IL-17 axis, which is involved in the pathogenesis of psoriasis (Calautti and others 2018). Activated STAT3 is closely related to Th17 cell differentiation and the response of KCs to inflammation (Calautti and others 2018). Cui and others (2018) reported that exosomes derived from both MSCs and hypoxic MSCs prevented STAT3 activation. In our study, we found that hucMSCs-Exo inhibited phosphorylation of p-STAT3 in the epidermis of IMQ-induced psoriatic mice and in HaCaT cells, suggesting that immunomodulatory effects of hucMSCs-Exo could be achieved by regulating the expression and phosphorylation of STAT3.
The composition of hucMSCs-Exo is complex, and the specific component of hucMSCs-Exo that contributes to their biological activities is unknown. Indeed, MSC-EVs contain various MSC-derived proteins, lipids, DNA, messenger RNAs (mRNAs), and microRNAs (miRNAs); among them, miRNAs have received the most attention because they play an important role in regulating immune responses. Exosomes from preconditioned MSCs (PC-MSCs) have been shown to improve learning abilities and memory of APP/PS1 mice through the regulation of inflammatory responses by regulating miR-21 (Cui and others 2018). MiR-181c in EVs was found to be critical for immunoregulation and efficiently reduced inflammation in burned rats (Li and others 2016). Li and others (2019) found that MSCs-Exo reversed the increased levels of Th17/Treg in aplastic anemia through SphK1-mediated exosomal S1P enrichment. However, the precise mechanisms of action or key therapeutic factors related to hucMSCs-Exo in psoriasis have not yet been fully uncovered. This study is limited to only 1 model of psoriasis; however, increasing emphasis should be placed to study psoriasis multiple models. Moreover, our research on KCs is limited to a study on HaCaT KCs, but is not performed in primary murine or human KCs.
In conclusion, our findings suggest that the regulatory role of hucMSCs-Exo in psoriasis is by the inhibition, maturation, and activation of DCs, regulation of expression and phosphorylation of STAT3, and secretion of cytokines and chemokines. Our results also suggest that hucMSCs-Exo might be a promising agent for the treatment of psoriasis.
Ethics Approval and Consent to Participate
MSCs were kindly donated by the Central Laboratory of Liaocheng People's Hospital. HaCaT cells (ZQ0044; Shanghai Zhongqiao Xinzhou Biotechnology Co., Ltd.) were supplied by the Shanghai Zhongqiao Xinzhou Biotechnology Co., Ltd. Animal experiments were approved by the Research Ethics Committee of Qilu Hospital of Shandong University [No. KYLL-2019(KS)-095].
Availability of Data and Materials
The datasets generated and/or analyzed during this study are available from the corresponding author on reasonable request.
Footnotes
Authors' Contributions
Conception and design of the study: Q.S. Implementation of experiments: Y.Z. and J.Y. Analysis and interpretation of data: J.Y. and Z.L. Drafting of the article: Y.Z., J.Y., and J.Z. Final approval of the article: all the authors have agreed to submit this article.
Acknowledgments
We would like to acknowledge the helpful comments on this article received from our reviewers. We also thank the staff of the laboratory of Basic Medical Sciences for their help during the experiment, and Editage (
Author Disclosure Statement
We declare that there is no conflict of interests.
Funding information
This work was supported by the National Major Science and Technology projects of China (Grant No. 2017YFA0104604), National Natural Science Foundation of China (Grant Nos. 81773323, 81972937 and 82003344), Health Science and Technology Development plan project of Shandong Province (2019WS117), and Liaocheng People's Hospital Hospital-level Youth Research Fund (LYQN201918) and the Medical.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
Supplementary Figure S10
Supplementary Figure S11
Supplementary Figure S12
Supplementary Figure S13
Supplementary Figure S14
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.