
Abstract
Increased oxidative stress and impaired endothelium-dependent relaxation could underlie many of the vascular complications associated with diabetes. We aimed to investigate the effect of supplementation with grape seed proanthocyanidin extract (GSPE), a natural antioxidant, on vascular responses and oxidative stress in streptozotocin-induced diabetic rats. Male Sprague–Dawley rats were divided into three groups: control rats, untreated diabetic rats, and GSPE (100 mg/kg, for 6 weeks)-supplemented diabetic rats. Thoracic aorta rings of the rats were mounted in organ baths, and relaxant responses to acetylcholine (ACh), A23187, and sodium nitroprusside (SNP) were assayed in tissues precontracted with 60 mM KCl. Plasma samples used for the measurement of malondialdehyde (MDA) level and superoxide dismutase (SOD) activity. The endothelium-dependent relaxations in response to ACh and A23187 were impaired, but endothelium-independent relaxation in response to SNP did not change in diabetic rats. Supplementation with GSPE significantly improved the relaxant responses to ACh and A23187. The MDA level was significantly elevated and the plasma SOD activity was decreased in diabetic rats, but supplementation with GSPE attenuated the elevated MDA levels and increased plasma SOD activity. Thus supplementation of GSPE may attenuate oxidative stress through the inhibition of lipid peroxidation and may restore endothelial function and reduce the risk of vascular disease in diabetes.
Introduction
D
Proanthocyanidins comprise a group of polyphenolic bioflavonoids ubiquitously found in fruits and vegetables. They are present at high levels in the seeds and skins of grapes. 4 Recently, a large number of studies have been conducted on an IH636 grape seed proanthocyanidin extract (GSPE). Different groups have studied GSPE for neuroprotective, 5 anti-inflammatory, 6 and cardioprotective 7 properties. In these studies, the antioxidant and free radical scavenging effects of GSPE have been demonstrated. Although other reports have shown the protective effect of GSPE on oxidant stress-related pathologies, the protective potential of GSPE against impaired endothelial relaxant response in streptozotocin (STZ)-induced diabetic rats has not been reported to date. Therefore, we aimed to investigate the effect of 6 weeks of oral GSPE supplementation on vascular relaxant responses and oxidative stress in STZ-induced diabetic rats.
Materials and Methods
The study protocol was approved by the Ethics Committee of the Selçuk University Experimental Medicine Research and Application Center. Eighteen adult male Sprague–Dawley rats (weighing 240±24 g) were used. Animals were divided into three groups as follows: (1) control group (nondiabetic animals) (n=6), (2) untreated diabetic animals (n=6), and (3) GSPE-supplemented diabetic rats (n=6). Diabetes was induced by a single intraperitoneal injection of 50 mg/kg STZ in sterile and fresh physiological saline solution. Age-matched control rats were injected with saline solution. A drop of tail blood was obtained at 1 week after administration of STZ to verify hyperglycemia using a glucometer (Abbott Diabetes, Alameda, CA, USA). Rats having blood glucose levels 300 mg/dL and higher were accepted as diabetic. All animals were fed standard rodent chow and tap water ad libitum. GSPE (100 mg/kg) was added to the drinking water of group 3 for 6 weeks. The GSPE supplementation started on day 7 after STZ injection. It is known that daily water consumption of rats is approximately 10–12 mL/100 g of body weight. GSPE was dissolved in a volume of tap water less than the rats consumed daily. First, rats were given tap water containing GSPE, and then water bottles were filled with tap water again. Thereafter, rats were given tap water ad libitum, and this procedure was repeated every day during the 6 weeks. The rats were housed in an aseptic animal room at a temperature of 20±2°C and a humidity of about 50% with a 12-hour light/dark cycle.
After 6 weeks of supplementation, the rats were sacrificed by cardiac puncture under kethamine/xylazine anesthesia. Plasma samples were collected and used for the measurement of MDA and superoxide dismutase (SOD) levels.
Determination of MDA
MDA levels were measured with the thiobarbituric acid reaction by the method of Wasowicz et al. 8 The quantification of thiobarbituric acid–reactive substances was determined by comparing the absorption to the standard curve of MDA equivalents generated by acid-catalyzed hydrolysis of 1,1,3,3-tetramethoxypropane. The values of MDA were expressed as μmol/L.
Determination of SOD activity
SOD activity was measured by using a commercially available kit according to instructions of the manufacturer (Cayman Chemical, Ann Arbor, MI, USA). The values were expressed as U/mL.
Vascular studies
The thoracic aorta was removed and placed in Krebs–Henseleit solution (KHS) of the following composition: 119 mM NaCl, 4.7 mM KCl, 1.5 mM MgSO4, 1.2 mM KH2PO4, 2.5 mM CaCl2, 25 mM NaHCO3, and 11 mM glucose. The aorta was cleaned of the adhering perivascular tissue and cut into rings 3 mm long. The rings were suspended in isolated tissue baths filled with 10 mL of KHS continuously bubbled with a mixture of 95% O2 and 5% CO2 at 37°C. At the beginning of the experiment, the rings were stretched to an initial tension of 1 g and allowed to equilibrate for 60 minutes in the KHS, which was changed every 15 minutes.
After the equilibration period, the rings were contracted with KCl (60 mM). After the maximal contractile response to 60 mM KCl was achieved, increasing concentrations of acetylcholine (ACh) (10−9–10−5 M), A23187 (10−9–10−5 M), or sodium nitroprusside (SNP) (10−9–10−5 M) were added cumulatively to the bath.
Drugs
GSPE (IH636), commercially known as ActiVin™, kindly donated by San Joaquin Valley Concentrates (Fresno, CA, USA), is a natural substance extracted from grape seeds with ethanol and water. ActiVin contains 54% dimeric, 13% trimeric, and 7% tetrameric proanthocyanidins and approximately 8% monomeric bioflavonoids. STZ, KCl, A23187, and SNP were purchased from Sigma Chemical Co. (St. Louis, MO, USA).
Data recording and analysis
In all groups, isometric recording of tension changes was obtained with force transducers (Grass FTO4, Grass Instrument Co., West Warwick, RI, USA) connected through amplifiers to a polygraph (Grass 7D, Grass Instrument Co.).
The relaxation responses to ACh, A23187, and SNP were calculated as the percentage of initial preconstruction to KCl. Values for the percentage of the maximal response of the tissue (E max) and the negative log of the concentration of a drug required to produce 50% of the maximal relaxation (pEC50) were calculated for each artery.
Results were expressed as mean±SD values. One-way analysis of variance and Bonferroni's correction was used. A value of P<.05 was considered significant.
Results
Control animals gained weight from 227±23 to 262±35 g at the end of the study, whereas the two groups of diabetic animals weighed 254±25 and 225±30 g at the start and end of the study, respectively. Diabetic animals had lower body weights than those in the control group (P<.05). After GSPE supplementation neither body weight nor glycemia differed significantly compared with untreated diabetic rats (Table 1).
Data are mean±SD values (n=6).
P<.05 compared with the control group.
GSPE, grape seed proanthocyanidin extract.
KCl (60 mM) induced contractions in aortic rings from control and diabetic rats. ACh (10−9–10−5 M), A23187 (10−9–10−5 M), and SNP (10−9–10−5 M) induced concentration-dependent relaxations in precontracted aortic rings from control and diabetic rats (Fig. 1). The relaxant response to ACh and A23187 significantly decreased in aortas of diabetic rats fed with normal diet, without a marked change in the sensitivity. The rings of diabetic rats supplemented with GSPE showed a significantly enhanced relaxant response to ACh and A23187 compared with the untreated diabetic rings; the latter relaxations were not significantly different from the control group (Fig. 1). Relaxant responses to SNP in aortic rings were not significantly changed by diabetes or GSPE supplementation of diabetic rats compared with control rings (Fig. 1). E max and pEC50 values for ACh, A23187, and SNP in control, diabetic, and GSE-supplemented diabetic groups are given in Table 2.
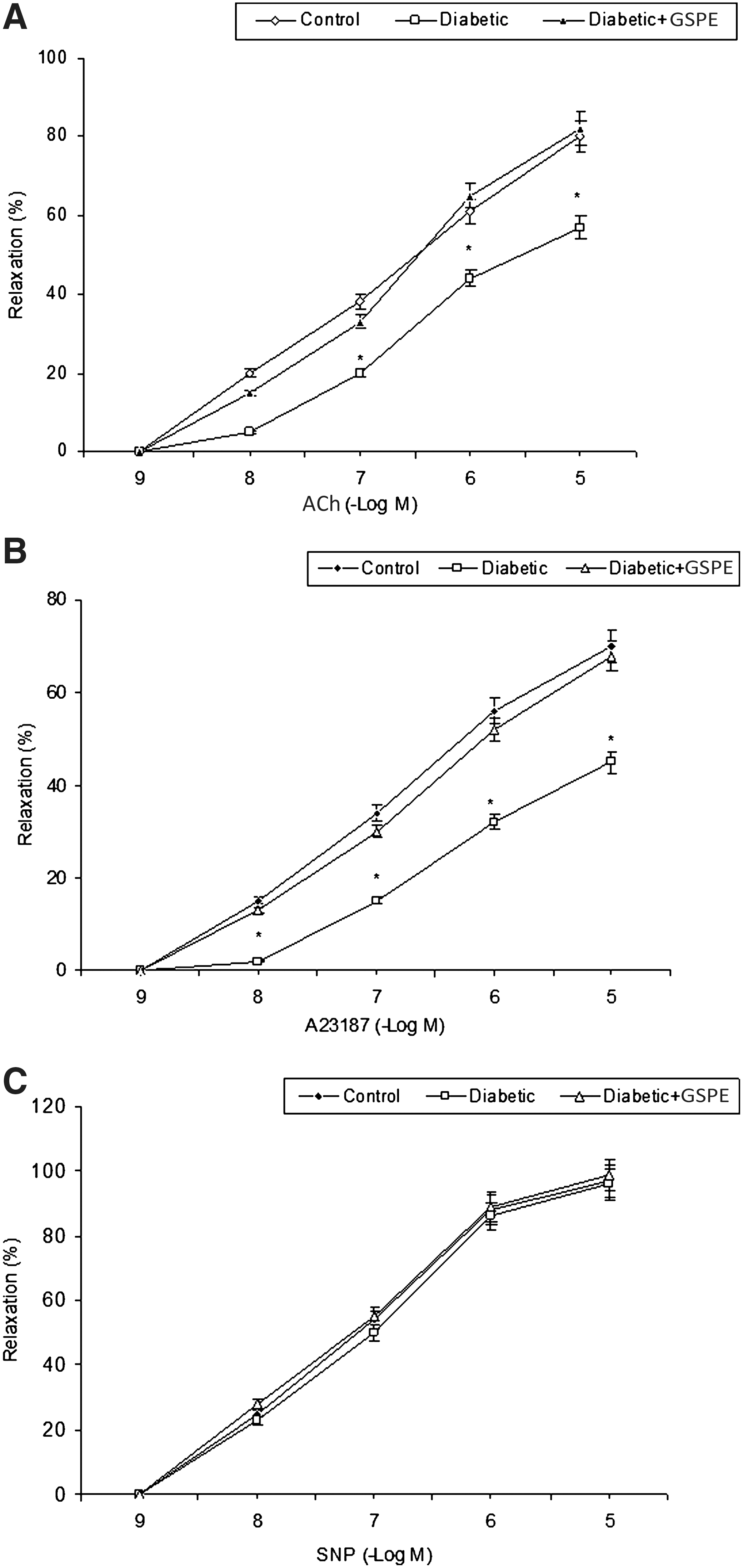
Relaxation induced by (
Data are mean±SD values (n=6 per tissue).
P<.05 compared with the control group, † P<.05 compared with the diabetic group.
E max, percentage of the maximal response of the tissue; pEC50, negative log of the concentration of a drug required to produce 50% of the maximal relaxation.
As shown in Figure 2A, the serum MDA level was significantly higher in diabetic rats (1.896±0.539 μmol/L) compared with that of control rats (1.295±0.053 μmol/L). Supplementation with GSPE reversed the MDA level back to the control level (1.462±0.139 μmol/L). Serum SOD activity showed a marked reduction in the STZ-induced diabetic group (0.783±0.067 U/mL) compared with the control group (0.903±0.054 U/mL). Similarly, GSPE supplementation increased the reduction in SOD activity (0.903±0.058 U/mL) to levels that were close to control values (Fig. 2B).
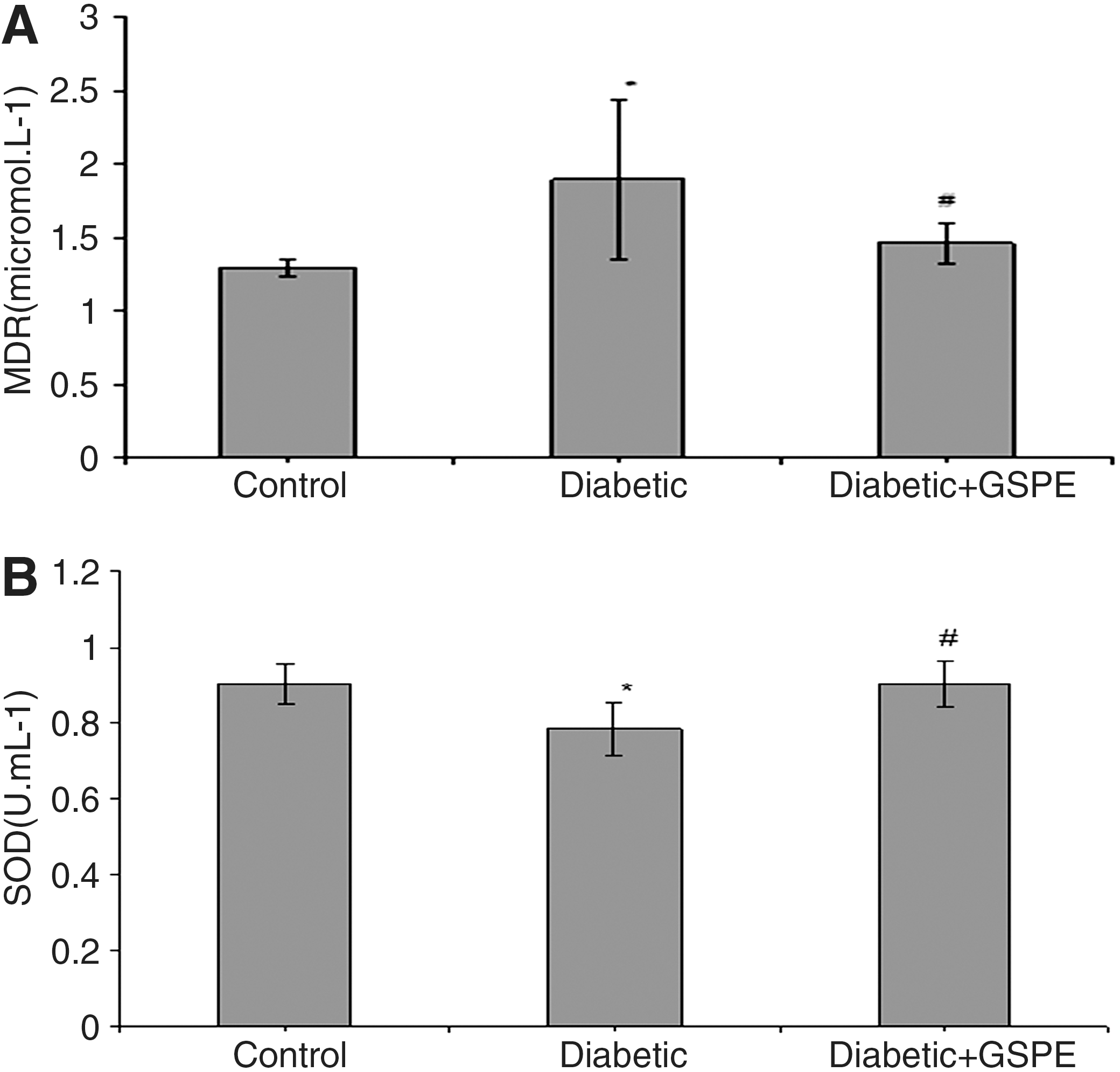
(
Discussion
Our study showed that oral supplementation with GSPE for 6 weeks resulted in (1) an improvement in endothelium-dependent relaxation to ACh and A23187, (2) a pronounced reduction in plasma level of MDA, and (3) an increase in SOD activity in STZ-induced diabetic rats.
Hyperglycemia, a characteristic of diabetes, increases intracellular ROS. Several studies have shown a reduction of NO and an increase in superoxide formation by the vessels of diabetic laboratory animals. 9 –11 Superoxide reacts readily with NO to produce peroxynitrite, a potent oxidant, which in turn stimulates lipid peroxidation and endothelial dysfunction. 12 The process of lipid peroxidation is one of the oxidative conversion of polyunsaturated fatty acids to products known as MDA, a highly toxic molecule. 3 The increase of MDA level in plasma suggests enhanced lipid peroxidation leading to tissue damage and failure of antioxidant defense mechanisms. 13
In metabolism, the existence of antioxidant systems protects the cell membrane against damage caused by free radicals. SOD is an enzymatic antioxidant that reduces superoxide radical to hydrogen peroxide and oxygen. Therefore, SOD activity is important for the regulation of oxidative status in diabetes. 14 The overproduction of superoxide in diabetic animals has been ascribed, in part, to a reduction in endogenous vascular SOD activity. 11 In our experimental model, induction of diabetes resulted in apparent oxidative conditions. STZ-induced diabetes caused a significant increase in plasma level of MDA, whereas the activity of SOD was significantly decreased. Similar results have been reported by other researchers in STZ-induced diabetic rats. 13,15 Furthermore, diabetes induced marked alterations in aortic vascular responses in rat aorta rings. The relaxation responses of aortic rings to the endothelium-dependent agents ACh and A23187 are weaker in STZ-induced diabetic rats than in normal rats in the present study. The responses to the endothelium-independent vasodilator SNP did not change. These data are consistent with previous reports of impaired endothelium-dependent relaxation in arteries from diabetic individuals. 15
The endothelium is an important modulator of vascular tone and function through the synthesis and release of endothelium-derived NO. Endothelial NO synthesis can be stimulated by receptor-dependent agonists and receptor-independent agonists. 16 Increased vascular production of superoxide anion contributes importantly to impaired endothelium-dependent vascular relaxation in animal models of diabetes. 17 Determination of vasodilatation in response to ACh in an isolated blood vessels has been used to infer stimulated NO-mediated vasodilatation in animal models with diabetic vascular disease. 18 Impaired ACh relaxation in vascular tissues from diabetic rats and humans may represent a hallmark of early endothelial dysfunction, as indicated by attenuated agonist stimulated release of NO. 19 Restoring endothelial function has important clinical implications for reducing the risk of vascular disease in diabetic patients. Several studies showed that some antioxidants are effectively used to protect against diabetes-induced endothelial damage. 20,21
In this study, we used GSPE (ActiVin, IH636), an antioxidant derived from grape seeds. This extract mainly contains novel oligomeric proanthocyanidins and small amounts of monomeric flavonoids. GSPE exhibited free radical scavenging abilities toward biologically generated free radicals such as superoxide anion, hydroxyl radicals, and peroxyl radicals. 4 Additionally, it has also been shown that GSPE significantly inhibits the activities of free radical–producing enzymes and changes antioxidant enzyme activity in biological systems. 22,23 For instance, Fuji et al. 24 reported that GSPE had potent protective effects against high glucose-induced oxidative stress in cultured LLC-PK1 cells. In the study of Balu et al. 25 it was reported that GSPE (100 mg/kg for 30 days) enhanced the antioxidant status and decreased the incidence of free radical–induced lipid peroxidation in the central nervous system of aged rats. In another study GSPE (50 and 100 mg/kg for 72 hours) significantly increased pancreatic glutathione levels and inhibited the increase in lipid peroxidation caused by alloxan. 26 In this study, the decrease of ACh- and A23187-induced endothelium-dependent relaxation in thoracic aorta from STZ-induced diabetic rats was reversed by GSPE supplementation. Release of NO by ACh is a muscarinic receptor-operated effect, whereas A23187 works in a receptor-independent manner. 20 The endothelium-independent relaxation responses to SNP in thoracic aorta from STZ-induced diabetic rats were not changed. However, the amount of MDA formation was significantly reduced in the plasma of the STZ-diabetic rats after GSPE supplementation. The lower plasma MDA concentration in the GSPE-supplemented group is consistent with reports that dietary GSPE had a strong radical scavenging activity. 13
In GSPE-supplemented diabetic rats, we found a moderate and significant increase in SOD activity. There are discrepancies in the literature about the effect of GSPE on SOD activity. Ahn et al. 27 showed that catalase and SOD activities were increased by GSPE administration in liver of rats. In an another study, there was no significant change in the basal levels of myocardial SOD and catalase on administration of GSP, but the reduction in SOD and catalase activities in the isoproterenol-induced myocardial injury group was normalized by GSPE supplementation. 28 On the other hand, Puiggros et al. 29 reported that increased Cu/Zn-SOD activity in diabetic GSPE rats is not significantly different from the activity in diabetic rats, but Cu/Zn-SOD activity increased significantly in the nondiabetic GSPE group relative to the nondiabetic group. They suggested that GSPE did not alter the Cu/Zn-SOD mRNA levels or the amount of protein, but the enzyme activity was significantly enhanced by direct interaction between molecules in GSPE and the enzyme. Based on our results we cannot comment on whether the moderate SOD up-regulation is responsible for the reduction of superoxide; however, future studies may facilitate the appreciation of the antioxidant effect of GSPE.
In summary, our present study demonstrated that GSPE supplementation may attenuate oxidative stress through the inhibition of lipid peroxidation and may normalize plasma SOD activity, restoring vascular endothelial function in STZ-induced diabetic rats. Hence, GSPE may reduce the risk of vascular endothelial complications in diabetes.
Footnotes
Author Disclosure Statement
No competing financial interests exist.