
Abstract
Dietary flavonoids have various biological functions, and there is increasing evidence that reduced prevalence and severity of allergic reactions are associated with the intake of flavonoids. Among natural flavonoids, apigenin is a potent anti-inflammatory agent. However, the mechanisms of apigenin's effect remain uncertain. Monocyte-derived chemokine (MDC) plays a pivotal role in recruiting T-helper (Th) 2 cells in the allergic inflammation process. In the late phase of allergic inflammation, the Th1 chemokine interferon-inducible protein 10 (IP-10) has also been found in elevated levels in the bronchial alveolar fluid of asthmatic children. We used human THP-1 monocyte cells, pretreated with or without apigenin, prior to lipopolysaccharide stimulation. By means of enzyme-linked immunosorbent assay, we found that apigenin inhibited production of both MDC and IP-10 by THP-1 cells and that the suppressive effect of apigenin was not reversed by the estrogen receptor antagonist ICI182780. The p65 phosphorylation of nuclear factor κB remained unaffected, but the phosphorylation of p38, c-Jun N-terminal kinase, and extracellular signal-regulated kinase mitogen-activated protein kinase pathways were all blocked. We found that inhibition of c-raf phosphorylation might be the target of apigenin's anti-inflammation property.
Introduction
T
The interaction between genetic and environmental factors is generally accepted as inducing allergic diseases. 13,14 Changing of dietary habits is thought to be one of the most important environmental factors contributing the increasing prevalence of allergic diseases. 15,16 Many nutrient components could be allergens, but may also be anti-allergic remedies. Recent evidence shows that flavonoids, ubiquitously present in vegetables, fruits, and teas, have anti-allergic activities. 17 Natural flavonoids are classified into eight groups, according to their skeletons, with more than 8,000 different flavonoids having been identified. 18 Several flavonoids, such as luteolin, fisetin, and apigenin, inhibit IL-4 and IL-13 synthesis by allergen- or anti-immunoglobulin E antibody-stimulated basophils. 19 The study by Woo et al. 20 showed that apigenin modulates prostaglandin and nitric oxide production by regulation of nuclear factor κB (NF-κB). Recently, apigenin was found to inhibit LPS-induced IL-1β, tumor necrosis factor, and IL-8 expression in human monocytes by inactivating NF-κB through the suppression of p65 phosphorylation. 21 However, the effects and mechanisms of apigenin on chemokine expression are still poorly understood.
Flavonoids have attracted a great deal of attention recently because of their phytoestrogenic effects and associated anticancer activity. They can stimulate transcriptional activity of both estrogen receptor (ER) α and β subtypes and may activate a physiological estrogenic effect. 22 Because of the observation that chronic inflammatory diseases, such as rheumatoid arthritis or inflammatory bowel syndrome, are ameliorated during pregnancy, 23 estrogen has been thought to offer a protective effect from inflammatory processes. Furthermore, the prevalence of coronary heart diseases decreases in postmenopausal females undergoing estrogen replacement therapy, which further confirms the role of estrogen in the prevention of cardiovascular inflammation and the pathogenesis of artherosclerosis. 24 Furthermore, 17β-estradiol normalizes inflammatory responses of epidermal keratinocytes following trauma-hemorrhage events through inhibition of mitogen-activated protein kinase (MAPK) pathways. 25 Recent studies have also shown that 17β-estradiol can regulate different components of NF-κB pathways. 26 However, there have been no reports discussing the correlation between phytoestrogens and their anti-inflammatory effects. This study might be the first to report the effect of apigenin on the Th1- and Th2-related chemokines MDC and IP-10 in THP-1 cells, which might be regulated through c-raf-mediated MAPK pathways. However, the ER and NF-κB pathways may not be involved in the anti-inflammatory effect of apigenin.
Materials and Methods
Cell preparation
The human monocytic cell line THP-1 (American Type Culture Collection, Rockville, MD, USA) was cultured in RPMI 1640 medium (Sigma Chemical Co., St. Louis, MO, USA) supplemented with 10% fetal bovine serum, 100 U/mL penicillin, and 100 μg/mL streptomycin at 37°C in a humidified incubator with a 5% CO2 atmosphere. Cells were centrifuged and resuspended in fresh medium in 24-well plates at a concentration of 5 × 105/mL for 24 hours before experimental use. Cells were pretreated with and without apigenin (10−7–10−5 M) (Sigma) or 17β-estradiol (10−8–10−6 M) (Sigma) 2 hours before LPS (0.2 μg/mL) (Escherichia coli) (Sigma) stimulation. The ER antagonist ICI182780 (10−6 M) (Sigma) was added 1 hour before apigenin treatment to block ERs. Free cell supernatant was collected after different incubation times.
Cell viability
Apigenin was dissolved in a few drops of dimethyl sulfoxide and diluted with phosphate-buffered saline to give final concentrations. Various concentrations of apigenin were incubated in a 96-well plate for 24–48 hours. The XTT cell proliferation assay (Biological Industries Ltd., Kibbutz Beit Haemek, Israel) was used to determine the cytotoxicity. XTT activation solution was mixed with reagent at a 1:50 ratio. Then 50 μL of reaction solution was added to each well, and the plate was incubated for 5 hours. The cell viability was calculated by measuring the absorbance of the sample with an enzyme-linked immunosorbent assay reader, using a wavelength of 540–630 nm. The mean value of the content of four wells was used to assess the cell viability expressed as a percentage of the control value.
Assaying levels of MDC and IP-10
MDC and IP-10 were measured by enzyme-linked immunosorbent assay kits (R&D Systems Inc., Minneapolis, MN, USA). Each experiment in which MDC and IP-10 levels were measured was performed at least three times. Samples were read in a Dynatech MRX plate reader at 450 and 540 nm using Revelation software (Dynatech Laboratories, Alexandria, VA, USA).
Cytosolic and nuclear protein extraction
THP-1 cells (1 × 106) were pretreated with apigenin (10−7–10−5 M) for 2 hours, then stimulated without or with LPS (0.2 μg/mL) for 1 hour, washed with iced phosphate-buffered saline once, and then resuspended. The cells were lysed in 10 mM HEPES (pH 7.9), 1.5 mM MgCl2, 10 mM KCl, 300 mM sucrose, 0.5% Nonidet P-40, and proteinase inhibitor cocktail (1.0 mM phenylmethylsulfonyl fluoride, 1.0 mM EDTA, 1 μM pepstatin A, 1 μM leupeptin, and 0.1 μM aprotinin) for 3 minutes on ice and then centrifuged at 3,800 g for 20 seconds. The supernatants were collected for cytosolic protein analysis. The precipitants were resuspended using 20 mM HEPES (pH 7.9), 1.5 mM MgCl2, 420 mM NaCl, 1 mM dithiothreitol, 0.2 mM EDTA, 25% glycerol, and proteinase inhibitor cocktail on ice for 30 minutes and then centrifuged at 13,200 g for 5 minutes. The supernatants (nuclear lysate) were diluted with an equal amount of 20 mM HEPES (pH 7.9), 10 mM KCl, 1 mM dithiothreitol, 0.2 mM EDTA, 25% glycerol, and proteinase inhibitor cocktail.
Analysis for immunoblot
Cells were treated with apigenin as described above, and equal amounts of whole-cell lysates were analyzed by western blotting with various antibodies, including anti-p38, anti-phospho-p38, anti-c-Jun N-terminal kinase (JNK), anti-phospho-JNK, anti-extracellular signal-regulated kinase (ERK), anti-phospho-ERK, anti-MAPK/ERK kinase kinase (MEK [synonym MAPKK]) 1/2, anti-phospho-MEK1/2, anti-MEK4, anti-phospho-MEK4, anti-c-raf, and anti-phospho-c-raf antibodies (Santa Cruz Biotechnology, Santa Cruz, CA, USA). In some cases, cytosolic and nuclear extracts were analyzed with anti-p65 and anti-phospho-Ser536-p65 antibodies (Cell Signaling Technology, Danvers, MA, USA). Immunoreactive bands were visualized and detected by the Molecular Imager ChemiDoc XRS system (Bio-Rad Laboratories, Richmond, CA, USA).
Assaying MAPK activity
Cell lysates were obtained as previously described. Each 200 μL of cell lysate had 20 μL of the immobilized antibody added (total and phosphorylated Ets-like gene 1 [Elk-1] and activating transcription factor [ATF]-2 obtained from Cell Signaling) and was incubated with gentle rocking overnight at 4°C. Samples were microcentrifuged for 30 seconds at 4°C. The pellet was washed five times with 500 μL of 1 × cell lysis buffer. After washing, the pellet was resuspended with 20 μL of 3 × sodium dodecyl sulfate sample buffer. It was then vortex-mixed and microcentrifuged for 30 seconds. The sample was heated to 95–100°C for 2–5 minutes and then loaded (15–30 μL) onto sodium dodecyl sulfate-polyacrylamide gel electrophoresis gel (12–15%). The sample was analyzed by the immunoblot protocol described above.
Statistical analyses
All data are presented as mean ± SD values. One-way analysis of variance was used for all statistical comparisons, and the Student-Newman-Keuls test was used for multiple comparisons. A value of P < .05 was considered indicative of significant between-group differences. SPSS version 12.0 for Windows (LEAD Technologies, Chicago, IL, USA) was used for all statistical analyses.
Results
Apigenin had no effect on proliferation of THP-1 cells
We determined the effect of apigenin on the cell proliferation of THP-1 cells by XTT assay. Our results revealed that apigenin was not cytotoxic to THP-1 cells at the concentration we used (10−5 M) after 24 and 48 hours of treatment (Fig. 1).
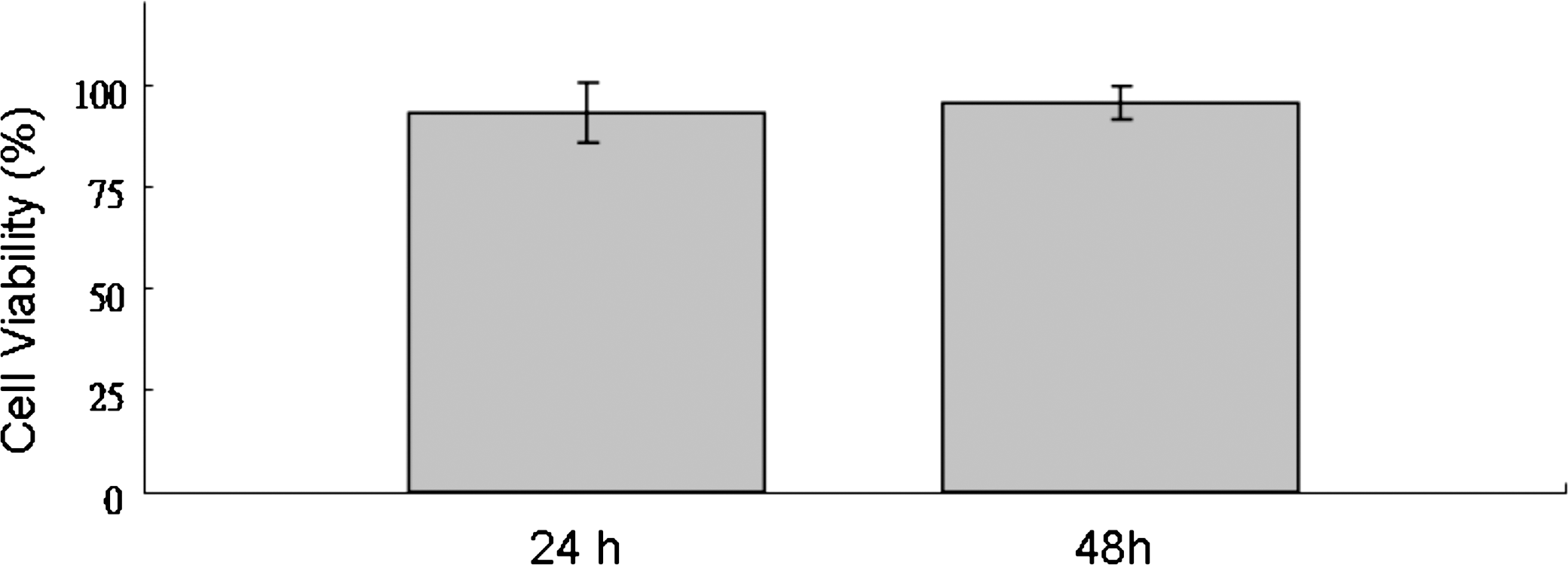
Effect of apigenin on cell viability of THP-1 cells. THP-1 cells were cultured with 10−5 M apigenin in a 96-well plate for 24–48 hours. Cell viability was determined by XTT assay and expressed as a percentage of the control (without apigenin). Data are mean ± SD values of six individual experiments.
Apigenin inhibits release of the inflammatory chemokines MDC and IP-10 in LPS-stimulated THP-1 cells
Apigenin has been found to inhibit inflammatory cytokines in LPS-stimulated human monocytes in previous studies. 21 To investigate the effect of apigenin on chemokine release, THP-1 cells were treated with LPS (0.2 μg/mL) alone or with LPS and different concentrations of apigenin 2 hours before or left untreated for different periods (12 and 48 hours). Stimulation with LPS resulted in increased expression of the Th1- and Th2-related chemokines MDC and IP-10, respectively (Figs. 2 and 3). Next we examined the effect of apigenin on LPS-stimulated THP-1 cells. The results showed significant suppression of MDC expression at apigenin concentrations of 10−6 M and higher (Fig. 2) at different times. Apigenin at concentrations of 10−5 M or higher significantly inhibited the expression of IP-10 by LPS-stimulated THP-1 cells (Fig. 3) at different times. These results indicate that apigenin inhibits the release of the Th1- and Th2-related inflammatory chemokines in THP-1 cells stimulated by LPS.
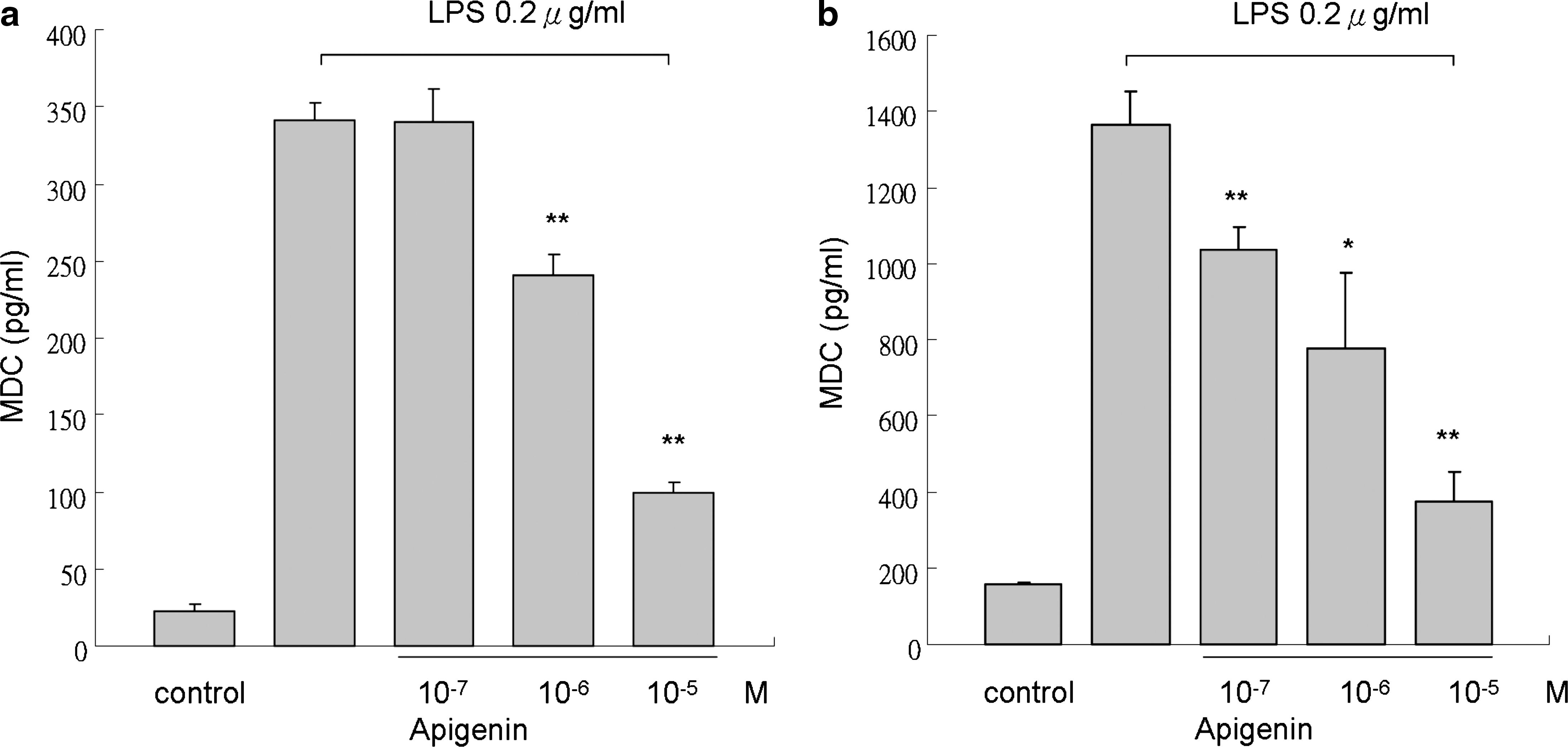
Effect of apigenin on release of MDC by THP-1 cells. MDC was quantified in supernatant of THP-1 cells cultured for (
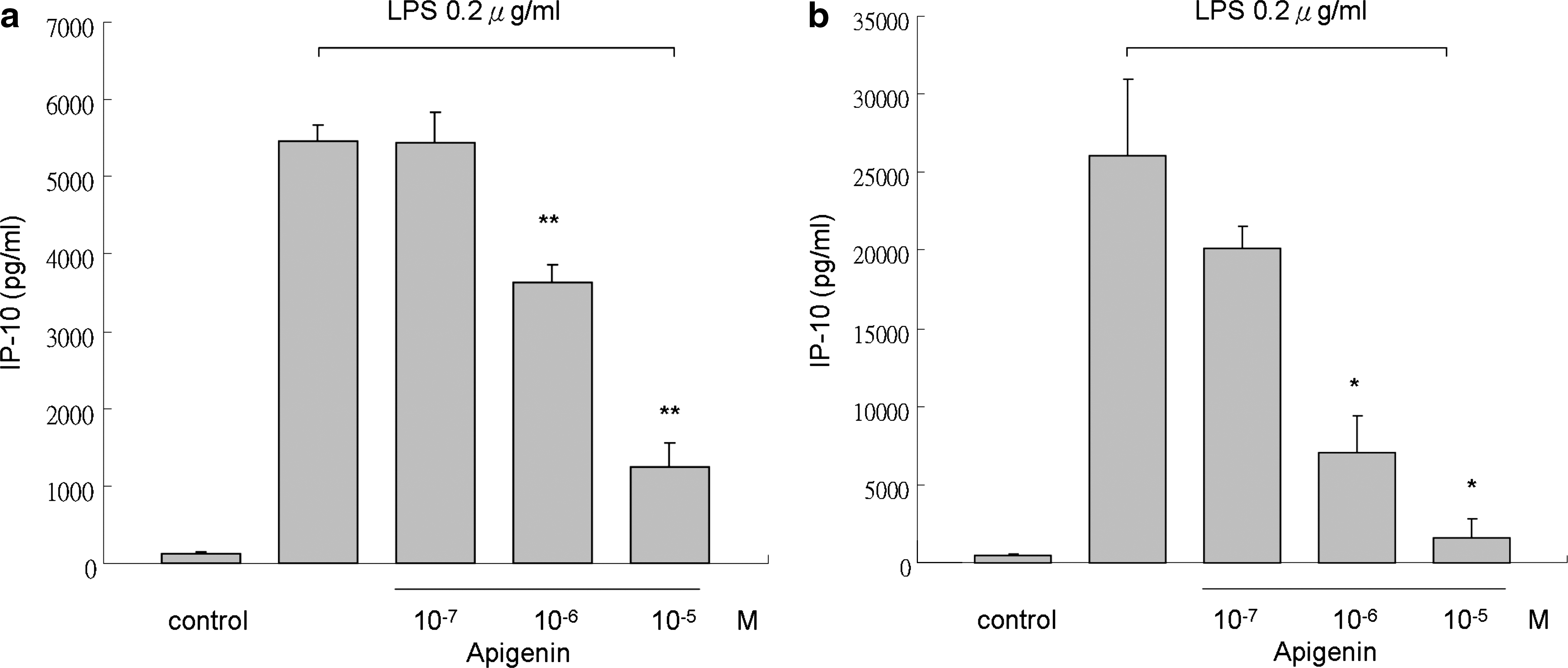
Effect of apigenin on release of IP-10 by THP-1 cells. IP-10 was quantified in supernatant of THP-1 cells cultured for (
17β-Estradiol inhibit MDC and IP-10 production in LPS-stimulated THP-1 cells
17β-Estradiol had been found to mediate the inflammatory process through various mechanisms and in different models. For example, estradiol blocks the induction of CD40 and CD40L expression in endothelial cells and prevents further neutrophil adhesion through an ERα-mediated pathway. 27 Estradiol has also been shown to increase endothelial cell nitrite oxide synthase secretion in rat blood vessels. 28 Consequently, IL-1β-induced cyclooxygenase 2 expression in rat cerebral blood vessels is suppressed by 17β-estradiol treatment. 29 Estrogen also manifested effects on cytokines and chemokines in an experimental autoimmune encephalomyelitis model. 30 We examined the effect of estrogen on chemokine expression in LPS-stimulated THP-1 cells because apigenin is capable of activating ERs. We found that 17β-estradiol, like apigenin, significantly suppresses MDC and IP-10 expression in LPS-stimulated THP-1 cells at 24 hours (Fig. 4).
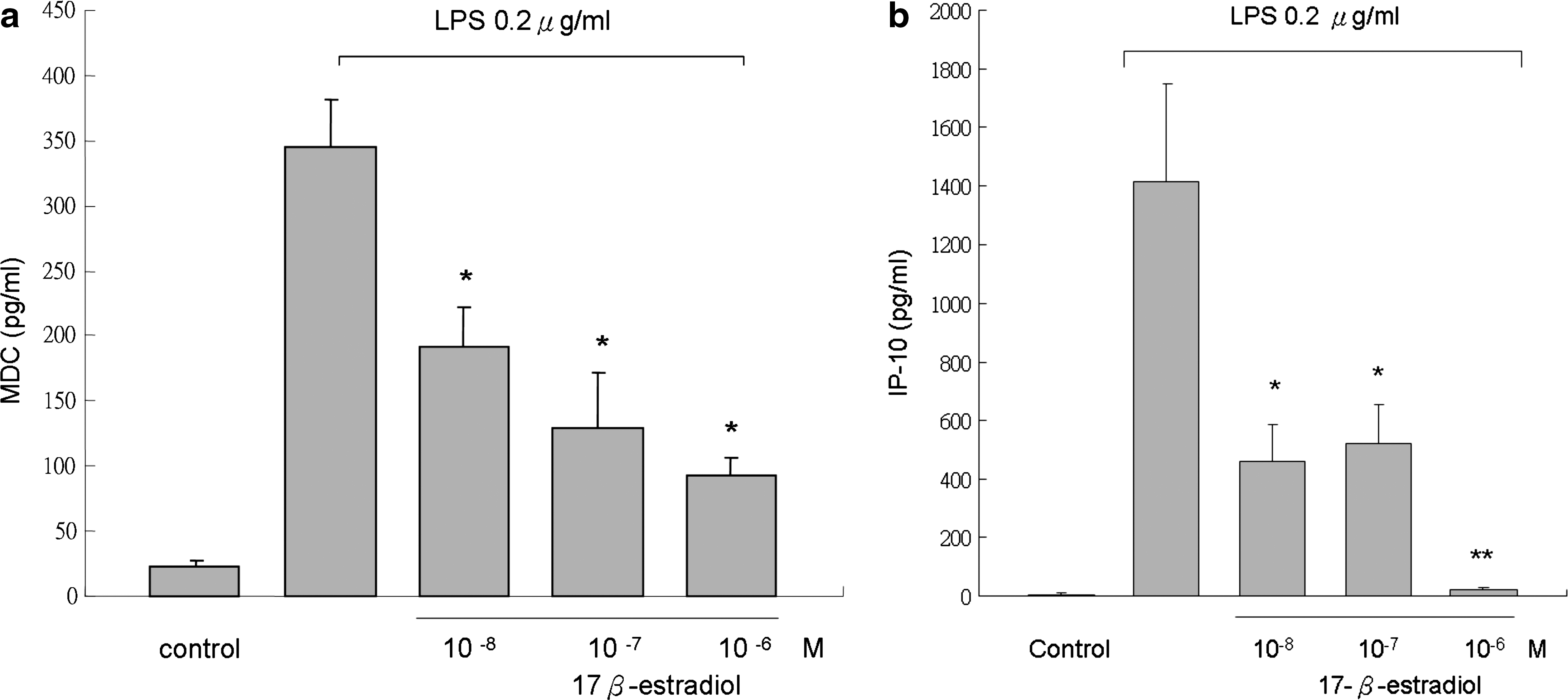
Effects of 17β-estradiol on (
ER antagonist ICI182780 could not reverse the suppressive effect of apigenin in LPS-stimulated THP-1 cells
According to the above data, we hypothesized that apigenin might suppress expression of inflammatory chemokine MDC and IP-10 through an ER-mediated pathway. We therefore added the ER antagonist ICI182780 1 hour before apigenin treatment and then stimulated THP-1 cells with LPS 1 hour after apigenin treatment. Cell supernatant was collected 24 hours later for chemokine analysis. The results showed that MDC and IP-10 expression was not reversed by ICI182780, which suggests that the effect of apigenin might not be mediated through ERs (Fig. 5).
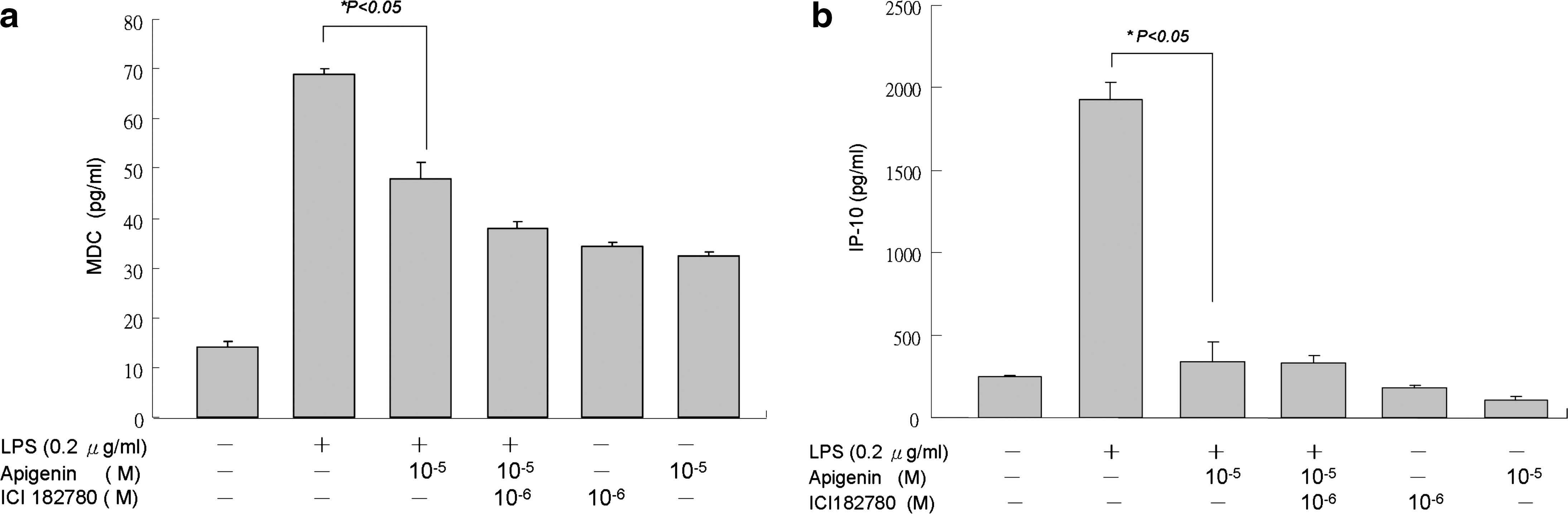
The inhibitions of (
Apigenin suppresses chemokine production through inhibition of JNK, ERK, and p38 MAPK pathways
Ligation of Toll-like receptor 4 with LPS activates downstream signaling pathways, such as MAPK and NF-κB, initiating production of pro-inflammatory cytokines and chemokines. We next determined the effect of apigenin on MAPK signaling pathways. As shown in Figure 6, apigenin reduced the expression of phosphorylated p38, JNK, and ERK MAPK expression after 60 minutes of stimulation. Meanwhile, the phosphorylation of MAPKKs, including MEK1/2 and MEK4, was also suppressed, but MEK3/6 was not detected (data not shown). The phosphorylation of MAPKK kinase (also referred to as MEKK or MAP3K) c-raf was also suppressed (Fig. 6d). Additionally, the MAPK activities were blocked (Fig. 7), which indicates that apigenin might inhibit MDC and IP-10 production through c-raf-mediated MAPK pathways.
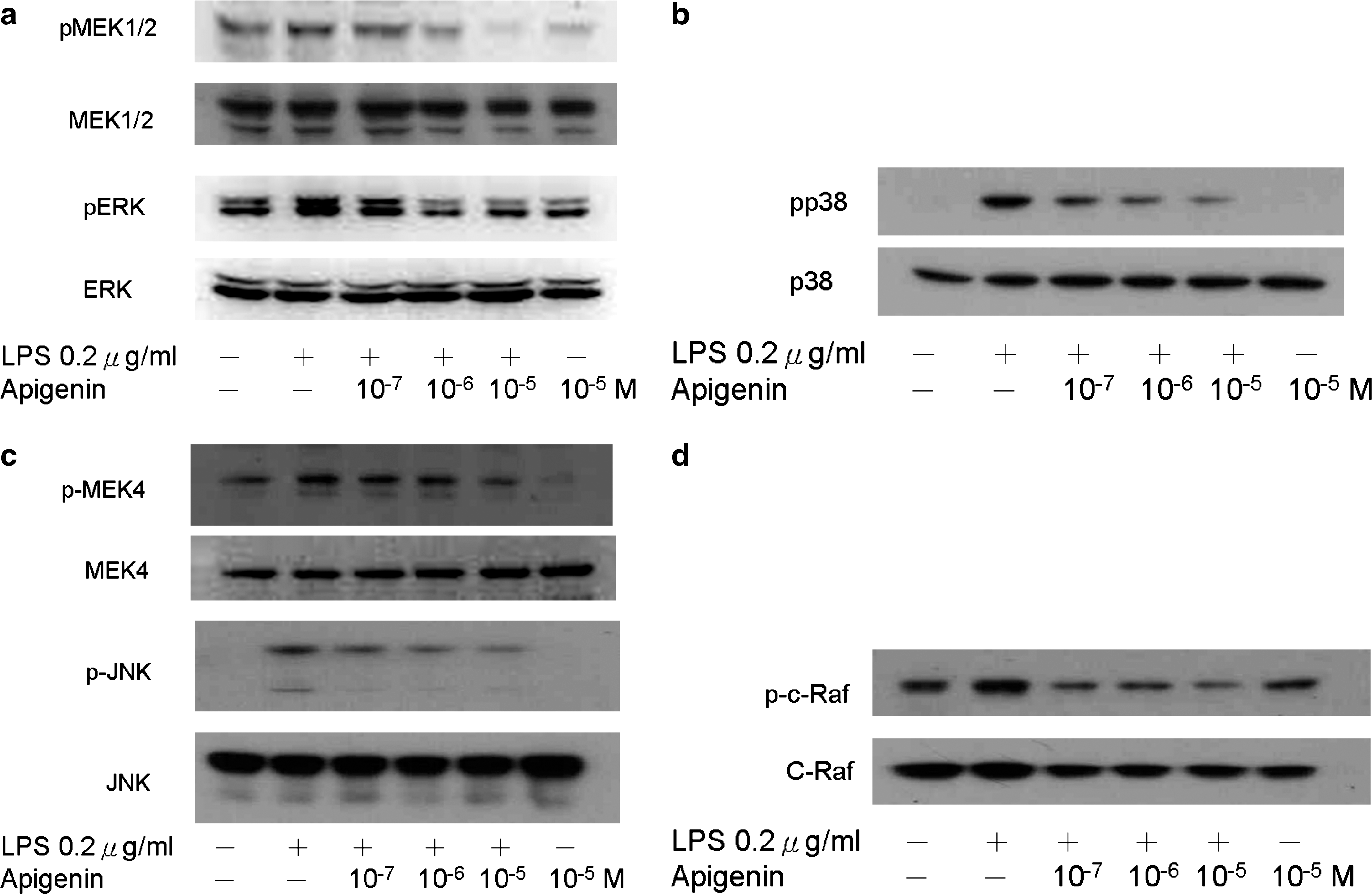
Role of MAPK pathways on the suppression effect of apigenin on chemokine production in THP-1 cells. THP-1 cells were incubated with different concentrations of apigenin (10−7–10−5 μM) for 2 hours and stimulated with LPS (0.2 μg/mL). Cell extracts were collected and treated with primary antibodies to total and phosphorylated p38, ERK, and JNK. The experiments shown are representative of three independent experiments. (
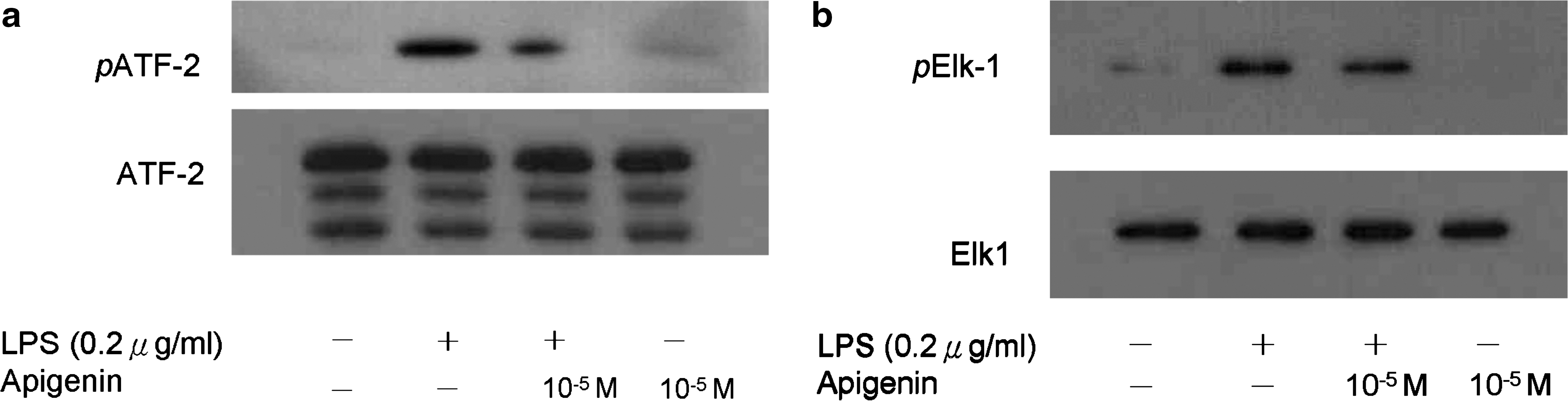
MAPK activity of ATF-2 and Elk-1. Cells were pretreated with 10−5 M apigenin 2 hours before LPS (0.2 μg/mL) stimulation. The phosphorylation of both (
Apigenin did not suppress phosphorylation of the p65 component of the NF-κB pathway
In previous studies, apigenin modulated LPS-induced phosphorylation of the NF-κB p65 subunit at Ser536 independently of the LPS-triggered degradation of IκBα. 21 We therefore examined the effect of apigenin on transcription activity of NF-κB. There was no translocation of pp65 in the nucleus (data not shown), and no significant changes of Ser536 phosphorylation in the cytosol was noted (Fig. 8). The results suggest that in the THP-1 cell model, NF-κB might not be the major pathway for inhibition of chemokine expression by apigenin.
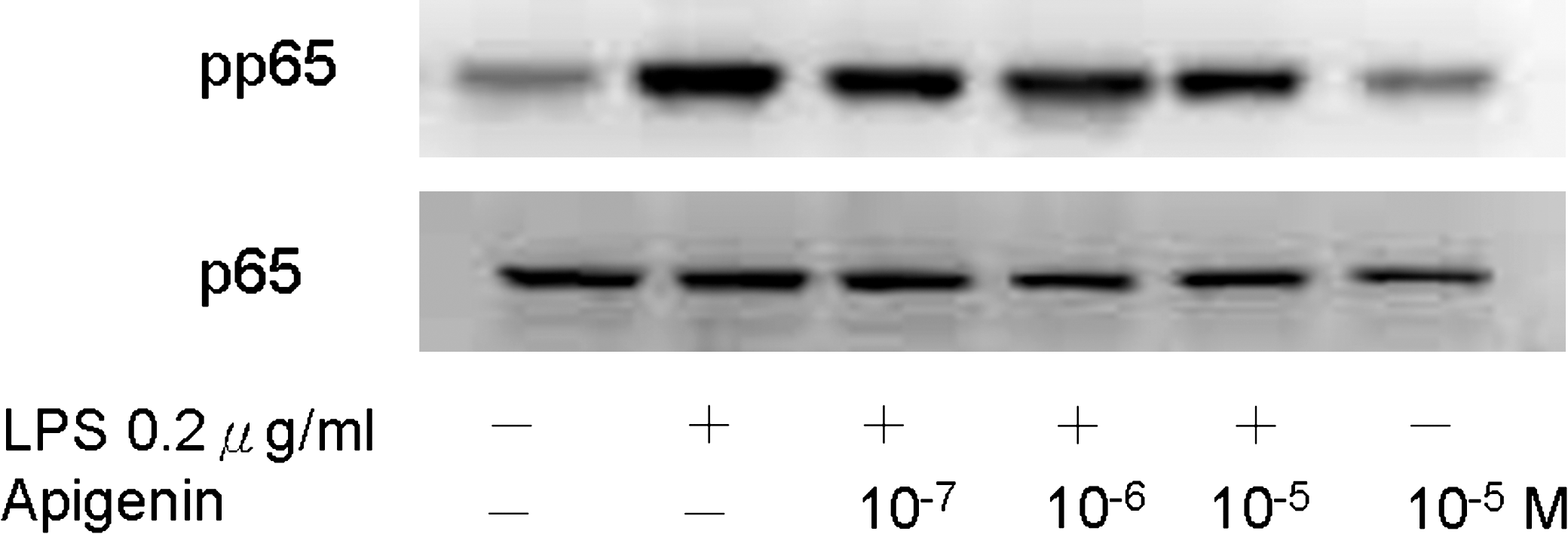
Role of the NF-κB pathway on the effect of apigenin in THP-1 cells. Cells were pretreated with different concentrations of apigenin (10−7–10−5 M) 2 hours before LPS (0.2 μg/mL) stimulation. The phospho-Ser536-p65 was not significantly affected by apigenin. The experiments shown are representative of three independent experiments.
Discussion
Immune responses are initiated by invasion of microbial pathogens. Homeostasis is decided by balancing the Th1 and Th2 responses after engagement of foreign antigens. Early exposure to microbial components, such as airborne LPS, leads to increased Th1 immune response and tolerance to allergens. 10,31 LPS can be sensed by monocytes and macrophages via Toll-like receptor 4, which in turn triggers signaling events that produce inflammatory cytokines and chemokines. 32,33 These pathways, including NF-κB and MAPK pathways, mediate responses of various inflammatory genes. Although the pathways regulating these mechanisms are still not fully understood, Toll-like receptors and the downstream signaling are important for innate recognition and subsequent connection to adaptive immunity. 34
The potential anti-inflammatory effects of plant flavonoids have been confirmed in both in vivo and in vitro experiments. 35 Despite the lack of large-scale epidemiological studies, dietary flavonoids have been found to be of some benefit in dealing with allergies. 17 Additionally, although exposure to airborne endotoxins in early infancy may provide protection from developing asthma, later in life they may aggravate asthmatic symptoms by increasing the severity of airway inflammation. 10,31 Therefore, environmental factors can be seen to play vital roles in allergic disease pathogenesis.
Apigenin and other flavonoids have been widely studied recently, especially for their anti-inflammatory properties. 35 Suppression of inflammatory cytokines through modulation of the NF-κB pathway has been noted in previous studies. 36 Because chemokines are important in the pathogenesis of allergic diseases and correlated with disease severity, we examined the effect of apigenin on chemokine expression. Our findings showed that apigenin inhibits MDC and IP-10 production in LPS-stimulated THP-1 cells in a dose-dependent manner and that the effect may persist at least 48 hours (Figs. 2 and 3).
According to the literature, flavonoids also possess phytoestrogenic properties, although differences exist in the ER binding affinity between various compounds. However, there are few reports about the flavonoids discussing the correlations between phytoestrogenic and anti-inflammatory properties, and there are also few studies about apigenin and its effects on chemokine expression. We used 17β-estradiol to simulate the phytoestrogenic property of apigenin and surprisingly found that 17β-estradiol also inhibited MDC and IP-10 production (Fig. 4). However, the ER antagonist ICI182780 was unable to significantly reverse the effect (Fig. 5). Therefore, the effect of apigenin on Th1- and Th2-related chemokine expression might not be mediated via ligation of ER.
As discussed in our previous studies, the MAPK signaling pathway is important for MDC and IP-10 production by LPS-stimulated THP-1 cells, especially the p38 and JNK-MAPK pathways. 9 Apigenin was also able to suppress IL-1β, IL-8, and tumor necrosis factor production by LPS-stimulated monocytes via modulation of the transcriptional activity of NF-κB. 21 Our findings showed that apigenin inhibited phosphorylation of JNK, ERK, and p38 MAPK (Fig. 6). The phosphorylation of upstream MAPKKs, MEK1/2 and MEK4, was also suppressed. The phosphorylation of MAP3K c-raf was suppressed, and the MAPK activity analysis revealed decreased phospho-ATF-2 and Elk-1 (Fig. 7). The MAPK cascades do not always proceed in a linear direction, but in more complex cross-reactive pathways. For example, c-raf contributes directly to ERK activation but not to JNK activation, whereas MEKK participated in JNK activation but caused ERK activation only after overexpression. 37 Our data indicate that apigenin might inhibit MDC and IP-10 expression through modulating c-raf-mediated MAPK pathways. However, the phosphorylation of the Ser536-p65 component of NF-κB was not evident (Fig. 8).
In summary, our study provides important clues to the mechanism of apigenin's anti-inflammatory activity. That is, apigenin may modulate inflammatory chemokine expression through MAPK pathways rather than the NF-κB pathway. Furthermore, the anti-inflammatory activity of apigenin might not be regulated through an ER-mediated pathway. Further animal model experiments and human trials should be carried out to verify the clinical benefits of apigenin on asthma and other allergic diseases.
Footnotes
Acknowledgments
The authors thank the Division of Research Resource, Department of Medical Research, Kaohsiung Medical University Hospital for providing experiment space and experimental instruments. This study was supported by research grant KMUH-96-6G23 from the Kaohsiung Medical University Hospital.
Author Disclosure Statement
No competing financial interests exist.