
Abstract
To elucidate whether a low methionine (Met)/glycine (Gly) ratio is responsible for the hypocholesterolemic effect of rice protein (RP), the effects of adding Met to RP, increasing its Met/Gly ratio, on hepatic cholesterol output was investigated in rats fed cholesterol-free diets. The hepatic secretion of cholesterol into bile or circulation was measured from isolated perfused livers of 7-week-old male Wistar rats fed RP (cultivar Koshihikari) with a lower Met/Gly ratio and methionine-supplemented RP with a higher Met/Gly ratio (RP-M) matched with casein (CAS). RP-M produced a significant hypocholesterolemic effect, whereas the effect of RP on plasma cholesterol level was comparable to that of CAS. Hepatic accumulation of total lipids, cholesterol, and phospholipids was higher in RP-M rats than in RP rats, again following a pattern similar to that in CAS rats. The hepatic total and very-low-density lipoprotein (VLDL)-cholesterol secretions into circulation were effectively decreased by RP, but not by RP-M. Bile flow and biliary outputs of bile acids, cholesterol, and phospholipids were significantly stimulated by RP-M, causing an increase in fecal sterol excretion compared with CAS and RP. Neither biliary output nor fecal excretion of bile acids was affected by RP compared with CAS. The present results demonstrate that the plasma cholesterol-lowering effect of RP cannot be merely ascribed to the low Met/Gly ratio of RP in growing rats fed cholesterol-free diets. Results suggest that the hypocholesterolemic response induced by RP-M with a higher Met/Gly ratio primarily contributes to the stimulation of hepatic cholesterol for elimination via biliary secretion rather than the inhibition of hepatic cholesterol release via VLDL into circulation.
Introduction
R
Met is mainly metabolized in the liver and is suggested to elicit atherogenic effects. 7,8 Several reports have shown that Met can stimulate the formation of phosphatidylcholine, which is the major phospholipid (PL) of plasma lipoproteins and is required for the assembly and secretion of very-low-density lipoprotein (VLDL) from liver into the plasma. 9 –11 In contrast, Gly suppresses phosphatidylcholine synthesis through consumption of methyl groups from S-adenosylmethionine, leading to a decrease in the secretion of lipoproteins from the liver. 12 In view of these facts, it is possible that the ratio of Met to Gly regulates the assembly and secretion of VLDL from the liver.
The liver plays a central role in maintaining cholesterol homeostasis through regulatory hepatic outputs of cholesterol into either bile or plasma. 13,14 Hepatic cholesterol secretion into plasma occurs principally through the formation and secretion of VLDL, 15,16 whereas the biliary pathway represents the major route for removal of hepatic cholesterol from the body through both the direct secretion of cholesterol itself and the conversion of cholesterol into primary bile acids. 17,18 Recent progress has demonstrated that protein and lipid factors can affect hepatic VLDL assembly and secretion. 19 In particular, several studies have led to a better understanding of these regulatory pathways for cholesterol output involved in the hypocholesterolemic response to dietary proteins. 20,21 Sugano et al. 20 showed that the cholesterol-lowering effect of soy protein in comparison with CAS was attributable to decreased hepatic secretion of cholesterol into blood circulation. Furthermore, our ex vivo studies indicated that the stimulation of biliary production and the inhibition of hepatic cholesterol secretion into VLDLs are responsible for the hypocholesterolemic action of RPs in growing rats fed cholesterol-enriched diets. 21 Taken together, the evidence suggests that attenuated hepatic cholesterol output might be the mechanism of the hypocholesterolemic action of RP.
Thus, the present study was conducted to obtain insight into the influence of the Met/Gly ratio of RP on hepatic cholesterol output and its effect on cholesterol metabolism in growing rats on cholesterol-free diets. The key questions addressed were as follows: (1) Does the alteration of Met/Gly ratio of RP affect plasma cholesterol concentration, and (2) how does the Met/Gly ratio of RP regulate hepatic cholesterol output and cholesterol metabolism in general? In addition, this study also focused on modulation of hepatic secretion of PLs through alteration of the Met/Gly ratio.
Materials And Methods
Protein sources
RP from Oryza sativa L. Japonica cv. Koshihikari and CAS were used as the dietary protein sources in the present study. RP was prepared by the alkaline extraction method (Shimada Inc., Nagaoka, Japan), and CAS was obtained from Fontera (Kanagawa, Japan). Amino acid analysis of these proteins was performed using a Hitachi (Tokyo, Japan) model L-880 amino acid analyzer according to the method of Yang et al. 6 The ratios of Met/Gly in RP and CAS were 0.67 and 1.65, respectively (Table 1).
Data are averages of triplicate analyses.
CAS, casein; Gly, glycine; Met, methionine; RP, rice protein (Koshihikari).
Animals and diets
The present experiments were performed in conformity with the Guidelines of the Committee for Animal Experimentation of Niigata University. 6 Seven-week-old male Wistar rats were purchased from Japan SLC (Hamamatsu, Japan) and used for in vivo and ex vivo perfusion studies. All animals were individually housed in metabolic cages in a room maintained at 22 ± 2°C under a 12-hour light–dark cycle (06:00–18:00 hours for light). Rats were allowed free access to commercial pellets (Nosan Corp., Yokohama, Japan) for 3 days. After acclimatization, they were randomly divided into three groups of similar body weight. Each group consisted of 12 animals (six rats for in vivo studies and six rats for ex vivo perfusion studies, respectively).
All animals were fed modified AIN-93 diets ad libitum. 22 For 2 weeks, rats were fed cholesterol-free diets with 20% dietary protein level (as crude protein). CAS, RP, and Met-supplemented RP (RP-M) (Met/Gly ratio adjusted to 1.65 to be equivalent to CAS) were used as dietary proteins. Diets were completed to 100% with starch. The composition of experimental diets is shown in Table 2.
Crude protein level: CAS, 88.0%; RP, 81.3%.
Mineral Mix, AIN-93G-MX (Nosan Corp.).
Vitamin Mix, AIN-93-VX (Nosan Corp.).
RP-M, Met-supplemented RP having a Met/Gly ratio with an equivalent level of CAS.
Ex vivo liver perfusion
The liver perfusion was performed according to the method of Yang and Kadowaki. 21 The rats were anesthetized with sodium pentobarbital (50 mg/kg of body weight) at from 10:00 to 11:00 hours. The bile duct was cannulated with a PE-10 polyethylene tube, and bile was collected in the preweighed 1.5-mL tube. The portal vein and vena cava were then ligated before the portal vein was cannulated, while the bile duct was avoided, and the exit cannula was inserted into the superior vena cava. During the perfusion period, the liver was continuously perfused with 210 mL of recirculating Krebs–Ringer bicarbonate medium containing 0.18% (wt/vol) glucose, 3% (wt/vol) bovine serum albumin (Sigma, St. Louis, MO, USA), and 25% (vol/vol) washed hemoglobin-free erythrocytes and was oxygenated with 95% O2 and 5% CO2, at the rate of 15 mL/minute at 37°C. The perfusate was continuously oxygenated with 95% O2 and 5% CO2 using the apparatus described by Yang and Kadowaki. 21 The perfusion was maintained for 4 hours. At 1-hour intervals, bile and 20 mL of perfusate were withdrawn for analysis, while the same volume of fresh perfusion medium was added at the time of each removal. Samples of perfusate were centrifuged at 12,000 g for 5 minutes to sediment erythrocytes, and the supernatant was used for analysis.
Sampling for in vivo studies
During the feeding period, food consumption and body weight were recorded daily in the morning before replenishing the diet. Feces were collected for the final 3 days of the experimental period, dried to a constant weight, and ground to a fine powder for fecal steroids determination according to the method of Yang et al. 6 After fasting for about 18 hours, the rats were sacrificed at from 10:00 to 11:00 hours. Blood was withdrawn from the abdominal vein into a heparinized syringe with the animal under anesthesia with sodium pentobarbital (50 mg/kg of body weight), immediately cooled on ice, and separated by centrifugation at 12,000 g for 5 minutes. The plasma obtained was frozen at −20°C until analysis. After blood collection, the liver was excised immediately, rinsed in saline, and weighed after blotting on a filter paper. The whole liver was cut into three portions, quickly freeze-clamped in liquid nitrogen, and stored at −80°C until analysis.
Isolation of perfusate lipoproteins
VLDL (d < 1.006 g/mL fractions) in the plasma and the erythrocyte-free perfusate were isolated by the ultracentrifugation method of Hatch and Lees. 23 Samples of plasma and erythrocyte-free perfusate (1.6 mL) were added to ultracentrifuge tubes, and 0.8 mL of d < 1.006 g/mL solution was layered over the surface. The tubes were capped and centrifuged in S 100 AT 4-557 rotors (Hitachi) in a Hitachi CS 100GXL ultracentrifuge for 16 hours at 50,000 rpm (114,000 g) at a chamber temperature of 16°C. The 0.8-mL layer at the top of the centrifuge tube contained the VLDL fraction and was recovered quantitatively for analysis.
Plasma lipid analysis
Plasma concentrations of total cholesterol (TC), VLDL cholesterol (VLDL-C), high-density lipoprotein cholesterol (HDL-C), and PL were measured colorimetrically with commercial kits (Cholesterol E-test Wako, HDL-Cholesterol E-test Wako, and Phospholipid E-test Wako, Wako Pure Chemicals, Osaka, Japan). Plasma low-density lipoprotein cholesterol (LDL-C) was calculated as: LDL-C = TC – HDL-C – VLDL-C.
Liver lipid analysis
Liver lipids were extracted and purified according to the method of Folch et al. 24 and were analyzed as described by Yang et al. 6 Samples of liver were extracted with chloroform/methanol (2:1, vol/vol). TC, free cholesterol, and PLs were measured with a commercial kit (cholesterols, Roche Diagnostics, Tokyo; Phospholipid E-test Wako, Wako Pure Chemicals). Total lipids were determined gravimetrically. The concentration of esterified cholesterol was calculated as: esterified cholesterol = TC – free cholesterol.
Determination of fecal excretion
Fecal bile acid and cholesterol concentrations were extracted and measured as previously described. 6 Total bile acids were determined enzymatically by 3α-hydroxysteroid dehydrogenase (Sigma) with sodium taurocholate as the standard, 21 and fecal cholesterol was measured with a commercial kit (Cholesterol E-test Wako, Wako Pure Chemicals).
Estimation of biliary secretion
Bile flow was determined gravimetrically, assuming a specific gravity of 1.0 for bile. Biliary bile acids were measured enzymatically by 3α-hydroxysteroid dehydrogenase using sodium taurocholate as the standard. 21 Biliary cholesterol and PLs were determined colorimetrically with commercial kits (Cholesterol E-test Wako and Phospholipid E-test Wako, respectively, Wako Pure Chemicals).
Estimation of hepatic secretion of lipids
The lipids in the erythrocyte-free perfusate and VLDL fractions were extracted and purified according to the method of Folch et al. 24 and analyzed as described by Yang and Kadowski. 21 Cholesterol and PLs in the perfusate were measured colorimetrically with commercial kits (Cholesterol E-test Wako and Phospholipid E-test Wako, respectively, Wako Pure Chemicals).
Statistical analysis
Data are presented as mean ± SEM values. Differences among groups were examined for statistical significance by one-way analysis of variance and separated by the least significant difference test. The criterion for significance was P < .05.
Results
Effects on food intake and body weight
Food intake, final body weight, and body weight gain were significantly lower in the RP and RP-M groups than in the CAS group. No differences were found in rats fed RP and RP-M, suggesting that the dietary Met/Gly ratio in RP did not affect food intake or growth performance in growing rats (Table 3).
Data are mean ± SEM values (n = 6).
Values with different superscript letters are significantly different (P < .05).
Effects on plasma lipids and lipoprotein profiles
Compared with CAS, plasma cholesterol and PL concentrations were significantly lower in growing rats fed RP-M with the higher Met/Gly ratio (TC, −24.16%; PL, −15.72%), whereas the hyperlipidemic action induced by RP with a lower Met/Gly ratio was similar to that of CAS (Table 4), in accordance with our previous results. 6 With the increasing ratio of Met/Gly, plasma concentrations of cholesterol and PL were significantly reduced by RP-M (TC, −23.13%; PL, −14.10%), compared with RP.
Data are mean ± SEM values (n = 6).
Values with different superscript letters are significantly different (P < .05).
HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; TC, total cholesterol; VLDL-C, very-low-density lipoprotein cholesterol.
Plasma HDL-C level was not significantly among the different dietary groups. However, plasma VLDL-C and LDL-C concentrations distinctly fell by −15.38% and −65.31% with RP-M, respectively, compared with CAS, resulting in a distinct decrease in the non-HDL-C/HDL-C ratio by −53.52% in RP-M. The cholesterol distribution in the RP group was similar to that of the CAS group, with similar concentrations of VLDL-C and VLDL-C/HDL-C. As a result, the non-HDL-C concentration was also significantly lowered by −50.88% in RP-M in comparison with RP.
Effects on liver lipids concentration
There was no significant difference in liver weight between the RP and RP-M groups, but both were significantly lower than CAS (Table 5).
Data are mean ± SEM values (n = 6).
Values with different superscript letters are significantly different (P < .05).
Hepatic accumulations of total lipids, TC, and PL were significantly lower in the RP group than in the CAS group, in accordance with our previous results. 6 The hepatic cholesterol-lowering action induced by RP feeding was mainly reflected by diminished concentrations of free and esterified cholesterol.
With the higher Met/Gly ratio, RP-M had significantly higher hepatic total lipids, free and esterified cholesterol, and PL compared with RP, with hepatic lipid accumulation similar to CAS.
Effects on fecal sterol excretion
The dietary Met/Gly ratio did not affect fecal outputs. The 3-day fecal output tended to be similar in all experimental groups, as shown in Table 6.
Data are mean ± SEM values (n = 6).
Values with different superscript letters are significantly different (P < .05).
Fecal total bile acid concentrations and daily fecal bile acid excretion were enhanced with the higher ratio of RP Met/Gly in growing rats fed cholesterol-free diets, showing higher fecal excretion of bile acids in the RP-M group than in the RP group. Compared with CAS, RP-M significantly increased daily excretion of fecal bile acids by 38.00%, whereas fecal outputs of bile acids were similar in the RP and CAS groups.
Fecal cholesterol concentrations, as well as daily excretion of fecal cholesterol, were also significantly stimulated by RP-M feeding, whereas fecal cholesterol excretion in RP-fed rats was similar to that in the CAS group. The results also suggest that fecal cholesterol excretion in rats was effectively enhanced with the increasing dietary ratio of RP Met/Gly (Table 6).
Effects on bile flow and secretion of biliary lipids
The rate of bile flow and biliary output of bile acids, cholesterol, and PLs by the perfused rat liver remained constant during the entire 4-hour period.
As shown in Figure 1, bile flow and biliary outputs of bile acids, cholesterol, and PLs in growing rats fed RP were comparable to those fed CAS. In contrast, biliary secretion of bile acids, cholesterol, and PLs were significantly increased about 33.66%, 31.61%, and 44.70% by RP-M, respectively, compared with CAS.
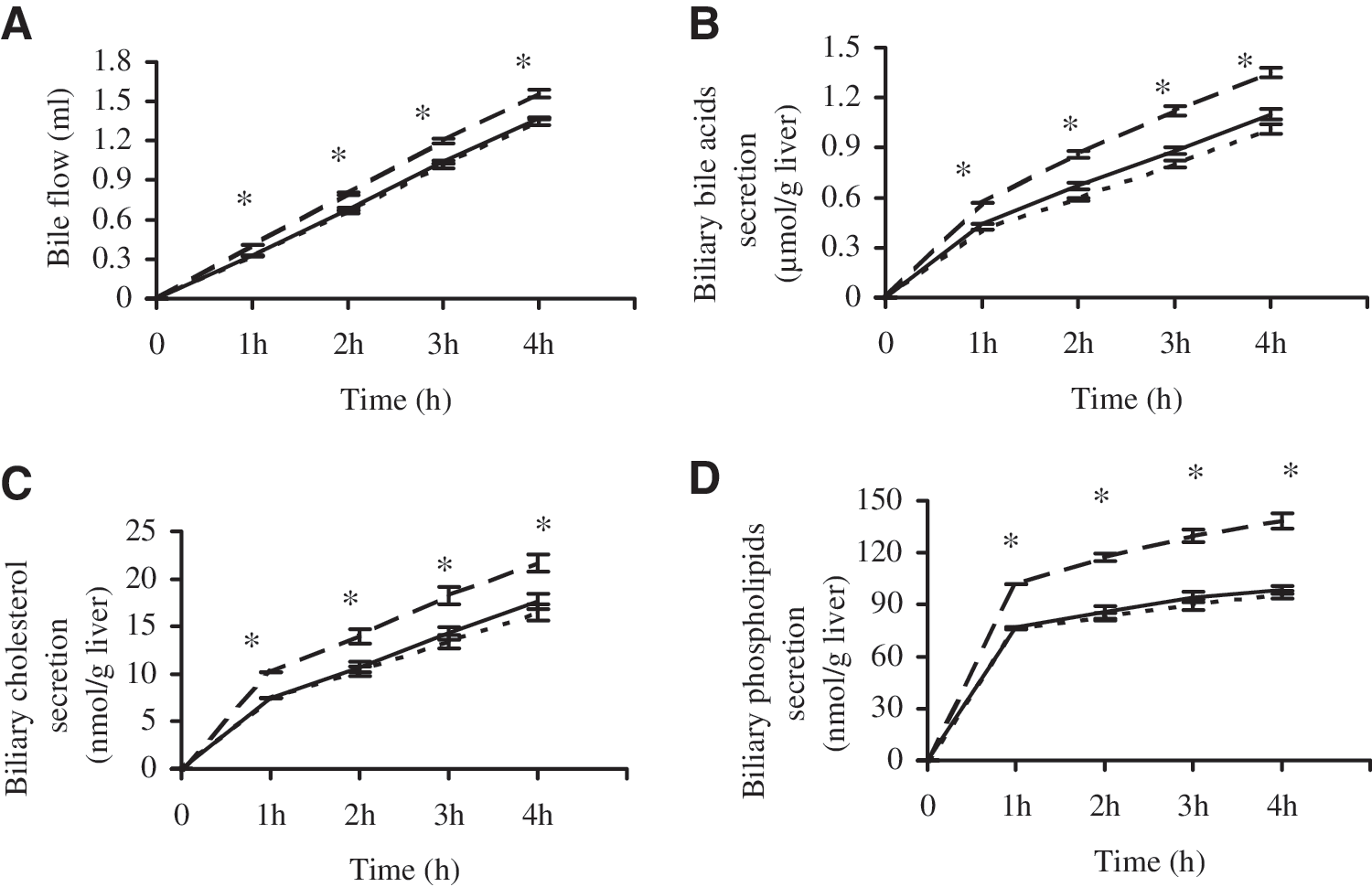
Effects of dietary proteins on bile flow and biliary secretion of bile acids, cholesterol, and phospholipids. Livers from 7-week-old male Wistar rats fed cholesterol-free diets with 20% dietary proteins (CAS [---], RP [—], or RP-M [– –]) for 2 weeks were continuously perfused with 210 mL of recirculating Krebs–Ringer bicarbonate medium containing 0.18% (wt/vol) glucose, 3% (wt/vol) bovine serum albumin, and 25% (vol/vol) washed hemoglobin-free erythrocytes for 4 hours. At 1-hour intervals, bile was drawn for analysis of (
The increased Met/Gly ratio by addition of Met to RP (RP-M) effectively stimulated bile flow (Fig. 1A). As a result, a marked enhancement of output of biliary lipids was observed throughout the 4-hour perfusion period in the RP-M group compared with the RP group, demonstrating that RP-M significantly increased biliary secretion of bile acids (Fig. 1B), cholesterol (Fig. 1C), and PLs (Fig. 1D) by 22.73%, 22.89%, and 40.53%, respectively, suggesting that increasing the Met/Gly ratio of RP increases biliary secretion in growing rats fed cholesterol-free diets.
Effects on secretion of hepatic lipids
As summarized in Figure 2, increasing the Met/Gly ratio of RP effectively affected secretion of hepatic lipids into circulation in growing rats fed cholesterol-free diets.
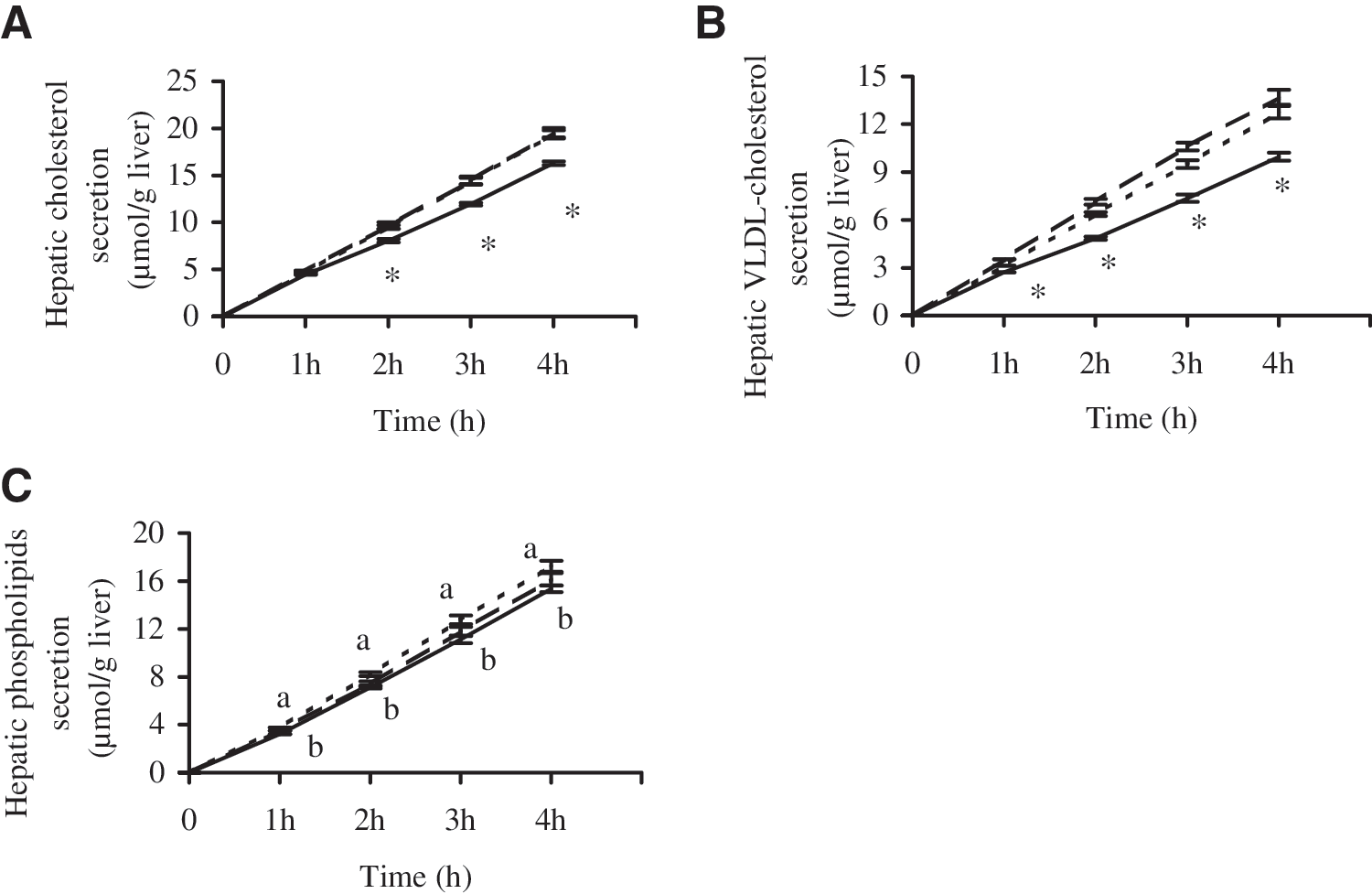
Effects of dietary proteins on hepatic secretion of cholesterol [(
Compared with CAS, total hepatic cholesterol secretion and hepatic cholesterol secretion into VLDL particles were significantly decreased in the isolated perfused liver in rats fed RP throughout the 4-hour perfusion (Fig. 2A and B). Observations in this study indicated that the suppression of hepatic TC and VLDL-C secretion induced by RP feeding was efficaciously abolished by addition of Met in RP-M. Compared with RP, RP-M significantly enhanced hepatic TC and VLDL-C secretion by 19.94% and 37.55%, respectively. As a result, the ratio of VLDL-C/TC in hepatic secretion was enhanced by 14.70% in RP-M relative to RP. Similar export of hepatic TC and VLDL-C from control livers of rats fed RP-M and CAS was found throughout the perfusion period (Fig. 2A and B).
Accompanying the increase export of hepatic cholesterol throughout the perfusion period, RP-M also increased PL secretion from liver relative to RP, but there was no significant difference between the two groups. Compared with CAS, RP significantly decreased hepatic PL secretion (Fig. 2C), whereas the hepatic output of PLs into circulation induced by RP-M did not differ from that in rats fed CAS (Fig. 2C).
Discussion
We examined the regulatory effect of RP with a lower or higher ratio of Met/Gly on cholesterol metabolism and especially its effect on hepatic outputs of cholesterol in growing rats fed cholesterol-free diets. In agreement with our previous observations, 6 RP with a lower ratio of Met/Gly could not exert a hypocholesterolemic effect in growing rats fed cholesterol-free diets, showing that the effect on plasma cholesterol level induced by CAS and RP was comparable. Unexpectedly, we observed a more pronounced lowering of plasma cholesterol in rats consuming RP-M with a higher Met/Gly ratio. RP-M intake resulted in 24.16% lower total cholesterol and 54.84% lower non-HDL-C compared with CAS with an equal Met/Gly ratio. These observations were inconsistent with previous reports showed by Morita et al. 5 and did not support our original hypothesis that an increased ratio of Met/Gly in RP could result in higher plasma cholesterol levels. Similar findings were reported by Kern et al., 25 who showed that supplementing a soy diet with Met did not abolish the hypocholesterolemic effects of soy protein relative to CAS, suggesting that Met might not be involved in the effects of soy protein on lipid metabolism. Furthermore, results of Moundras et al. 26 indicated that Met deficiency in rats fed soy protein induces hypercholesterolemia. In view of these facts, therefore, we can speculate that the dietary Met/Gly ratio of RP might not be the major contributor to plasma cholesterol levels in growing rats fed with cholesterol-free diets.
An important metabolic alteration that takes place in liver after RP-M treatment—increased accumulation of hepatic cholesterol—followed a pattern similar to that of CAS. This finding, consistent with previous studies, 27 suggests that a compensatory increase in hepatic cholesterol synthesis resulting from increased 3-hydroxy-3-methylglutaryl-coenzyme A reductase expression might result from Met supplementation in RP.
As a result of expanded hepatic cholesterol pools due to higher Met/Gly ratio intake, lipoprotein metabolism was altered. Some studies have demonstrated that increases in hepatic cholesterol are related to higher rates of hepatic apolipoprotein B secretion, which is important for assembly and secretion of VLDL from the liver. 15,19,28 Our results confirmed and expanded this view. The major finding in this study was that hepatic cholesterol secretion into circulation is responsive to the dietary Met/Gly ratio, showing that a higher ratio of Met/Gly in RP can increase the VLDL production and secretion from liver, whereas a lower ratio of Met/Gly in RP can depress hepatic VLDL secretion. In addition, the observed RP-M-induced metabolic alterations in hepatic cholesterol output via VLDL were similar to those of CAS. Accordingly, a significant positive correlation between the secretion of hepatic VLDL-C and daily intake of dietary Met/Gly was observed in this study (r = 0.7788). It is fairly likely, therefore, that the stimulation of hepatic cholesterol output via VLDL into circulation might be closely linked with the increased hepatic cholesterol accumulation resulting from a higher dietary Met/Gly ratio, supporting our hypothesis that the Met/Gly ratio might be an important factor for altering hepatic assembly and secretion of VLDL.
Of interest was the finding that inhibiting VLDL secretion from liver could not cause a hypocholesterolemic action in growing rats fed RP with a lower ratio of Met/Gly. Also, compared with CAS, we did not observe a plasma cholesterol-elevating effect of RP-M, although the observed changes tended to parallel the effects on hepatic VLDL-C secretion in both groups. These observations indicated a lack of association of Met/Gly-altered hepatic cholesterol release into circulation with the plasma cholesterol concentration. Thus, the question arises why the increase outward transport of cholesterol from liver induced by the higher ratio of Met/Gly in RP-M did not lead to increased plasma cholesterol. The precise mechanism by which the Met/Gly ratio affects cholesterol metabolism is not fully understood, but the mechanism responsible for the effect on the removal of VLDL from circulation by hepatic uptake should be taken into account. 29 Our data obtained from plasma HDL-C and TC concentrations indicated that RP-M feeding produced the highest ratio of HDL-C/TC in all experimental groups (RP-M, 0.75; RP, 0.61; CAS, 0.58). Thus, increasing the Met/Gly ratio by adding Met to RP mainly increased the proportion of HDL-C, which normally promotes the transfer of peripheral free cholesterol to the liver by a mechanism known as “reverse cholesterol transfer.” 30 –32 In support of this, previous findings have established the critical role of hepatic scavenger receptor class B type I expression in selectively controlling the utilization of HDL-C for biliary secretion. 33 Therefore, the higher hepatic accumulation of cholesterol for biliary secretion observed in this study with an RP-M diet might bring about an increased uptake of lipoprotein cholesterol by the liver, leading to an inhibition of cholesterol accumulation into plasma. Thus, the stimulation of the uptake of lipoprotein cholesterol by the liver may participate in the hypocholesterolemic action of a high Met/Gly ratio in RP, where high-density lipoprotein is the major plasma lipoprotein.
Additionally, it must be noted that Met appears to have two effects, namely, the plasma cholesterol-elevating effect due to its methyl group and the plasma cholesterol-lowering effect due to its non-methyl portion. 34 Consequently, the final plasma cholesterol concentration induced by Met is a consequence of the “hypercholesterolemic” effect of the labile methyl groups and the “hypocholesterolemic” effect of the greater intake of the non-methyl portion. Furthermore, it has been suggested that Met is more effective in reducing plasma cholesterol concentration when the supply of Gly is sufficient in diets. 35 In light of this view, addition of Met to RP, which is a rich source of Gly, results in a higher Met/Gly ratio, causing a stronger cholesterol-lowering effect, contributing to the overcoming of its plasma cholesterol-elevating effect. These findings further provide a favorable explanation for the observation why the same ratio of Met/Gly in RP-M and CAS caused different plasma cholesterol levels in this study. Clearly, the findings observed in this study suggest that the hypocholesterolemic response to RP cannot be merely ascribed to the depressed secretion of hepatic cholesterol via VLDL into plasma, which resulted from a lower ratio of Met/Gly; other mechanisms may come into play to fully explain the hypocholesterolemic action of RP.
Conversion of cholesterol to bile acids is the principal regulatory pathway whereby cholesterol is eliminated from the body, primarily via the hepatobiliary system. 13,17,18 Thus, whatever the mechanism of the hypocholesterolemic response to RP, the effect on hepatic cholesterol output into bile is important. Consistent with this view, the lack of an apparent increase in fecal excretion of bile acids rats fed RP, causing a failure in hypocholesterolemic action, might be explained by the fact that biliary secretion was not enhanced by RP. Furthermore, data obtained here showed that plasma cholesterol concentration in growing rats fed cholesterol-free diets was negatively correlated with secretion of biliary bile acids (r = –0.7774) or excretion of fecal sterols (r = –0.8280). The present study, therefore, makes clear that the increase in the availability of hepatic cholesterol for elimination from the body via biliary secretion seems to be the principal mechanism of the hypocholesterolemic action of RP-M. It would appear that a link between the stimulation of excretion of bile acids and a higher Met/Gly ratio in RP in this study is conceivable. However, this linkage is not sufficiently appreciated in CAS feeding. In fact, the different extent to which RP-M and CAS caused secretion of hepatic bile acids as well as fecal excretion was observed in this study, showing the increment of bile acid removal was more than 37.79% greater with RP-M than with CAS, despite the same ratio of Met/Gly in both groups. This apparent discrepancy is not fully understood. Possibly, the same Met/Gly ratio with different hepatic cholesterol output into bile may have an effect in that (1) larger amounts of Gly in RP-M may efficaciously stimulate the conversion of cholesterol into bile acids related to CAS, because Gly is required in disproportionately large amounts for the formation of bile salts, 36 or (2) cholesterol needed as a precursor for bile acid synthesis with RP-M seems to be generated from lipoprotein cholesterol via up-regulated receptor uptake rather than forming newly synthesized cholesterol. Clearly, additional studies are required to confirm these views.
Indeed, identifying which of the components of RP is the major contributor to effects on lipid metabolism is still a subject of controversy. The current studies clearly showed that several mechanisms for the cholesterol-lowering effect of RP probably exist. Thus, besides the Met/Gly ratio, other factors in RP possibly impacting cholesterol homeostasis (e.g., other aspects of the amino acid profile, bioactive peptides, etc.) should be taken into account and remain to be elucidated in future studies.
It should be also emphasized that alterations in the Met/Gly ratio of RP were associated with changes in the metabolism of liver PLs. The observed changes induced by RP-M tended to parallel the effects of CAS on hepatic accumulation and hepatic secretion of PLs into circulation, consistent with the view that a high dietary Met level might stimulate metabolism of hepatic PLs. 37 However, results observed in this study indicated that RP-M with a higher Met/Gly ratio significantly promoted enhanced biliary secretion of PLs, compared with CAS or RP. This might be a result of the stimulation of PLs on the transport of unesterified cholesterol from HDL-C to the liver (“reverse cholesterol transfer”) because PLs are key molecules controlling the biodynamic exchanges of lipids and lipoproteins. 17,32
In conclusion, in the absence of dietary cholesterol, a lower ratio of Met/Gly, leading to a decrease in hepatic VLDL secretion, might not be the main modulator responsible for the cholesterol-lowering action of RP in growing rats. The hypocholesterolemic response induced by a higher ratio of Met/Gly in RP primarily contributes to the stimulation of hepatic cholesterol for elimination via biliary secretion and the increased capability of lipoprotein clearance by hepatic uptake. Our results demonstrate that alteration of the Met/Gly ratio effectively regulates cholesterol metabolism in the liver, especially hepatic cholesterol outputs. The relevance of the dietary Met/Gly ratio with regard to cholesterol metabolism and the precise mechanisms involved in the hypocholesterolemic responses to RP await more detailed investigation.
Footnotes
Acknowledgments
We are grateful to the Laboratory of Nutritional Regulation of Niigata University for their constant support. This work was funded in part by the National Natural Science Foundation of China (number 31071526), by the China Postdoctoral Science Foundation (numbers 200902391 and 20080440864), and by the Technology and Technical Development Program for agribusiness in the form of utilizing concentrated know-how from the private sector in Japan.
Author Disclosure Statement
No competing financial interests exist.