
Abstract
This study was designed to evaluate the effect of dietary conjugated linoleic acid (CLA) on endurance exercise and availability of metabolic substrates in mice. Four-week-old male BALB/c mice were randomized to a control group (normal diet) and a CLA group (normal diet + 1% CLA). Each animal group received its respective diet for 10 weeks and was subjected to forced running on a treadmill system to estimate their maximum endurance capacity at the end of the experiment. All mice were immediately sacrificed after prolonged exercise, and the major metabolic substrates were measured in serum and liver. The body weights of mice in the CLA group were lower than those of the control group after the 10 weeks. The maximum running time in CLA-fed mice was significantly longer, by 26%, compared to that of the control mice. Dietary CLA decreased the serum concentrations of triglycerides, nonesterified fatty acids, and urea nitrogen and significantly reduced the consumption of liver glycogen. The present study demonstrated that dietary CLA enhances the endurance capacity of mice by increasing fat utilization and reducing the consumption of stored liver glycogen as substrates for energy metabolism.
Introduction
C
Park and Pariza 6 described the mechanisms of body fat reduction by CLA, which included four factors: increasing energy expenditure, reducing adipose cell mass and/or cell numbers, modulating adipokines and cytokines, and increasing fatty acid β-oxidation in skeletal muscle. In particular, increasing fatty acid β-oxidation by CLA was suggested by enhanced activity and/or expression of carnitine palmitoyl transferase I, the rate-limiting enzyme for fatty acid β-oxidation, in skeletal muscle. 7,8 In addition, increased muscle lipoprotein lipase activity by CLA may play a role in enhanced fatty acid oxidation in skeletal muscle. 9 These results indicate that CLA causes fat to be preferentially used as an energy source in skeletal muscle.
Fatty acid oxidation in muscular mitochondria followed by aerobic reactions of the tricarboxylic acid cycle is adequate to generate a large proportion of the ATP required for muscular contraction during prolonged exercise. 10 Thus, enhanced fat oxidation could decrease carbohydrate consumption by the muscle during exercise and increase endurance capacity. An enhanced exercise capacity could be accounted for by the reduced rate of hepatic and muscular glycogen breakdown and by a greater potential for fatty acid metabolism. 11 Mizunoya et al. 9 reported that the maximum swimming time of mice was increased with enhanced fat oxidation after supplementing their diets with CLA for 1 week. The purpose of this study was to evaluate the effects of dietary CLA on endurance capacity and the consumption of metabolic substrates in mice during treadmill exercise after feeding for 10 weeks.
Materials and Methods
Animals and diet
Male BALB/c 4-week-old mice (Orient Bio Inc., Seongnam-Si, Republic of Korea) were used and housed in standard cages (33 × 23 × 12 cm, five mice per cage) under controlled conditions of temperature (22 ± 0.5°C), humidity (50%), and lighting (light from 9 a.m. to 9 p.m.). After a 1-week adaptation period, the mice were assigned to two groups that were fed ad libitum with a standard diet (control group) or a diet containing 1% CLA in triacylglyceride form (CLA group). The composition of each diet, stored frozen until used, is presented in Table 1. The CLA was provided by HK Biotech (Jinju, Republic of Korea). Each animal group received its respective diet for 10 weeks. Body weight and food intake were recorded every week throughout the study. All mice were acclimatized to the treadmill five times during the last week of the experiment with 7 minutes of running at 17 m/minute and 0° inclination. At the end of the experimental period, mice were subjected to forced running to estimate their maximum endurance capacity. They were then immediately anesthetized with ethyl ether, and blood was collected from the vena cava. The blood samples were centrifuged to separate the serum and stored at −20°C until further assays. The livers were dissected, immediately frozen in liquid nitrogen, and stored at −80°C until analysis. The care and treatment of experimental animals conformed to a protocol approved by the Institutional Animal Care and Use Committee of Korea University (Seoul, Republic of Korea).
Mineral mixture and vitamin mixture were based on the AIN-76A formulation. 12
Conjugated linoleic acid (CLA) contained 5.75% C16:0, 2.28% C18:0, 14.50% C18:1, 0.78% C18:2, 37.33% c9,t11-CLA isomer, 37.13% t10,c12-CLA isomer, 0.93% c9,c11/c10,c12-CLA isomer, and 1.31% t9,t11/t10,t12-CLA isomer.
Evaluation of the maximum running capacity of mice
Mice were exercised on a motorized treadmill (Mirae-ST Corp., Daejeon, Republic of Korea) that had an adjustable belt speed (0–50 m/minute) and adjustable inclination (0–20°). To evaluate the endurance capacity, the mice were placed on individual treadmill lanes at room temperature. The exercise regimen was started at a velocity of 21 m/minute on a slope of 10°. The speed was then increased by 2 m/minute from 18 minutes until it was up to 29 m/minute. When mice slid into the electric shock grid (delivered 1.2 mA of current at 60 Hz) located at the back of the treadmill, they were electrically shocked; this served to encourage them to move to the front of the treadmill. The total running time until exhaustion was recorded, and it was used as the index of endurance capacity. Mice were assessed as fatigued when they failed to run and remained for more than 3 seconds on the shock grid. At the moment of exhaustion the mouse was removed from the treadmill.
Estimation of biochemical parameters
The serum levels of glucose, triglycerides, and urea nitrogen were measured with a PRIME automatic photometer (BPC Biosed, Rome, Italy) using commercial kits (Asan Pharmaceutical, Seoul). Serum nonesterified fatty acid (NEFA) was measured using a commercial kit (NEFA Test, Wako Pure Chemical Industries, Richmond, VA, USA).
The tissue glycogen content was measured as glucose residues after hydrolysis of the muscle sample in 1 mol/L HCl at 100°C for 2 hours using the method of Hassid and Abraham. 13
Statistical analyses
Data are expressed as mean ± SEM values for eight to nine animals. Statistical differences were determined using the two independent sample t test in the SAS statistical software package (SAS Institute Inc., Cary, NC, USA); P < .05 was accepted as a significant difference.
Results
No significant difference in food intake was evident between the control group (3.3 g per mouse per day) and the CLA group (3.7 g per mouse per day) during the experimental period. The body weights of mice in the CLA group were lower than those of the control group after 10 weeks (Fig. 1).
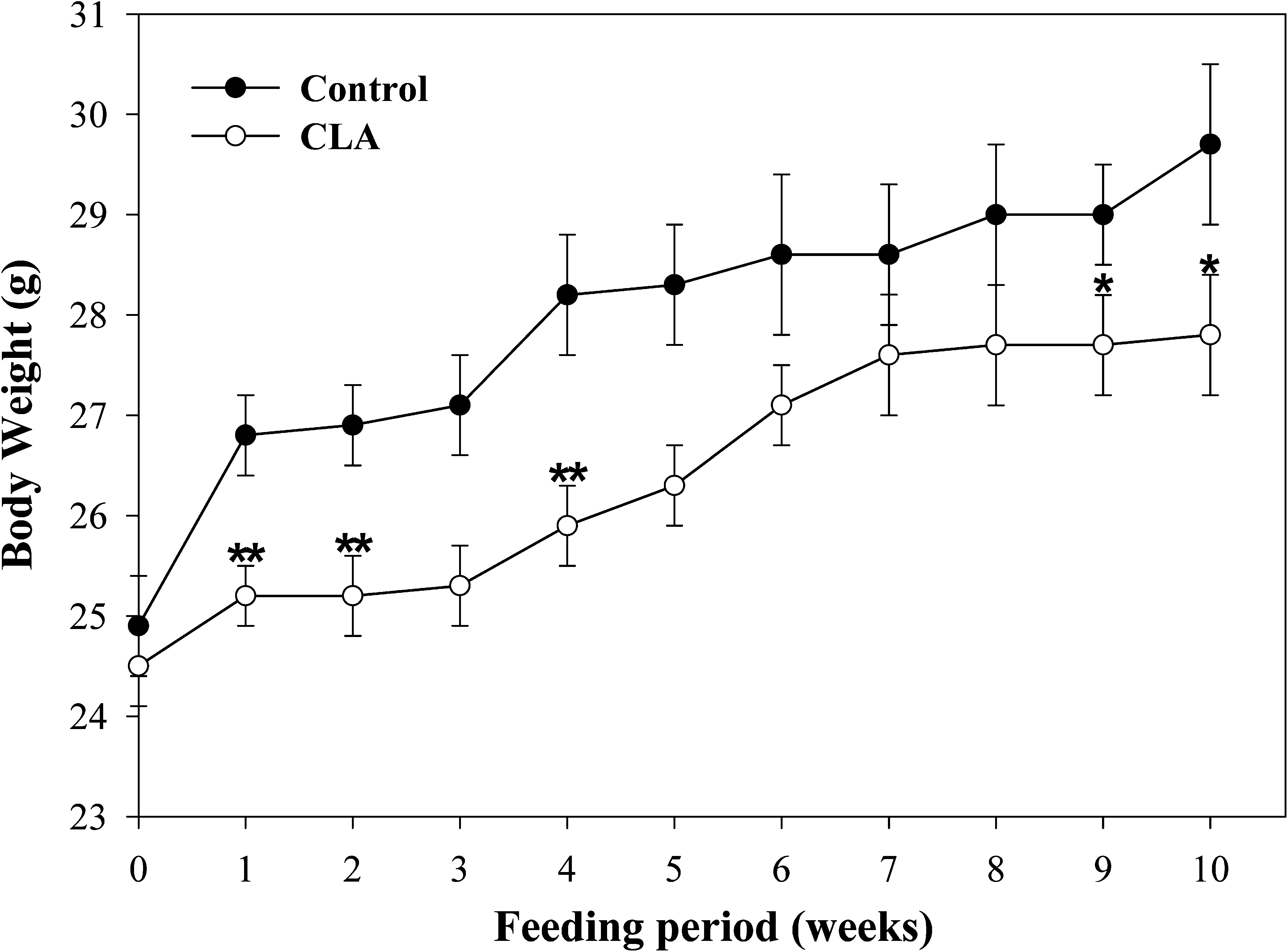
Effect of dietary CLA on body weight for 10 weeks. CLA, normal diet + 1% CLA. Data are mean ± SEM values for eight or nine animals. *P < .05, **P < .01 compared to the control group.
All mice were acclimatized to the treadmill 1 week before the maximum running time was estimated, and at the end of the experimental period the maximum running time of mice was estimated. The maximum running time for the CLA-fed mice was significantly longer, by 26%, compared to that of the control mice (P < .05) (Fig. 2).

Effect of dietary CLA on maximum running time of mice after 10 weeks. CLA, normal diet + 1% CLA. Data are mean ± SEM values for eight or nine mice. *Value is significantly different from that of the control group (P < .05). All mice were subjected to forced running in individual treadmill lanes until they were fatigued.
The concentrations of major metabolic substrates were immediately measured after endurance exercise (Table 2). The concentrations of serum triglyceride and urea nitrogen were significantly lower in the CLA-fed mice compared to mice fed a standard diet (P < .05). Supplementation of CLA decreased the serum NEFA concentration but not significantly (P > .05). The content of liver glycogen was significantly higher in the CLA group than in the control group (P < .05).
Data are mean ± SEM values for eight or nine mice.
CLA, normal diet + 1% CLA. All mice were immediately dissected after estimation of their maximum running time.
Values are significantly different from that of the control group (P < .05).
NEFA, nonesterified fatty acid.
Discussion
There have been several attempts to utilize bioactive compounds that display anti-obesity activity for improving endurance capacity. For example, oral administration of capsaicin increased the swimming endurance capacity of high-fat-fed mice by enhancing fat mobilization. 14 Fushiki et al. 10 reported that the chronic consumption of medium-chain triglycerides enhanced the endurance capacity of mice with increased utilization of fatty acids and ketone bodies. These results could be accounted for by the reduced carbohydrate consumption because of the increased use of fatty acids as a substrate for energy production during exercise. In both animal and human studies, “glycogen sparing” was accompanied by enhanced exercise endurance. 15,16 Therefore, these previous studies led us to hypothesize that CLA, an established anti-obesity bioactive compound, might have the ability to increase endurance capacity.
Swimming pool or treadmill systems have been used in many studies to determine the endurance capacity of mice. However, various factors can influence the data, including floating, climbing, continuous swimming, diving, or bobbing behaviors of experimental animals in the swimming system. 17,18 These swimming behaviors can confound the interpretation of results by introducing intermittent bouts of hypoxia. 19 Meanwhile, treadmill running has the distinct advantages over other forms of exercise, including swimming, in that the total amount of external work done by the experimental animal can be easily calculated 20 and the investigator can control both exercise intensity and duration. 19 The present study used treadmill running, which can more clearly evaluate the endurance capacity of mice. Dietary CLA significantly increased the maximum running time of mice after 10 weeks. Mizunoya et al. 9 reported that 1 week on the CLA diet significantly increased the swimming time of mice. The previous study in combination with the results obtained in this study supports the notion that dietary CLA improves endurance exercise capacity in mice.
To confirm that dietary CLA could help conserve carbohydrate stores in the liver by increasing lipid mobilization, we analyzed the concentrations of metabolic substrates in the serum and liver of mice after they had exercised until exhaustion. Serum concentration of triglycerides was significantly lowered (P < .05), and NEFA level was also decreased (P > .05) in the CLA group relative to the control group. Conversely, hepatic glycogen content was markedly higher in the CLA group than in the control group. These data suggest that dietary CLA could improve the endurance capacity in mice by increasing fat utilization and reducing the consumption of stored hepatic glycogen. Liver glycogen depletion might be an important factor in the development of fatigue because as liver glycogen is depleted during exercise, there is an inability to maintain blood glucose level, and the ensuing hypoglycemia could result in impaired nervous function. 21 Although muscle glycogen concentration was not measured in this study, our results could partially establishes a role for CLA in the modulation of energy substrate utilization during exercise.
Proteins can be catabolized to a greater degree when the body cannot obtain enough energy from carbohydrate and fat sources; accordingly, urea nitrogen concentrations increase substantially following sustained exercise. Thus, urea nitrogen is a very sensitive index for evaluating the body endurance ability. 22 In the present study, reduced serum levels of urea nitrogen with CLA supplementation reflect reduced protein metabolism, which is indicative of enhanced endurance.
In conclusion, the present study demonstrated that dietary CLA enhances the maximum running time of mice by increasing fat utilization and reducing the consumption of stored liver glycogen. In addition, the reduced urea nitrogen levels indicate increased lipid metabolism by CLA during prolonged exercise. Our results provide an important basis for developing CLA as a novel ergogenic aid. However, further research is needed to provide more information on the mechanism of action of CLA in energy metabolism, including hormonal regulation, during prolonged exercise.
Footnotes
Acknowledgment
This work was supported by a Korea University grant.
Author Disclosure Statement
No competing financial interests exist.