
Abstract
Mushrooms and mushroom extracts have traditionally been used as therapies for a wide variety of ailments, including allergy, arthritis, and other inflammatory disorders. However, more evidence is required on the mechanism by which mushrooms exert these effects. In the present study, the anti-inflammatory properties of ethanol and hot water extracts prepared from 27 fungal samples collected between October and November 2011 at various forest locations in the southwest of Ireland were investigated using the lipopolysaccharide (LPS)–stimulated mouse macrophage (RAW264.7 cells) model of inflammation. LPS-stimulated cells were incubated in the presence of mushroom extracts at nontoxic concentrations for 24 h and the production of interleukin-6 (IL-6) was quantified by ELISA. Seven ethanolic and one hot water extract that decreased IL-6 production were selected for further study. The extracts were then incubated with LPS-stimulated cells for 24 h and the production of IL-6, tumor necrosis factor-alpha (TNF-α), and nitric oxide (NO) was measured. Ethanolic extracts prepared from Russula mairei, Lactarius blennius, Craterellus tubaeformis, Russula fellea, and Craterellus cornucopioides demonstrated selective anti-inflammatory activity by decreasing the production of NO and IL-6 but not TNF-α in LPS-stimulated RAW264.7 cells. These findings support existing evidence of the anti-inflammatory potential of mushroom extracts.
Introduction
T
Chronic inflammation, which is characterized by an excess of circulating proinflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6), is a causative factor in the development of heart disease, rheumatoid arthritis, diabetes, and cancer. 11 Wild mushrooms and their extracts have been shown to possess both proinflammatory and anti-inflammatory activities depending on the mushroom species, the method used in the preparation of the extracts, and the experimental model selected for study. 12 Both edible and medicinal mushrooms and mushroom extracts, including a glycoprotein extracted from Pleurotus citrinopileatus, a triterpene isolated from Ganoderma lucidum, and crude extracts prepared from either the fruiting body or mycelia of Antrodia cinnamomea, Phellinus baumii, Pleurotus ostreatus, or Lentinus polychrous, have demonstrated anti-inflammatory effects in the lipopolysaccharide (LPS)–stimulated RAW264.7 mouse macrophage model. 13 –18
While the evidence for their anti-inflammatory activity is building, to date only a relatively small number of wild species have been investigated and no studies have been carried out on species collected in Ireland. Therefore, the objective of the present study was to determine whether crude extracts prepared from wild mushrooms harvested in Ireland demonstrate anti-inflammatory effects in RAW264.7 cells by measuring the production of IL-6, TNF-α, and nitric oxide (NO) following exposure of the cells to LPS in the presence of the mushroom extracts.
Materials and Methods
Materials
RAW264.7 cells were purchased from the American Type Culture Collection (ATCC; LGC Standards, Middlesex, United Kingdom). Cell culture plastics were purchased from Cruinn Diagnostics (Dublin, Ireland). Fetal bovine serum (FBS) was purchased from Invitrogen (Paisley, Scotland). All other chemicals and reagents were purchased from Sigma Chemical Co. (Dublin, Ireland) unless otherwise stated.
Sample collection
The 27 fungal samples investigated in this study, which included both edible and nonedible species, were collected between October and November 2011 from various forest locations around the southwest of Ireland, including Curragh Chase Forest Park, Cratloe Forest Park, and Killarney National Park.
Each mushroom specimen was collected and identified by a trained mycologist. These included Russala nigricans, Clitocybe nebularis, Lactarius deterrimus, Paxilllus involutus, Cortinarius croceus, Russula mairei, Hydnum repandum, Lactarius blennius, Craterellus tubaeformis, Leotia lubrica, Lycoperdon perlatum, Cortinarius sect. cinnamomeobadei, Russula fellea, Craterellus cornucopioides, Suillus luteus, Hebeloma crustuliniforme, Trichloma ustale, Collybia confluens, Amanita citrina, Agarius semotus, Inocybe flocculosa, Russula ochroleuca, Amanita rubescens, Helvella crispa, Thelophora terrestris, Laccaria amethystina, and Mycena pura. Each of the mushroom samples was cleaned with a soft brush to remove soil debris and frozen. All samples were freeze dried (A12/60 Freeze Dryer, Frozen In Time Ltd., York, United Kingdom) and ground to a fine powder and stored in vacuum-packed bags at −80°C prior to extraction.
Extraction
Solid-liquid extraction was employed to extract the freeze-dried mushroom powders with two different solvents, ethanol (room temperature) and hot water (60°C), in a sequential manner. Ethanol and hot water have previously been used by Hu et al. 19 to prepare bioactive extracts from mushrooms. For the ethanol extracts, each sample of dried mushroom powder was mixed in ratio of 1:7.9 w/w with ethanol and for the hot water extractions (carried out on the material remaining after ethanol extraction) this ratio was changed to 1:20 w/w due to the viscosity of the material extracted. Extractions were performed in an orbital shaker (MaxQ 6000 Shaker, Thermo Fisher Scientific, Westmeath, Ireland) at 175 rpm. Following 3 h of incubation, ethanol extracts were filtered through a buchner funnel and the material was re-extracted with fresh solvent for a further 3 h. The process was repeated with a final overnight extraction to ensure exhaustive extraction occurred and the solvents were combined. The residue remaining for each mushroom sample was dried using nitrogen and then used for the hot water extract. The hot water extraction was filtered initially after 6 h and then after 24 h through glass wool filters and combined. Ethanol was removed from the ethanol extractions using a rotary evaporator (Heidolph Rotary Evaporator with WB eco bath, Schwabach, Germany) with the water bath set at 50°C. The hot water extracts were frozen at −20°C and then freeze dried (A12/60 Freeze Dryer, Frozen In Time Ltd., York, United Kingdom).
The stock solutions of the wild mushrooms (10 mg/mL), prepared with 10% DMSO in water, were diluted to 100 μg/mL in Dulbecco's modified Eagle's medium (DMEM) and filter sterilized using 0.22 μm Millex GP, PES membrane filter units (Millipore, Cork, Ireland) before addition to the cells in culture.
Cell culture
RAW264.7 cells were maintained in DMEM, supplemented with 10% FBS, and incubated at 37°C and 5% CO2 in a humidified atmosphere. For experiments, the serum concentration was reduced to 2.5% FBS.
Cell viability
RAW264.7 cells were seeded in 96-well plates at a density of 0.2×104 cells/well and allowed to adhere overnight. In an initial experiment, the cytotoxicity of each of the extracts prepared from the 27 mushroom species was investigated at concentrations ranging from 2.5 to 75 μg/mL and noncytotoxic concentrations (Table 1) were selected for screening the effect of the extracts on IL-6 production in LPS-stimulated RAW264.7 cells. The concentration of extract at which cell viability exceeded 80% was considered as noncytotoxic as previously reported by López-García et al. 20 In the second series of experiments, the eight selected wild mushroom extracts were added to the cells at concentrations ranging from 0 to 50 μg/mL and incubated for 24 h.
Data are expressed as a percentage of the control (100%) LPS-stimulated cells, for three independent experiments±standard error.
Denotes significant difference from control cells (P<.05), ANOVA followed by Dunnett's.
IL, interleukin; LPS, lipopolysaccharide; ND, not determined.
Cell viability was determined using the MTT [3-(4,5-dimethylthiazol-2-yl)-2,5 diphenyltetrazolium bromide] assay (MTT I proliferation kit, Roche Diagnostics, West Sussex, United Kingdom) as previously described. 21 Cell viability was quantified by measuring the absorbance at 570 nm (Varioskan Flash, ThermScientific, Tewksbury, MA, USA) and was expressed as a percentage of control, untreated cells.
Cytokine production
RAW264.7 cells were seeded in 96-well plates at a density of 0.2×104 cells/well and allowed to adhere overnight. In the initial screen of the 27 mushroom species, cells were stimulated to produce IL-6 by the addition of 0.2 μg/mL LPS in the presence of the mushroom extracts at noncytotoxic concentrations (outlined in Table 1) and samples were incubated for 24 h. In the second series of experiments, cells were stimulated to produce cytokines by the addition of 0.2 μg/mL LPS (IL-6) or 0.1 μg/mL LPS (TNF-α) in the presence of the eight selected mushroom extracts at concentrations of 5 and 10 μg/mL and samples were incubated for 24 h.
The cell media were harvested and the quantity of IL-6 and TNF-α in the media was measured by ELISA (eBiosciences, Hatfield, United Kingdom) as described previously. 21 Absorbance was determined at 450 nm and corrected at 590 nm (Varioskan Flash; ThermScientific). Results are expressed as a percentage of the control cells that were exposed to LPS alone.
NO production
For the measurement of NO production, RAW264.7 cells were seeded at a density of 8×104 cells/well as detailed in Kenny et al. 21 and incubated for 48 h; cells were incubated with the mushroom extracts at concentrations of 10 and 20 μg/mL in the presence of LPS (0.5 μg/mL) for a further 24 h. The neutral red uptake assay (NRUA) was used to assess cell viability under the conditions used to assess NO production as this assay is more suitable than the MTT for measuring viability at higher cell densities. For the NRUA, media were removed and 200 μL of neutral red dye (40 μg/mL) was added to the wells. Cells were incubated for 3 h to allow uptake of the dye. The cells were washed and burst using 1% glacial acetic acid and the absorbance was measured at 540 nm (Varioskan Flash; ThermScientific).
For the measurement of NO production, 50 μL of media from each well was transferred to a 96-well plate and 50 μL of Greiss reagent (1% sulfanilamide in 5% phosphoric acid: 0.1% N-1-naphtyl-ethylenediamine dichloride in water; 1:1) was added. Samples were incubated for 20 min and the absorbance was then measured at 540 nm (Varioskan Flash; ThermScientific). Results were expressed as a percentage of the control cells that were exposed to LPS alone.
Statistical analysis
Data represent the mean of at least three independent experiments±standard error. Statistical analysis was evaluated by Dunnett's post-test, P<.05 (Graphpad prism 4.0; GraphPad, Inc., San Diego, CA, USA).
Results
The anti-inflammatory activity of the 27 mushroom extracts, extracted using either ethanol or hot water, was initially screened by measuring the production of IL-6 in LPS-stimulated RAW264.7 cells. The concentration of mushroom extract added to the cells varied from 5 to 75 μg/mL (Table 1) dependent on the cytotoxicity of the extracts; selected concentrations were not cytotoxic (i.e., cell viability exceeded 80%; data not shown). In our study, the extracts prepared using ethanol were less toxic than those prepared by hot water extraction and therefore it was possible to add the ethanol extracts at higher concentrations. Six of the mushroom extracts (C. croceus, R. mairei, C. tubaeformis, R. fellea, C. cornucopioides, and T. ustale) prepared by ethanol extraction caused a significant decrease (P<.05) in IL-6 production (Table 1) and were selected for further investigation. The extract of H. crispa prepared using hot water also significantly decreased (P<.05) IL-6 production and the ethanol extract of L. blennius nonsignificantly decreased IL-6 production to 67.6% of the LPS-treated control (Table 1) and both these samples were also included for further investigation.
The viability of RAW264.7 cells exposed to the selected wild mushroom extracts (0–50 μg/mL) was determined following a 24-h incubation (Fig. 1) using the MTT assay. Each of the wild mushroom extracts induced a dose-dependent decrease in the amount of viable cells. H. crispa hot water extract caused a significant decrease (P<.05) in the number of viable cells at the 30 and 50 μg/mL concentrations. The initial cytotoxicity screen found that C. tubaeformis was not cytotoxic (viability >80%) up to a concentration of 75 μg/mL following a 24-h incubation; however, a more thorough investigation found that this extract induced a significant decrease (P<.05) in cell viability at the 50 μg/mL concentration to 55.6% of the untreated control cells (Fig. 1). The cytotoxicity data for the remaining extracts were consistent between the two series of experiments. Concentrations of 5 and 10 μg/mL were selected for investigation of the effect of the extracts on cytokine (IL-6 and TNF-α) production as cell viability exceeded 80% at these concentrations for all of the extracts.
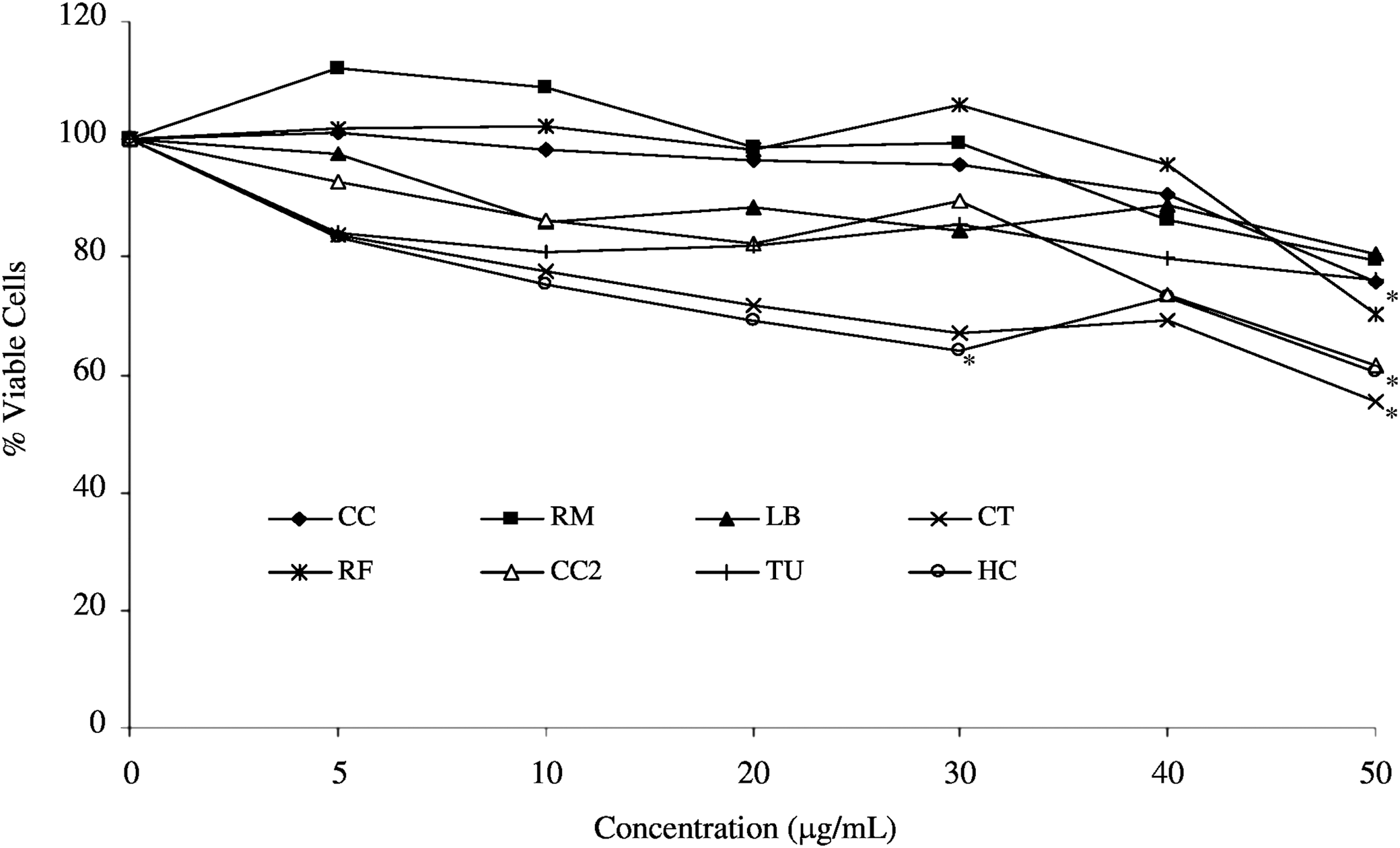
Cell viability, as determined by MTT, in RAW264.7 cells following 24-h incubation with extracts of wild mushrooms. Data are expressed as percentage of untreated control cells and represent three independent experiments. C. croceus (CC), R. mairei (RM), L. blennius (LB), C. tubaeformis (CT), R. fellea (RF), C. cornucopioides (CC2), T. ustale (TU) (all ethanol extracts), and H. crispa (HC) (hot water extract). *Denotes significant difference from control cells (P<.05), ANOVA followed by Dunnett's.
The addition of LPS (0.2 μg/mL) to RAW264.7 cells caused a 25-fold increase in the level of IL-6 produced by the cells, from 2.7 to 68.3 pg/mL (data not shown). At the lower concentration (5 μg/mL) the wild mushroom extracts did not significantly alter the production of IL-6 in LPS-stimulated RAW264.7 cells. However, ethanol extracts from five of the samples (R. mairei, L. blennius, C. tubaeformis, R. fellea, and C. cornucopioides) significantly (P<.05) decreased IL-6 production at the 10 μg/mL concentration, to between 56.4% and 72.1% of the LPS-stimulated control cells (Table 2).
Data are expressed as a percentage of the control LPS-stimulated cells, for three independent experiments±standard error.
Denotes significant difference from control cells (P<.05), ANOVA followed by Dunnett's.
Denotes ethanol extraction.
Corresponds to hot water extraction.
Exposure of RAW264.7 cells to LPS (0.1 μg/mL) resulted in a 15-fold increase in TNF-α production from 12.9 to 191.1 pg/mL (data not shown). The production of TNF-α in LPS-treated cells incubated with the mushroom extracts was not significantly different from TNF-α production in cells exposed to LPS only (Table 2).
Six of the wild mushroom extracts [R. mairei, L. blennius, C. tubaeformis, R. fellea, C. cornucopioides, and T. ustale (all ethanol extracts)] demonstrated a dose-dependent decrease in NO production in LPS-stimulated RAW264.7 cells (Table 3). NO production was significantly reduced (P<.05) to between 44.0% and 66.7% in LPS-stimulated cells in the presence of these six extracts at the 20 μg/mL concentration (Table 3). Cell viability was not significantly altered in cells exposed to LPS and mushroom extracts (10 or 20 μg/mL) in comparison to untreated control cells, as determined by the NRUA (data not shown).
Data are expressed as a percentage of the control LPS-stimulated cells, for three independent experiments±standard error.
Denotes significant difference from control cells (P<.05), ANOVA followed by Dunnett's.
Denotes ethanol extraction.
Corresponds to hot water extraction.
Discussion
LPS interacts with Toll-like receptor-4 (TR4) and activates several pathways involved in the macrophage immune response, including the NF-κB pathway, the MAPK pathway, and the JAK-STAT pathway, and results in the production of IL-6 and TNF-α. A decrease in the production of NO, TNF-α, and IL-6 was previously observed in LPS-stimulated RAW264.7 cells exposed to a triterpene isolated from G. lucidum, 14 a methanolic extract of the mycelia of A. cinnamomea, 17 and a water-soluble lyophilized extract of P. ostreatus. 16
In an initial screen of IL-6 production, each of the extracts was added to the cells at the maximum concentration at which no cytotoxic effects were evident for that extract (Table 1). Eight of the extracts were selected from this screen for further investigation; the concentrations selected for the assessment of cytokine production were 5 and 10 μg/mL as none of the selected extracts demonstrated cytotoxic effects at these concentrations (Fig. 1). In the initial screen (Table 1) C. croceus (ethanol extract) and H. crispa (hot water extract) did induce a significant decrease in IL-6 production at concentrations of 75 and 20 μg/mL, respectively. However, at the lower concentrations (5 and 10 μg/mL) that were not cytotoxic for any of the extracts, C. croceus and H. crispa did not significantly alter IL-6 production. Ethanol extracts prepared from the wild mushrooms (R. mairei, L. blennius, C. tubaeformis, R. fellea, and C. cornucopioides) reduced the production of IL-6 and NO in LPS-stimulated RAW264.7 cells (Tables 2 and 3). T. ustale (ethanol extract) also reduced NO production (P<.05) relative to the LPS-treated control cells but IL-6 production was not significantly altered.
To our knowledge, anti-inflammatory activity has not previously been reported for R. mairei, L. blennius, C. tubaeformis, T ustale, or R. fellea. Three of the mushroom species demonstrating anti-inflammatory activity (R. mairei, R. fellea, and L. blennius) belong to the family Russulaceae and may contain a similar profile of secondary metabolites that could be responsible for their anti-inflammatory effects. Anti-inflammatory activity has been reported in other species of Russula, such as Russula virescens. Hur et al. 22 demonstrated in vitro anti-inflammatory activity of a 70% ethanol extract of Russula virescens in RAW264.7 cell lines at a concentration of 0.5 mg/mL, which is considerably higher than the concentrations found to exhibit significant reduction in NO production in Russula species investigated in this study (R. mairei (10 μg/mL) and R. fellea (10 μg/mL). Moro et al. 4 recently reported that C. cornucopioides along with other wild mushroom species exhibited potential anti-inflammatory activity. The authors speculated that the anti-inflammatory effect of the mushroom extracts including C. cornucopioides may be, at least in part, related to their content of pyrogallol. The present study has demonstrated significant anti-inflammatory activity, in terms of reduced NO production, at concentrations 50-fold lower (10 μg/mL) than that reported in Moro et al. 4
The mushroom extracts investigated in the present study did not alter the production of TNF-α in LPS-stimulated cells. Similarly, Fangkrathok et al. 15 observed that TNF-α was not significantly altered at the mRNA level in LPS-stimulated RAW264.7 cells incubated with 50 and 100 μg/mL extract from the mycelia of L. polychrous; a significant decrease in the expression of IL-6 mRNA, to 80% of the control, was observed under these conditions. Mollugin, a bioactive phytochemical extracted from the Rubia cordifolia plant, was also shown to decrease the production of NO and IL-6 but not TNF-α in LPS-stimulated RAW264.7 cells. 23 Further investigation revealed that mollugin inhibited the JAK-STAT pathway responsible for the induction of IL-6 but not the NF-κB or MAPK pathways that are involved in TNF-α production. Therefore, it is possible that the mushroom extracts (R. mairei, L. blennius, C. tubaeformis, R. fellea, and C. cornucopioides) may operate by a similar mechanism to mollugin with the ability to selectively inhibit NO and IL-6 production. Moro et al. 4 also found that the mushroom species A. bisporus, C. cibarius, and L. deliciosus inhibited NO production and IL-6 expression but did not affect TNF-α expression in LPS-stimulated RAW264.7 cells. Further research is necessary to determine the precise mechanism involved in the anti-inflammatory effects of wild Irish mushroom extracts examined in this study.
Several different solvents, including chloroform, hot water, ethanol, methanol, and hexane, have been used in the preparation of mushroom extracts 24 –27 and the composition of mushroom extracts can vary significantly depending on the method of extraction. Kim et al. 28 found that the β-glucan content of an extract of Hericium erinaceus was three- to fourfold higher in hot water (HWE) and microwaved ethanolic (MWE) extracts than in either acid (ACE) or alkaline extracts (AKE); additionally, there were 40 and 27 identified compounds in HWE and MWE, respectively, in comparison to 16 and 13 in ACE and AKE, respectively. The ACE and AKE extracts did not demonstrate any bioactive effects, indicating the importance of the extraction solvent employed. In our study we found that the ethanolic extracts demonstrated greater anti-inflammatory potential than the mushroom extracts prepared using hot water.
In conclusion, a preliminary screen of extracts from 27 different wild Irish mushroom species demonstrated that seven extracts prepared using ethanol and one hot water extract significantly decreased the production of IL-6 in LPS-stimulated RAW264.7 cells. Further investigation revealed that extracts prepared from R. mairei, L. blennius, C. tubaeformis, R. fellea, and C. cornucopioides caused selective anti-inflammatory effects by decreasing NO and IL-6 in LPS-stimulated mouse macrophage cells. To date, limited information is available on the chemical composition of these anti-inflammatory species, but based on the activities demonstrated in this study these species warrant further investigation to determine the precise chemical identity of anti-inflammatory compounds responsible for the activities observed. In addition, while more detailed studies would be required, extracts from the five species outlined just now have demonstrated anti-inflammatory potential.
Footnotes
Acknowledgment
All authors read and contributed toward the final article.
Author Disclosure Statement
The authors declare that no competing financial interests exist.