
Abstract
Medicinal plants represent an important resource in new drug research. Antioxidant properties of plants can help to scavenge reactive oxygen species. The objective of this work was to evaluate the genotoxic, antigenotoxic, tumoricidal, and apoptotic effect of some major phenols (apigenin, bisabolol, and protocatechuic acid) from two medicinal plants, Matricaria chamomilla and Uncaria tomentosa. The wing spot test of Drosophila melanogaster was used to evaluate the genotoxicity and antigenotoxicity of the three phenols. The human model of HL-60 leukemia cells was used for the assessment of the cytotoxic effect, growth, and cellular viability. The apoptotic effect was evaluated using a DNA fragmentation assay based on the formation of internucleosomal units. Protocatechuic acid (0.25 and 1 mM), apigenin (0.46 and 1.85 mM), and bisabolol (0.56 and 2.24 mM) did not exhibit any genotoxic effect. The three phenols showed an antigenotoxic effect against the hydrogen peroxide effect and also exhibited tumoricidal activity. Apigenin (2.24–35.96 mM) showed a lower 50% inhibitory concentration (0.75 and 3.87 mM for the trypan blue test and WST-8 colorimetric assay, respectively) than bisabolol and protocatechuic acid. These phenolics also induced apoptosis in HL-60 leukemia cells. This study suggests that the antioxidant activity of Chamomilla and Uncaria could be partially responsible of their beneficial activity.
Introduction
M
Bisabolol is one of the most important sesquiterpenes of M. chamomilla, representing up to 50% of its total phenolic composition. It has been shown to be nonmutagenic by the Salmonella microsomal test, and it can even neutralize the effect of various mutagenic substances 8 and also induces apoptosis in rat glioma cells. 9
Apigenin is a nontoxic flavonoid with an antitumoral effect. This phenol inhibits the development of cancer and induces apoptosis on various tumoral cell lines. Apoptosis induction with apigenin generally occurs by caspase-3 activation, liberation of cytochrome c, and generation of reactive oxygen species. 10
Protocatechuic acid is present in fruits, vegetables, and some medicinal plants. 11 This phenol can exhibit a contradictory biological activity: it displays an anticarcinogenic activity in liver 12 and induces apoptosis in human leukemia cells via the reduction of retinoblastoma phosphorylation and Bcl-2 expression, 13 whereas in other cases this phenol has been shown to promote cancer development on skin. 14
Chemoprevention refers to the administration of synthetic naturally occurring agents to prevent initiation and promotional events associated with carcinogenesis. This effective approach for the management of neoplasm is increasingly appreciated. Genotoxicity/antigenotoxicity and cytotoxicity tests are rapid methods to assess the innocuousness and possible beneficial effects of single compounds or complex mixtures. The objective of this study was to investigate the genotoxic/antigenotoxic effects of some main phenolic compounds of M. chamomilla and U. tomentosa (i.e., bisabolol, apigenin, and protocatechuic acid) against H2O2 as a genotoxicant in order to evaluate their potential DNA protection. The somatic mutation and recombination test (SMART) with Drosophila melanogaster is a suitable eukaryotic tool for genotoxicity and antigenotoxicity studies. 15 Cytotoxicity assays with HL-60 leukemia cells were used to investigate the cytotoxic effect of substances already genetically evaluated. On the basis of the observed cytotoxic effects, further studies of the phenols' mechanisms of action are needed. A next step would be to detect an ordered cleavage of DNA that constitutes a hallmark of apoptosis.
Materials And Methods
Phenolic compounds
The three phenolic compounds used in this study—bisabolol, apigenin, and protocatechuic acid—were purchased from Sigma (St. Louis, MO, USA) (catalog numbers 14462, P5630, and 10798, respectively). The phenols were dissolved in ethanol and filtered before use.
Genotoxicity and antigenotoxicity tests
The assays were essentially performed as described by Graf et al. 15 using the standard cross. The flies carry wing genetic markers on the left arm of chromosome 3: multiple wing hair (mwh, 3–0.3) and flare (flr 3 , 3–38). Hybrid eggs derived from crossing optimally fertile flies were collected over an 8-hour period. Larvae that emerged 72 ± 4 hours later were cleaned from remaining feeding medium with 20% sodium chloride solution and subsequently transferred to treatment vials. These vials contained 0.85 g of Drosophila Instant Medium (formula 4-24, Carolina Biological Supply, Burlington, NC, USA) wetted with 4 mL of the phenol solutions at physiological concentrations for D. melanogaster (0.46 and 1.85 mM apigenin, 0.56 and 2.24 mM bisabolol, and 0.25 and 1 mM protocatechuic acid). Concurrent negative controls with the solvent alone (water) and positive controls with H2O2 (120 mM) were also assayed. Antigenotoxicity tests were carried out by mixing the mutagen (H2O2, 120 mM) with the phenolic solution. In order to explain the mechanism of action of phenols, two additional treatments were carried out: pretreatment with H2O2 for 2 hours followed by a subsequent treatment with the phenols until pupation, and vice versa. After the 2-hour pretreatment, the larvae were washed out and then transferred to newly prepared medium containing either H2O2 or the phenol only.
After emergence, adult flies were collected from the treatment vials and stored in 70% ethanol. Transheterozygous marker wings (mwh flr+/mwh+ flr 3 ) were mounted, and wing hair mutations (spots) were scored using a photonic (Leica CME, Leica Microsistemas S.L.U., Barcelona, Spain) microscope (×400).
Data evaluation and statistical analysis
The frequencies of spots per fly of treated series were compared with its corresponding negative control series (water) to analyze SMART data. For maximum power, statistical analyses were done exclusively for the total number of spots recovered. The nonparametric U test of Mann, Whitney, and Wilcoxon was used to avoid weak positive and inconclusive results on SMART and to achieve a well-defined statistical diagnosis whether a given treatment should be regarded as genotoxic or not. 16
The inhibition percentage (IP) values were calculated using the total spots per wing with the following formula:
17
Cytotoxicity tests
The human leukemia HL-60 cell line was kindly provided by Dr. C. Gómez, Department of Cell Biology, University of Córdoba, Córdoba, Spain. Cells were cultured in suspension at 37°C in RPMI 1640 medium (catalog number BE12-167F, BioWhittaker, Walkersville, MD, USA) supplemented with 10% fetal bovine serum (catalog number DE14-801F, BioWhittaker), glutamine (catalog number G7513, Sigma), and an antibiotic–antimycotic solution (catalog number A5955, Sigma) in a humidified atmosphere containing 5% CO2. The cultures were plated at a density of 2.5 × 104 cells/mL in 10 mL of culture medium and were passaged every 2 days. The cytotoxic potential of different concentrations of apigenin (0.23–3.71 mM), bisabolol (2.24–35.96 mM), and protocatechuic acid (1.62–25.95 mM) was evaluated. Cells were seeded at 105 cells/mL in 12- or 96-well plates and incubated with the different concentrations of phenols for 72 hours for trypan blue exclusion assay (catalog number 93595, Sigma) and 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium (WST-8) assay (catalog number 96992, Sigma), respectively.
The reactivity of trypan blue, a vital dye, is based on the fact that the chromopore is negatively charged and does not interact with the cell unless the membrane is damaged. Therefore, all the cells that exclude the dye are viable. Nonviable cells stained purple–violet, whereas viable ones remained unstained. Trypan blue was added to cell cultures at a 1:1 ratio, and 20 μL of cell suspension was loaded into a Neubauer chamber. The cells were counted under an inverted microscope at ×100 magnification. Curves are expressed as survival percentage with respect to control growth at 72 hours, and 50% inhibitory concentration (IC50) values were estimated. All data come from at least three independent experiments.
The WST-8 assay is based on the extracellular reduction of the highly water-soluble tetrazolium salt WST-8 by NADH produced in the mitochondria via trans-plasma membrane electron transport and an electron mediator. The amount of formazan dye generated by the activity of dehydrogenases in cells is directly proportional to the number of living cells. The WST-8 assay was used according to the manufacturer's instructions. In brief, HL-60 cells, at 2 × 104 cells/200 μL of culture medium, and various concentrations of apigenin, bisabolol, and protocatechuic acid were placed in 96-well microtiter plates. After a 72-hour incubation, 20 μL of WST-8 solution was added to each well, and incubation was continued for 1 hour at 37°C. The plates were shaken for 10 minutes, and the absorbance was read at 450 nm in a microplate reader. Comparisons of a group's data for trypan blue assay and WST-8 results were done by one-way analysis of variance. In the case of significant differences, Dunnett's test was used to compare the control series with the treated ones. The IC50 of the three phenols was calculated for both cytotoxic assays.
Assessment of apoptosis
HL-60 cells (1.5 × 105/mL) were treated with apigenin (0.23–3.71 mM), bisabolol (2.24–35.96 mM), and protocatechuic acid (1.62–25.95 mM) for 5 hours. An untreated cell control was included for each experiment. Cells were centrifuged at 197 g for 5 minutes, washed with ice-cold phosphate-buffered saline, and pelleted. The DNA was extracted using a commercial kit (catalog number MBL 243, Dominion mbl, Córdoba, Spain), and the resulting total DNA was treated with RNase for 30 minutes at 37°C. Samples of 1.5 μg of DNA were mixed with loading buffer and loaded onto a presolidified 2% agarose gel containing ethidium bromide. The agarose gels were electrophoresed at 50 V for 90 minutes in TBE buffer and then observed and photographed under ultraviolet light.
Results
Genotoxicity and antigenotoxicity analysis of apigenin, bisabolol, and protocatechuic acid
The results of chronic treatment of larvae with different concentrations of phenols alone or combined with one fixed concentration (120 mM) of H2O2 in the Drosophila wing spot assay (SMART) using standard cross are presented in Table 1. Negative (water) and positive (H2O2) controls were included in each experiment. The statistical analysis showed that H2O2 was genotoxic at a concentration of 120 mM. H2O2 behaved as a gentoxin inducing oxidative stress and DNA damage, increasing the mutation frequency (0.55) in the SMART test.
Data were evaluated by the nonparametric U test of Mann, Whitney, and Wilcoxon.16
NS, nonsignificant (P > .05); *significant (P < .05).
H2O2, hydrogen peroxide; m, multiplication factor for the assessment of significantly negative results.
The genotoxicity assay confirmed that apigenin is safe for the integrity of DNA because the difference between the mutation frequencies and the negative control was not statistically significant. The results obtained in the combined treatment of larvae with apigenin (0.45 and 1.85 mM) and H2O2 showed nonsignificant differences compared with the negative control (Table 1). The inhibition percentage of genotoxicity by apigenin ranges from 34.36% to 54.54% at 0.45 and 1.85 mM, respectively (Fig. 1). This compound would act as an antimutagen against H2O2 in SMART. The antimutagenic capacity of apigenin has never been tested on the proliferative somatic cells of D. melanogaster until the present study.
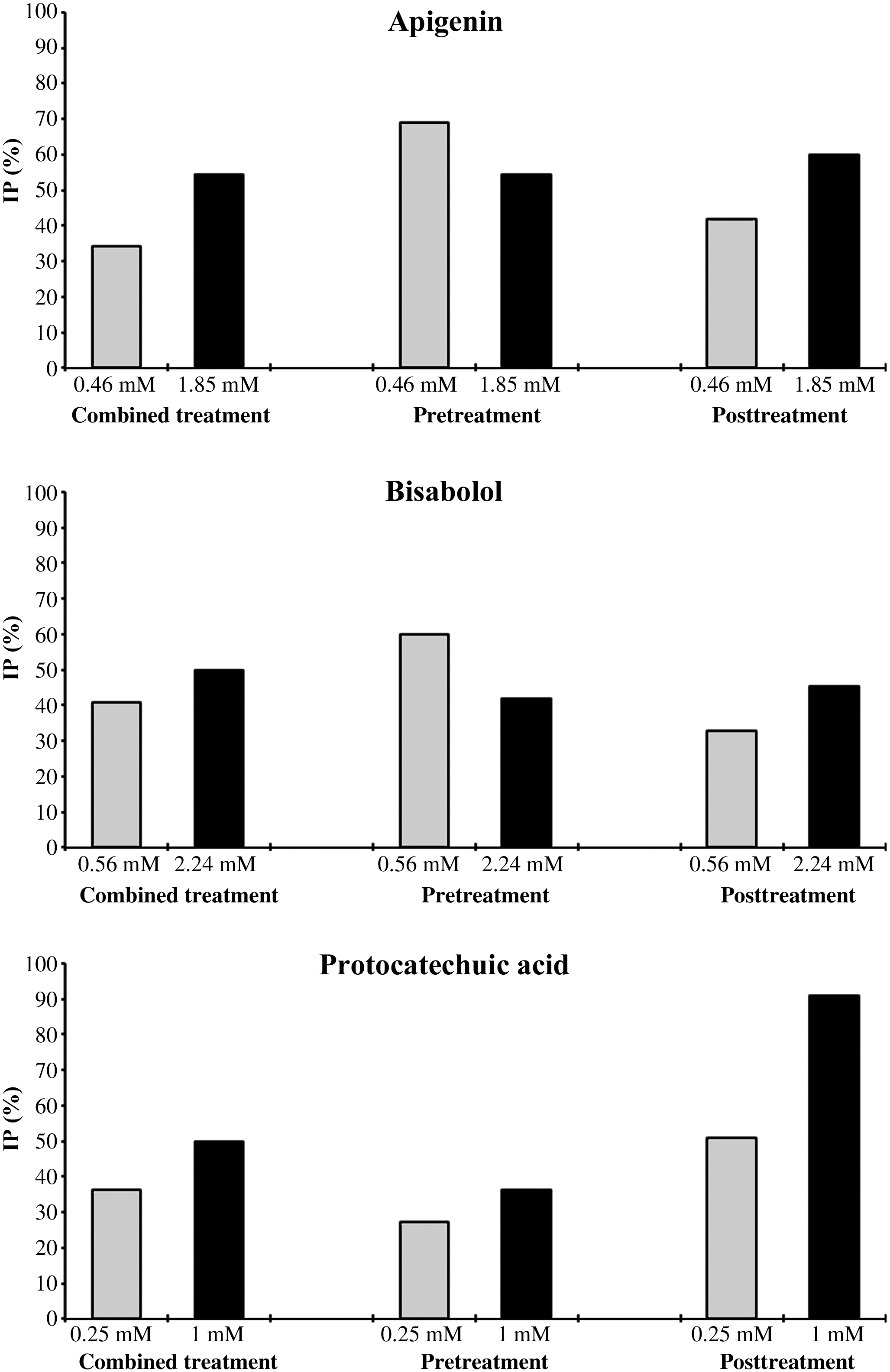
Inhibition percentage (IP) values of apigenin, bisabolol, and protocatechuic acid against H2O2-induced damage in the Drosophila wing spot test. Larvae at 72 hours were co-treated with H2O2 (120 mM) and the compounds (combined treatment), pretreated with the compounds for 2 hours followed by a subsequent treatment with H2O2 until pupation (pretreatment), or the reverse (post-treatment).
The genotype mwh/flr 3 revealed nonsignificant results in the simple treatment (Table 1) by bisabolol, being not genotoxic at concentrations of 0.56 and 2.24 mM. Furthermore, when larvae were co-treated with H2O2, this phenol was antigenotoxic, and it was able to avoid the damage caused by the gentoxin. The statistical diagnosis was not significant when the combined treatment was compared with the negative control. Inhibition percentages were 40.40% and 50% at 0.56 and 2.24 mM, respectively.
Table 1 shows the data obtained in SMART for both concentrations of protocatechuic acid (0.25 and 2.24 mM). The difference was not significant when the frequencies of total spots were compared with the negative control ones. Protocatechuic acid exhibited an antigenotoxic effect when larvae were co-treated by protocatechuic acid and H2O2. The mutation frequencies at 0.25 and 1 mM (0.35 and 0.275, respectively) were smaller than that obtained with the positive control (0.55). Nonsignificant statistical differences were found between the combined treatment and the negative control. This means that protocatechuic acid exerted a desmutagenic activity against H2O2. This acid was shown to inhibit up to 50% of the genotoxic activity of H2O2 in the combined treatment.
The pretreatments with phenol/mutagen and mutagen/phenol give us an idea whether the beneficial effects of the studied phenols are previous or subsequent to the treatment with the mutagen, which cannot be discerned in a combined treatment.
The pretreatment with apigenin (0.46 mM) and bisabolol (0.56 mM) for 2 hours followed by post-treatment of larvae with H2O2 displayed mutation frequencies smaller than that obtained in the combined treatment (Table 2). The inhibition percentages were 69.09% and 60%, respectively (Fig. 1). The effect of protocatechuic acid was more moderate with inhibitions of 27.27% and 36.36% at 0.25 and 1 mM (Fig. 1), respectively.
Data were evaluated by the nonparametric U test of Mann, Whitney, and Wilcoxon. 16
NS, nonsignificant (P > .05); *significant (P < .05).
m, multiplication factor for the assessment of significantly negative results.
As shown in Table 2, the results were different in pretreatments of larvae with H2O2 for 2 hours. Protocatechuic acid showed a significant decrease of mutation frequencies (0.27 and 0.05 at 0.25 and 1 mM, respectively). The inhibition percentages were 50.90% and 90.90% (Fig. 1). Apigenin exhibited a maximal inhibition of 60% at 1.85 mM, and bisabolol inhibited by 45.45% at 2.24 mM.
Effects of apigenin, bisabolol, and protocatechuic acid on tumoral cell growth and apoptosis
Cultured mammalian cells provide an important tool for evaluating the cytotoxicity of compounds with potential therapeutic activity. 18 The cytotoxicity assessment of apigenin, bisabolol, and protocatechuic acid involved two different assays: trypan blue and WST-8.
Apigenin showed a tumoricidal effect in the trypan blue exclusion assay with an IC50 of only 0.75 mM, bisabolol also showed a cytotoxic effect with an IC50 of 6.25 mM, and protocatechuic acid was tumoricidal with an IC50 of 4.54 mM (Fig. 2). The differences between most concentrations of the three phenols and the their respective control groups were statistically significant when Dunnett's test was applied.
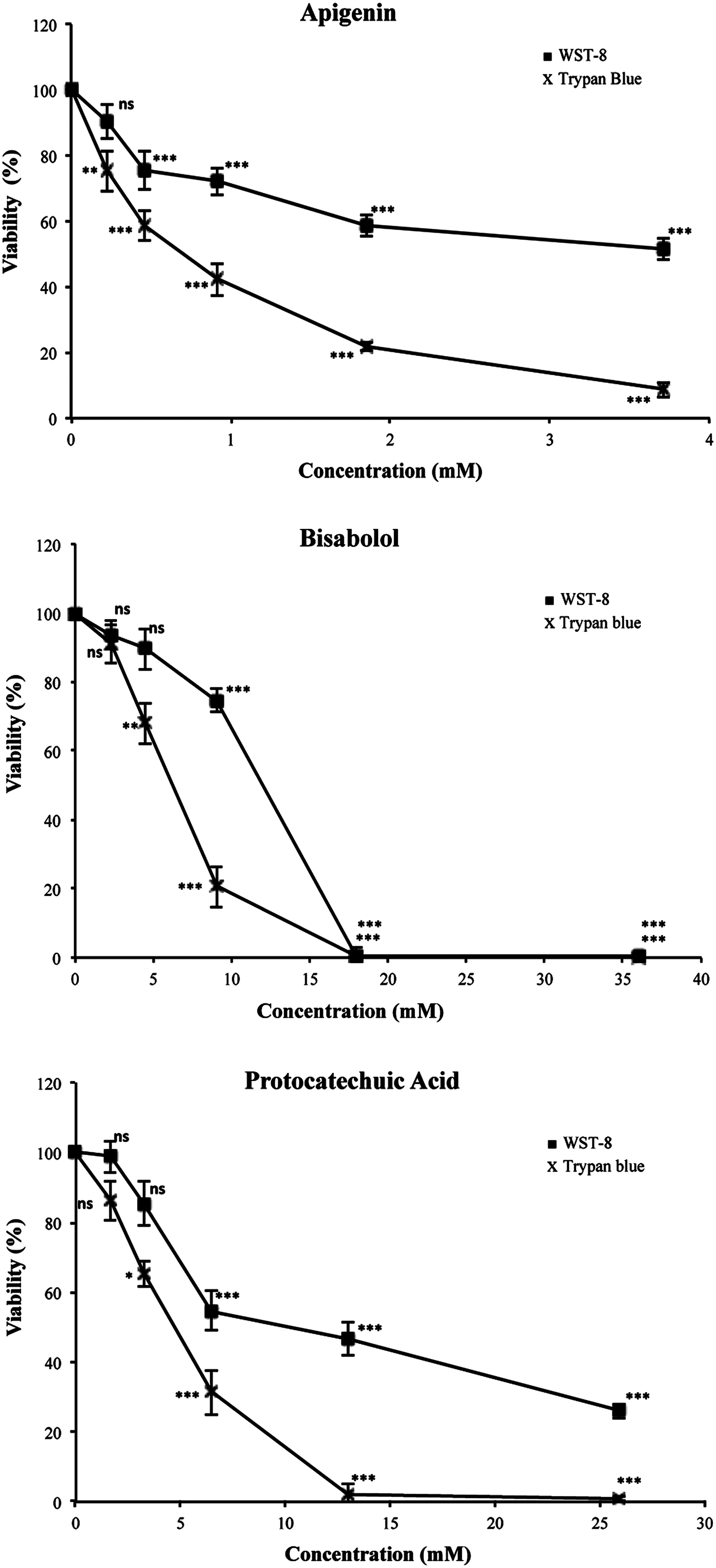
Effects of apigenin, bisabolol, and protocatechuic acid on viability of HL-60 leukemia cells. Cell viability was assessed after 72 hours by trypan blue exclusion test and 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium (WST-8) assay. Data are expressed as percentages of control (mean ± SD values from three independent experiments). ns, nonsignificant; *P < .05, **P < .01, ***P < .001.
Cell viability with the WST-8 assay was also determined by measuring the degradation of tetrazolium salt by mitochondrial dehydrogenation enzymes. The respective IC50 values of apigenin, bisabolol, and protocatechuic acid were 3.87, 11.87, and 12.72 mM. The differences between the results of trypan blue and WST-8 assays could be attributed to the fact that WST-8 can be reduced by phenolics, which can sometimes lead to misinterpretation of results. The comparison of means by Dunnett's test showed that the differences between the treatment and the control groups are significant (Fig. 2).
A hallmark of apoptosis is the degradation of genomic DNA into oligonucleosomal fragments. This DNA fragmentation end point was analyzed by conventional agarose gel electrophoresis. As shown in Figure 3, the high concentrations of apigenin, bisabolol, and protocatechuic acid showed an internucleosomal fragmentation ladder pattern of DNA for treated HL-60 cells. This did not occur in the control and at some lower concentrations. A dose-dependent effect is suggested.
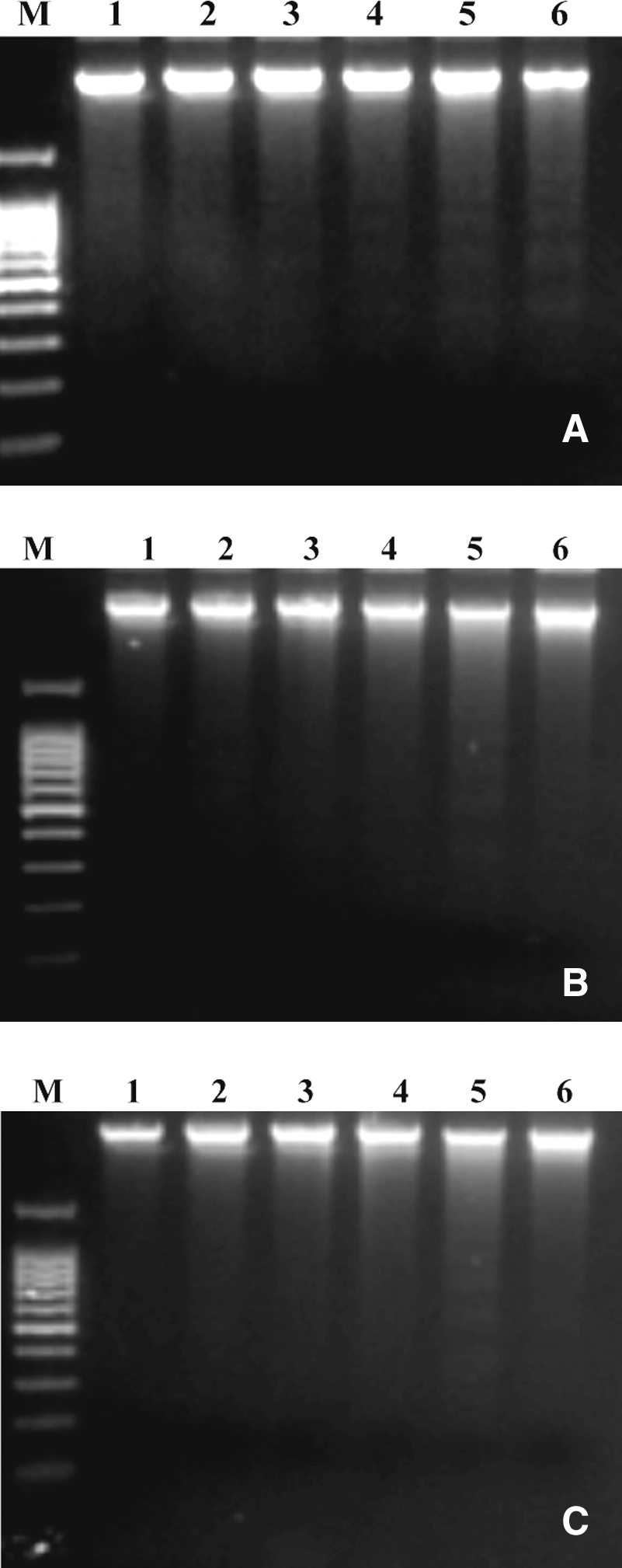
Nucleosomal DNA fragmentation. HL-60 leukemia cells were exposed to various concentrations of (
Discussion
The oxidative stress caused by H2O2 can trigger the expression of more than 100 genes, 19 inducing DNA single-strand breaks, gene mutations, and chromosomal aberrations through oxygen-radical mechanisms via the Fenton reaction. 20 H2O2 as a genotoxin induced DNA damage and a significant increase of mutation frequencies (0.55) in SMART. Nevertheless, the epigenetic interactions that can occur between highly oxidative elements (H2O2) and highly antioxidant elements (phenols) acting mutually are not known.
The genotoxicity assay confirmed that apigenin is safe in SMART as the difference between the negative control and the treatment was not significant at any concentration. This phenol also exhibited an antigenotoxic effect and behaved as an antimutagenic agent against H2O2 in SMART with an inhibition of 54.54% (1.85 mM). It is known that phenols detoxify and act as final acceptor molecules or terminators of free radicals. Apigenin displayed an antigenotoxic effect against Salmonella typhimurium, 21 although Noel et al. 22 detected clastogenic effects in the mouse micronucleus test at high doses. Based on the results of previous studies with phenols and our results in SMART, we can infer that apigenin acts as an inactivator of the reactive oxygen species at the studied doses.
Bisabolol was not mutagenic and partially inhibited the damage caused by H2O2. The inhibition percentage was 50% (2.24 mM). These results agree with those of Gomes-Careneiro et al., 8 who showed that bisabolol was not mutagenic and also indirectly induced the reduction of mutagenic effects in the Salmonella/microsomal system. The results of Hernández-Ceruelos et al. 5 reported that camomile oil blocks the sister chromatid exchange induced by daunorubicin and methylmethanesulfonate in bone mouse cells.
Protocatechuic acid was not genotoxic and exhibited a protection percentage of 50% (1 mM) against H2O2. This phenol is one of the most important simple phenolic components of U. tomentosa, 7 and it is thought that the beneficial effects on health of this plant are associated with its antioxidant activity. 23 Protocatechuic acid inhibited the carcinogenic effect induced by certain chemical agents in different organs. 13,14 However, the biological effects of protocatechuic acid are contradictory. In other studies this phenol induced alterations at the chromosomal level in Chinese hamster ovary cells. 24 This leads us to infer that the effect of protocatechuic acid was dose-dependent and that the compound is capable of exhibiting antioxidant or pro-oxidant activities accordingly. 25
With the aim of investigating in more detail the mechanisms of action of the phenols studied, we developed another type of antigenotoxicity test. These tests involved treating the larvae either previously with the mutagen or with phenol for 2 hours and later continuing the chronic treatment with the phenol or mutagen. The pretreatments, with phenols and mutagen (H2O2), give us an idea if the beneficial effects of the phenols studied are previous or subsequent to the treatment with the mutagen, which cannot be discerned in a combined treatment. Pretreatments with apigenin and bisabolol exhibited inhibition percentages (69.09% at 0.46 mM and 60% at 0.56 mM, respectively) higher than that shown in the combined treatment with H2O2. The antimutagenic effect was more significant, reducing the number of single spots and total number of spots. We suggested that this could be due mainly to a direct protective mechanism by scavenging free radicals produced by H2O2 and in a lesser extent to inhibition of enzymes, including oxidases and oxygenases. The percentage of inhibition with the post-treatment by protocatechuic acid (90.09%) is higher than the pretreatment one (36.36%). This phenol seems to exhibit a protection against H2O2-induced damage mainly by enzyme inhibition pathways. 14
Our data indicated that the phenols were not able to return H2O2-induced DNA damage to negative control levels but did reduce significantly the genotoxic effect caused by the genotoxin.
Apigenin showed a cytotoxic effect in the HL-60 viability assay; our results corroborated those of various studies. The IC50 values of apigenin were the lowest (0.75 and 3.87 mM for the two cytotoxicity assays) after a 72-hour incubation. Cárdenas et al. 26 detected selective cytotoxic activity of apigenin against tumoral cells but not against normal cells. Apigenin inhibited the promotion of tumors and induced apoptosis in cancerous cells. This phenol induced apoptosis via caspase-3 with liberation of cytochrome c. 10 Bisabolol showed a tumoricidal effect in the HL-60 viability assay with IC50 values of 6.25 and 11.87 mM (in trypan blue and WST-8 tests, respectively). In previous works this sesquiterpenic alcohol was able to kill some human cancerous lines quickly, including glioma cells. 9 The tumoricidal effect of this phenol in HL-60 leukemia cells can be explained by apoptosis induction. Bisabolol exhibited apoptotic activity in glioma cells and induced liberation of cytochrome c and the induction of apoptosis via Fas receptor. The action of this phenol seems to be selective as its effect in normal mouse astroglial cells was not cytotoxic. 27 The data obtained in the treatment with protocatechuic acid indicated that viability of HL-60 cells was significantly reduced in a concentration-dependent manner with IC50 values of 4.54 and 12.72 mM. This phenol was apoptotic and cytotoxic in cancerous breast cells. 28 However, at high concentrations, this phenol induced oxidative stress in human leukemia cells. 14 In previous studies it has been shown that the effect of protocatechuic acid is dose-dependent, exhibiting a protective effect against oxidative stress in low concentrations, 29 whereas it induced oxidative stress in laboratory animals at high concentrations. 30
Programmed cell death, or apoptosis, is a physiological process of autodestruction. Apoptosis plays critical roles in development, maintenance of homeostasis, and host defense in multicellular organisms. During apoptosis, intracellular contents are not released, and potentially harmful inflammatory responses are prevented. Apoptotic cell death is characterized by fragmentation of DNA into 180–200 pb nucleosomal units. The detection of DNA fragmentation in HL-60 leukemia cells exposed to different concentrations of apigenin, bisabolol, and protocatechuic acid indicated that the cells exposed to these concentrations suffer cell death via apoptosis, and therefore their activity could be catalogued like an antiproliferative action. Compounds that promote apoptosis should become an important addition to the arsenal of target-specific drugs in the fight against cancer.
Conclusions
Our results confirmed the antigenotoxic and chemopreventive potentials of the three phenols evaluated using SMART with D. melanogaster. The leukemic cell line HL-60 provides an important model for studying the mechanisms and relationships among cytotoxicity, apoptosis, and antitumor efficacy of new drugs. Protection of genetic material to avoid oncogenic mutations and selective inhibition of tumoral cell growth are two important pathways for cancer prevention. Apigenin, bisabolol, and protocatechuic acid showed promising results and can play beneficial roles in human nutrition and health status.
Footnotes
Author Disclosure Statement
No competing financial interests exist.