
Abstract
Lycopene is one of the major carotenoids and is found almost exclusively in tomatoes and tomato products. This study was performed to evaluate the effect of lycopene on methanol-induced liver injury and to compare the results with those after fomepizole, which is used in treatment of methanol intoxication. Experiments were carried out with 30 female Wistar rats weighting 180–200 g. Rats were injected with a intraperitoneally dose of 3 g/kg methanol as a 50% solution in isotonic saline once for intoxication. Rats were pretreated with fomepizole (50 mg/kg) and/or lycopene (10 mg/kg) before methanol. After 24 hours all the drug-treated and intoxicated rats were sacrificed under anesthesia. Malondialdehyde (MDA) levels were determined in order to assess lipid peroxidation, and caspase-3 activity was determined by immunostaining of liver tissues to evaluate apoptosis. Methanol administration significantly increased the MDA level and caspase-3 activity in liver. Pretreatment with lycopene and/or fomepizole decreased the MDA levels significantly. Similarly, lycopene and fomepizole decreased methanol-induced caspase-3 activity. The findings of the present study demonstrate that methanol intoxication causes hepatic toxicity in rats and that this is likely a result of reactive oxygen species and apoptosis induction. Lycopene has protective effects against methanol-induced hepatic injury similar to fomepizole. It was demonstrated for the first time that both lycopene and fomepizole prevent methanol-induced hepatic injury by reducing the increase of lipid oxidation and caspase-3 activation.
Introduction
L
Methanol is extensively used as an industrial solvent and cleaner. It is also ingested as a substitute for ethanol when ethanol is either too expensive or unavailable. 9 Methanol has a relatively low toxicity, and its metabolism is responsible for the transformation of methanol to its toxic metabolites. In mammalian species, methanol is oxidized by alcohol dehydrogenase to formaldehyde in the liver. The oxidation of formaldehyde to formic acid is facilitated by formaldehyde dehydrogenase. Formic acid is converted by 10-formyl tetrahydrofolate synthetase to carbon dioxide and water. 10 For the methanol-poisoned patient without evidence of clinical toxicity, the first priority is to inhibit methanol metabolism with intravenous ethanol or fomepizole. 11
Acute ethanol intoxication induces oxidative stress and apoptosis in primary cultured 12 and in vivo 13 hepatocytes. Similarly, ethanol intoxication triggers widespread apoptotic neurodegeneration in the infant rat or mouse brain and T cells in humans. 14,15 McVicker et al. 16 have noted that ethanol administration leads to an increase (45–60%) in caspase-3 activity, causing the activation of apoptosis in hepatocytes, and that the induction of apoptosis was found to be linked to the metabolism of alcohol. Oxidative stress can trigger mitochondrial cytochrome c release, initiating the mitochondrial pathway of apoptosis. 14 However, methanol consumption can lead to a variety of pathological consequences by as yet undefined mechanisms. Formic acid and methanol have common mechanisms of toxicity because formic acid is a metabolic end product of methanol and is mainly responsible for the toxic inhibition of cytochrome c oxidase. Inhibition of the cytochrome c oxidase complex leads to anaerobic glycolysis and lactic acidosis-“histotoxic hypoxia.” 17 It was hypothesized that because of acidosis, the generation of oxygen radicals might be enhanced, leading to membrane damage, lipid peroxidation, and mitochondrial damage. 18 –20
Therefore, this study was conducted to demonstrate whether methanol leads to apoptosis with the oxidative stress and to investigate the possible protective effect of lycopene against methanol-induced damage in liver and compare it with fomepizole, which is used in treatment of methanol intoxication. We evaluated the levels of malondialdehyde (MDA) and activation of caspase-3 enzyme by immunostaining to assess lipid peroxidation and apoptosis, respectively. Methanol-induced histopathological alterations in liver were also determined.
Materials and Methods
Animals
Experiments were carried out with 30 female Wistar rats weighting 180–200 g. They were placed in a quiet and temperature (21 ± 2°C) and humidity (60 ± 5%) controlled room in which a 12/12-hour light-dark cycle was maintained. The animals were housed five per cage and fed with standard pellet diet and water ad libitum. All experiments in this study were performed in accordance with the National Institutes of Health guidelines 21 and were approved by the Committee on Animal Research at Firat University, Elazig.
Rats were randomized acording to their weights by simple random sampling. Animals were divided into six groups, each consisting of five animals as follows: (1) control group, injected intraperitoneally with 1.0 mL of saline; (2) lycopene group, injected intraperitoneally with a dose of 10 mg/kg lycopene; (3) methanol group, injected intraperitoneally with a dose of 3 g/kg methanol; (4) fomepizole + methanol group, injected intraperitoneally with a dose of 50 mg/kg fomepizole followed by a dose of 3 g/kg methanol; (5) lycopene + methanol group, injected intraperitoneally with a dose of 10 mg/kg lycopene followed by a dose of 3 g/kg methanol; and (6) fomepizole + lycopene + methanol group, injected with fomepizole and lycopene followed by methanol. The rats were given the same dose of lycopene 12 hours after intoxication. The rats were sacrificed 24 hours after methanol administration.
The dose of methanol causing 50% lethality in rats is 9.5 g/kg. Methanol was administered intraperitoneally to avoid uncertainties of gastrointestinal absorption. The methanol dosage used for this study was 3 g/kg (one-fourth of the 50% lethality dose) mixed with saline (1:1, vol/vol). Fomepizole was dissolved in ethanol and further diluted in saline (0.09% NaCl wt/vol) to give a final concentration of 1%. Lycopene was dissolved in saline. Methanol and fomepizole (4-methylpyrazole) were purchased from Sigma Chemical Co. (St. Louis, MO, USA). The water-soluble formulation of lycopene (10%) was supplied from DSM Nutritional Products (Istanbul, Turkey).
Tissue preparation
After 24 hours the entire drug dose had been administered, and intoxicated rats were sacrificed under anesthesia (75 mg/kg ketamine hydrochloride and 8 mg/kg xylazine). Their livers were removed quickly and divided into two pieces. One of the pieces was placed in 10% formaldehyde solution for routine histopathological examination by light microscopy. The other piece was placed into liquid nitrogen for MDA analysis and stored at –70°C until assayed.
Determination of MDA level
The tissue MDA level was determined by a method 22 based on the reaction with thiobarbituric acid (TBA) at 90–100°C. In the TBA test reaction, MDA or MDA-like substances and TBA react with the production of a pink pigment with a maximum absorption at 532 nm. The reaction was performed at pH 2–3 at 90°C for 15 minutes. The sample was mixed with 2 volumes of cold 10% (wt/vol) trichloroacetic acid for protein precipitation. The precipitate was pelleted by centrifugation, and an aliquot of the supernatant was reacted with an equal volume of 0.67% (wt/vol) TBA in a boiling water bath for 10 minutes. After cooling, the absorbance was read at 532 nm. The results were expressed as nmol/g wet tissue.
Immunohistochemistry
The detection of caspase-3 activity in paraffin-embedded liver tissues was conducted by using the indirect method of enzyme immunohistochemistry with a rabbit polyclonal antibody (catalog number RB-1197-R7, Thermo Scientific, Fremont, CA, USA) in ready-to-use form. Tissue sections 4 μm thick were deparaffinized, rehydrated, and boiled in 10 mM citrate buffer (pH 6.0) solutions for 10 minutes followed by cooling at room temperature for 20 minutes. H2O2 was applied over tissue sections to block endogenous peroxidase activity, and afterward they were incubated with primary antibody for 30 minutes at room temperature. The rest of the steps were done according to the standard immunohistochemistry staining procedures, and using phosphate-buffered saline solution for washing steps and 3,3′-diaminobenzidine for the chromogen at the last step. Predominantly cytoplasmic fine granular staining was accepted as positive. The caspase activity was evaluated and expressed numerically by counting the number of strongly positive-stained hepatocytes per 10 consecutive high power fields ( × 400) and calculating the mean number.
Histological analysis
For light microscopic evaluation, liver tissues were put in 10% neutral buffered formalin solution and processed by automatic tissue processor for routine tissue processing. The fixed livers were embedded in paraffin blocks and sectioned at 4 μm thickness. Sections were stained with hematoxylin and eosin for light microscopic evaluation (BX-51 light microscope, Olympus, Tokyo, Japan).
The histological activity indices were graded according to the reference evaluation system of Ishak et al. 23 for liver biopsies.
Statistics
All data were expressed as the arithmetic mean ± SEM of the number (n) of experiments; when P < .05, the difference was considered to be statistically significant. Analysis of variance was used to assess the differences between multiple groups for analysis of biochemical data. If statistically significant, the mean values obtained from each group were then compared by LSD multiple comparisons test. The histological results were statistically analyzed by the Kruskal-Wallis H test. The differences between the groups were evaluated by the Mann-Whitney U test.
Results
The levels of liver MDA, as proceeding from lipid peroxidation, are given in Table 1. Although the methanol-alone group had significantly higher liver MDA concentrations compared to the control group, the groups pretreated with lycopene and/or fomepizole had lower MDA levels than the methanol group (P < .0001).
Data are mean ± SEM values.
P < .05 versus control.
MDA, malondialdehyde.
The results of histological evaluation and caspase-3 activity are given in Table 2. In control group the histomorphology was normal (Fig. 1A). No significant lobular lytic necrosis (Fig. 1B) and portal inflammation were observed in the methanol group (Fig. 1C). The results were similar to those of the control group in the lycopene + methanol, fomepizole + methanol, and fomepizole + lycopene + methanol groups (Table 2). The methanol (Fig. 2A) and lycopene (Fig. 2B) groups showed significant increases in caspase-3 activity compared with the control group. In the lycopene + methanol (Fig. 2C), fomepizole + methanol (Fig. 2D), and fomepizole + lycopene + methanol groups (data not shown), caspase-3 activity returned to normal histologic appearances compared with the methanol and lycopene groups.
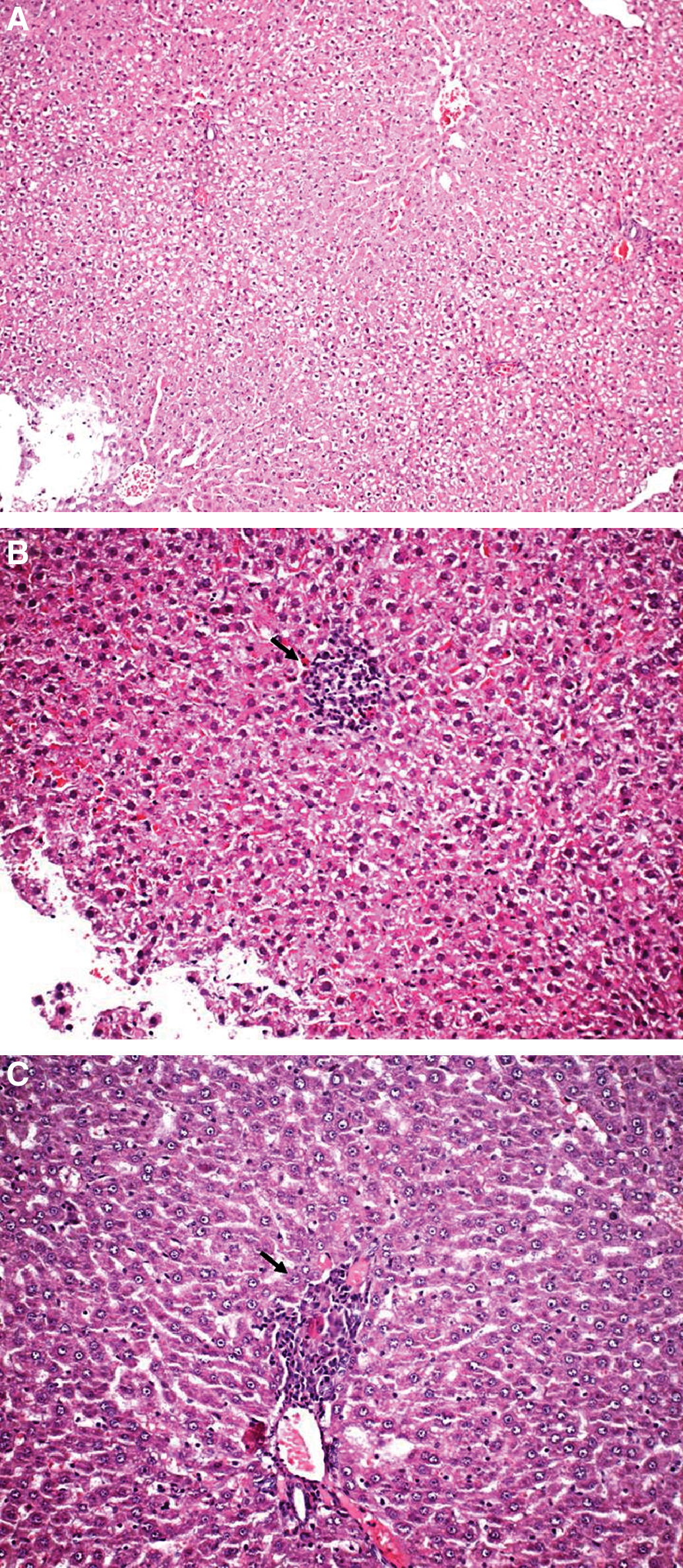
(
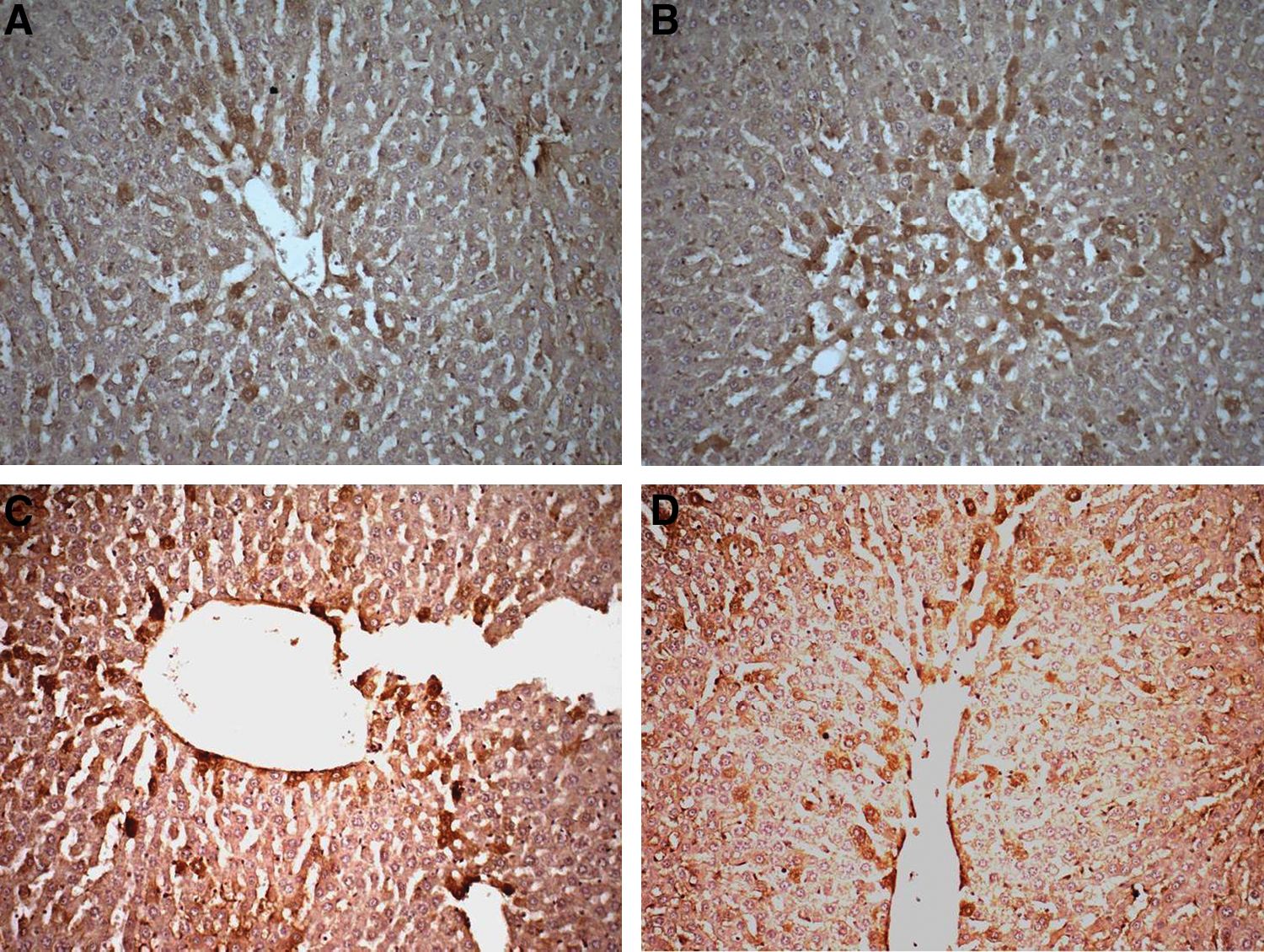
Strong positive cytoplasmic expression of caspase-3 activity in the (
Data are mean ± SEM values.
P < .05 versus control group; † P < .05 versus methanol group.
Discussion
In the present study the increases in MDA level and caspase-3 activity indicate oxidative damage and apoptosis in liver of rats after methanol intoxication. The increases in MDA level and caspase-3 activity were normalized by pretreatment with lycopene and/or fomepizole.
Recent studies reported that lipid peroxidation in liver, erythrocytes, and blood serum and also protein oxidation in liver occurred during methanol intoxication. 24 –26 In our previous study, we observed a significant increase in both lipid peroxidation and protein oxidation of the tissues in methanol-induced hepatic injury. Although the metabolic steps of methanol intoxication are different in humans and rats, the metabolites are the same. 10,27 The main system that functions in methanol intoxication in rodents is the catalase-peroxidase system and alcohol dehydrogenase, which catalyze the conversion of methanol to formaldehyde. As a sequential step, aldehyde dehydrogenase and oxidases, including xanthine oxidase, rapidly metabolize formaldehyde to formate. These processes are accompanied by the formation of superoxide anion and further that of hydrogen peroxide, which are very harmful to liver cells and other cells. 28 These generated free radicals cause extensive damage to cellular membranes leading to cellular dysfunction and death. 29 In this study, the MDA level was high in the methanol group in comparison with the control, lycopene, and fomepizole groups. However, administration of fomepizole and lycopene in combination was not more effective in the decrease of lipid oxidation than using each agent alone.
Apoptosis, a physiological program of cellular death, may contribute to many disorders in a wide range of clinical settings. The mitochondrial-dependent pathway plays an important role in apoptosis by inducing release of cytochrome c and activating caspase-9, which activates caspase-3, which is responsible for cleavage action. 30 Although in previous studies it has been shown that ethanol intoxication induces oxidative stress and apoptosis in liver, brain, and T cells, in this study it is shown for the first time that methanol intoxication also may induce apoptosis via the activation of caspase-3 in liver of rats. The cell death process occurs over a 6–16-hour period following ethanol administration, is accompanied by a robust display of caspase-3 enzyme activation, and meets ultrastructural criteria for apoptosis. 14 Two apoptotic pathways (intrinsic and extrinsic) have been described, either of which may end in the activation of caspase-3. The mitochondrial-dependent apoptotic pathways in rat hearts were activated by long-term intermittent hypoxia. 31 It has been suggested that methanol is responsible for the toxic inhibition of cytochrome c oxidase and that inhibition of the cytochrome c oxidase complex leads to anaerobic glycolysis and lactic acidosis-“histotoxic hypoxia.” 17 We conclude that mitochondrial dysfunction after methanol administration may represent the initiating cellular events that trigger the sequential activation of caspase-dependent signaling cascades, leading to apoptotic cell death in the liver.
Lycopene, an acyclic hydrocarbon carotenoid found in tomatoes and their products, is a well-established potent antioxidant, and its anticancer properties have been shown in cultured cells and animal models.
32
Lycopene, the most effective antioxidant among the carotenoids, is known as a highly efficient scavenger of 1O2 and other excited species. During 1O2 quenching, energy is transferred from 1O2 to the lycopene molecule, converting it to the energy-rich triplet state. Thus, lycopene may protect in vivo against oxidation of lipids, proteins, and DNA.
4,33
In this study, lycopene alone significantly decreased the increased MDA level in liver tissue by methanol, similar to fomepizole, but administration of fomepizole and lycopene in combination was not more effective in decreasing the MDA level. Tomatoes contain many bioactive components, including vitamins C, K, and E, as well as fiber, folate, and polyphenols such as quercetin.
34
Similarly, in this study the lycopene administered contains
Although primarily considered an antioxidant, lycopene appears to have influence on cellular proliferation and differentiation and immune response. 4,5,32,33,35 Lycopene increases apoptosis in several cancer cell lines, including the prostate, 36,37 leukemia, 38 and colon. 39 Lycopene is reported to induce apoptosis and inhibit lung squamous metaplasia and cell proliferation of gastric mucosa in cigarette smoke-exposed ferrets. 40,41 Zhang et al. 38 demonstrated that 4-methyl-8-oxo-2,4,6-nonatrienol, a major oxidation product of lycopene, induces apoptosis in HL-60 cells through down-regulation of Bcl-2, a major anti-apoptotic protein, and up-regulation of caspase-8 and -9. However, Fazekas et al. 32 have suggested that lycopene may act as a preventative agent via reducing inflammatory responses, maintaining normal cell proliferation, and possibly preventing DNA damage as indicated by blocking the necessitating step of apoptosis following ultraviolet B-induced photodamage. Similarly, in this study it has been shown that lycopene pretreatment prevents caspase-3 activation in liver of methanol-administered rats. However, in our study, only lycopene administration induces caspase-3 activation in liver of rats. Velmurugan et al. 42 reported that the activity of caspase-3 and −8 in gastric mucosa of lycopene alone-administered rats was not significantly different from that in controls. Wang et al. 43 suggested that dietary phytochemicals such as lycopene may be altered the apoptotic threshold. In vitro, lycopene has been shown to inhibit the growth of human androgen-independent PC cells and rat Dunning AT6.3 cells, as well as in vivo by increasing the apoptotic rate in the human prostate cells. 43 –45 Tomato polyphenols also enhanced the sensitivity to apoptosis in the Dunning AT6.3 cell line. 43 Interestingly, Canene-Adams et al. 34 reported that in the Dunning R3327-H prostate cancer model, neither lycopene-fed group showed changes in apoptotic rates in tumors, yet tomato consumption resulted in increased apoptosis. These differences among the studies' results may be related to experimental models such as dose and administration route of lycopene, animal species used, tissue evaluated, or disease of animals.
In our study, histopathologic damage scores were found to be higher in methanol-induced intoxicated rats compared with the controls, but not significantly. Controversially, in our previous study, we observed significant methanol-induced histopathologic damage, such as piecemeal necrosis, lobular lytic necrosis, and portal inflammation. 20 This condition may be related to the administration route of methanol. In our previous study, methanol was administered via the intragastric route. We consider that oral administration of methanol produced much more toxicity in the liver than the intraperitoneal route.
In conclusion, the findings of the present study demonstrate that methanol intoxication causes hepatic toxicity in rats and that this is likely a result of reactive oxygen species and apoptosis induction. It was demonstrated for the first time that both lycopene and fomepizole prevent methanol-induced hepatic injury by reducing the increase in lipid oxidation and caspase-3 activation. Lycopene has protective effects against methanol-induced hepatic injury similar to fomepizole. However, administration of fomepizole and lycopene in combination was not more effective in decreasing lipid oxidation and caspase-3 activity than each agent alone.
Footnotes
Author Disclosure Statement
No competing financial interests exist.