
Abstract
Although several animal and cell studies have indicated that blackcurrant anthocyanins exert antioxidative and anti-inflammatory properties, which could potentially improve bone mass, the effect of blackcurrant on bone health has not been reported yet. Thus, this study was aimed to evaluate the effect of blackcurrant anthocyanins on bone mass in an estrogen deficiency mouse model. Fourteen-week-old C57BL/6J mice (n = 54) were ovariectomized or sham operated. The ovariectomized mice were divided into two groups, basal diet (OVX) or basal diet containing 1% anthocyanin-rich blackcurrant extract (OVX+BC), and sacrificed at 4, 8, and 12 weeks. Femoral bone mineral density (BMD) and trabecular bone volume by dual-energy X-ray absorptiometry and micro-computed tomography, respectively, and serum bone markers were measured. Ovariectomy significantly reduced BMD and trabecular bone volume at all time points (P < .05). Blackcurrant supplementation attenuated ovariectomy-induced bone loss measured by BMD and trabecular bone volume at 8 weeks (P = .055 and P = .057) and the effect was more pronounced at 12 weeks (P = .053 and P < .05). Ovariectomy and blackcurrant treatment did not alter serum biomarkers of bone formation and resorption. Bone marrow cells extracted from OVX mice significantly induced osteoclast-like (OCL) cell formation compared with cells from sham controls (P < .05). Blackcurrant treatment decreased the number of TRAP(+) OCL compared with OVX mice at 8 and 12 weeks (P < .05). Furthermore, blackcurrant supplementation reduced bone resorption activity when measured by resorption pit assay, compared with OVX group (P < .05). These results demonstrate that blackcurrant may be effective in mitigating osteoclast-induced postmenopausal bone loss.
Introduction
P
Plums and blueberries, which contain some of the same polyphenols as blackcurrant, have decreased bone loss in female rat models of osteoporosis. 9,10 We have recently reported that blackcurrants contain more anthocyanins than commonly well-known berries such as blueberries, blackberries, raspberries, or cranberries. 11 The major anthocyanin present in blackcurrants is delphinidin, which has the highest free-radical scavenging capacity among major anthocyanidins. 12 Several studies indicated that blackcurrant anthocyanins exhibited a range of health benefits, including antioxidant 13,14 and anti-inflammatory effects, 15 –17 which could potentially improve bone remodeling. However, the effects and mechanisms of blackcurrant on bone health are not well understood. Because osteoporosis is accelerated by postmenopausal ovarian hormone deficiency, ovariectomized rodent models have been widely used for studying osteoporosis. 18 Thus, this study aimed to determine whether blackcurrant anthocyanins prevent estrogen deficiency-induced bone loss using an ovariectomized mouse model.
Materials and Methods
In vitro study
Bone marrow macrophages (BMMs) were isolated from bone marrow (BM) cells collected from wild-type mouse femurs. BMMs were cultured with mouse macrophage colony-stimulating factor (M-CSF) and mouse receptor activator of nuclear factor kappa-B ligand (RANKL) both at 30 ng/mL (R&D Systems, Minneapolis, MN, USA). Blackberry was tested with blackcurrant to compare the relative effects on osteoclastogenesis at the same anthocyanin concentration. Freeze-dried blackcurrant and blackberry were provided by VDF FutureCeuticals (Momence, IL, USA). Polyphenols of the berries were extracted into 80% aqueous methanol containing 0.1% HCL and anthocyanin fractions from the berry polyphenol extracts were purified using Sep-Pak C18 Plus Short SPE Cartridge (Waters, Milford, MA, USA). 17 Blackcurrant and blackberry anthocyanins had no impact on cell viability at 1 and 3 μg/mL, while toxic effects were seen at 10 μg/mL and higher concentrations. Therefore, the study used 1 and 3 μg/mL as test dosages to compare the effects of blackcurrant anthocyanin and blackberry anthocyanin on osteoclast differentiation assay. Alkaline phosphatase (AP) assay (Sigma-Aldrich, St. Louis, MO, USA) was performed to compare the effects of blackcurrant and blackberry anthocyanins at 1 μg/mL on osteoblast differentiation.
In vivo study
Animals
Fifty-four 14-week-old female C57BL/6J mice were purchased from Jackson Laboratory (Bar Harbor, ME, USA), either ovariectomized (n = 36) or sham operated (Sham, n = 18) 1 week before delivery. Mice were housed two per cage in a temperature and humidity-controlled room in the Center for Laboratory Animal Care at the University of Connecticut. Sham mice (n = 18) were fed a basal diet (AIN-93M purified diet), whereas ovariectomized mice were randomly divided into two groups that were either fed a basal diet (n = 18, OVX) or basal diet containing 1% anthocyanin-rich blackcurrant extract (n = 18, OVX+BC) provided by Just the Berries Ltd. (Palmerston North, New Zealand).
Mice were sacrificed at weeks 4, 8, and 12 (n = 6 from each group). The uterine weight of mice was measured to confirm the ovariectomy. Based on the uterine weight, mice that failed to exhibit ovariectomy were excluded from the analysis As shown in Figure 1 analysis was performed using 52 mice, Sham (n = 18), OVX (n = 17), OVX+BC (n = 17), respectively. BM cells from long bones (tibia) were collected at 8th week and 12th week to evaluate blackcurrant anthocyanin effects on osteoclast differentiation and activity.
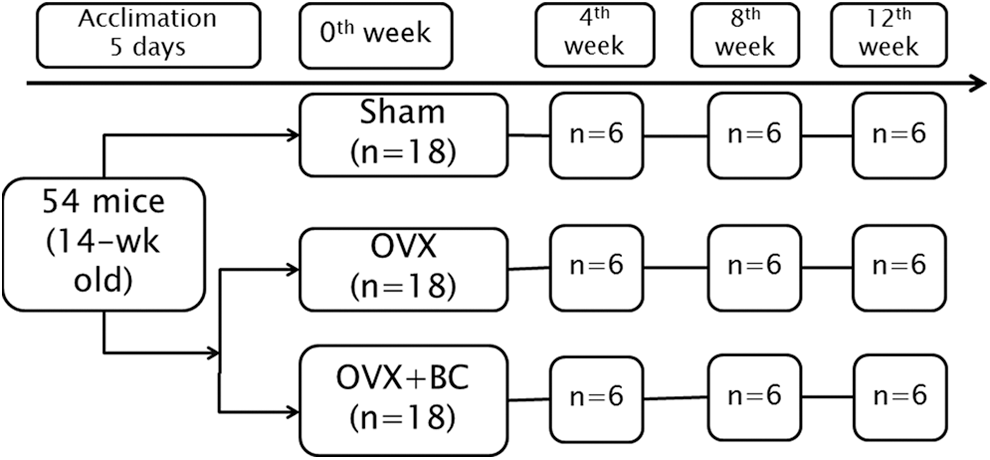
Study design.
Dual-energy X-ray absorptiometry and micro-computed tomography analyses
The left femur and first lumbar vertebrae (L1) were isolated and fixed in 70% ethanol at 4°C for bone density analysis by using dual-energy X-ray absorptiometry (DXA) to determine bone mineral density (BMD) and bone mineral content. The femur was further tested by using high-resolution X-ray micro-computed tomography (μCT 40; Scanco Medical, AG, Bassersdorf, Switzerland). These measurement techniques were performed in accordance with the recommendations of the Committee of the American Society for Bone and Mineral Research. 19 Trabecular and cortical morphometry was quantified within the metaphyseal region of the distal femur. Three-dimensional images of femur were reconstructed. Trabecular morphometry was assessed by the bone volume fraction (an indicator of trabecular bone volume), trabecular thickness (Tb.Th), trabecular number (Tb.N), and trabecular spacing (Tb.Sp). Cortical morphometry was analyzed by assessing average thickness and cross-sectional area within a 600 μm long section at mid-diaphysis of the femur.
Bone histomorphometric analysis
Histomorphometric analysis was performed on the right femur of mice fed either the blackcurrant or basal diet for 12 weeks. Femurs were embedded in paraffin and their sections were stained with tartrate-resistant acid phosphatase (TRAP) for quantification of osteoclasts (OC). All measurements were confined to the secondary spongiosa and within an area between 400 and 200 μm distal to the growth plate-metaphyseal junction of the distal femur. OC was identified as multinucleated TRAP(+) cells adjacent to bone. Histomorphometric data were collected using the OsteoMeasure System (Osteo-Metrics, Inc., Atlanta, GA, USA). Statistical cancellous bone endpoints included cellular (OC and osteoblasts [OB]) and trabecular bone parameters (BV/TV, Tb.Th, and Tb. Sp). OC/OB perimeters were expressed as percent of total bone perimeter: Oc.S/BS, Ob.S/BS, ES/BS, and N.Oc/T.Ar.
Analysis of serum bone turnover markers
Serum samples were collected after an overnight fast. Two serum bone turnover markers, C-terminal telopeptide of type I collage (CTX, bone resorption marker) and amino-terminal propeptide of type I collagen (P1NP, bone formation marker) were measured using a RatLaps ELISA kit and a Rat/Mouse P1NP ELISA kit (Immunodiagnostic Systems, Scottsdale, AZ, USA), respectively.
Ex vivo study
BM cell culture
BM cells were obtained by flushing the tibia of mice in all treatment groups at 8 and 12 weeks with minimum essential medium alpha (α-MEM; Gibco, Life Technologies, Grand Island, NY, USA) using a 25-gauge needle. Collected cells were spun down and resuspended in α-MEM supplemented with 10% heat-inactivated fetal bovine serum (Hyclone, Logan, UT, USA) and 10% penicillin in the presence of mice M-CSF and RANKL both at 30 ng/mL (R&D Systems). 20
Osteoclast formation assay
The number of viable BM cells was determined using trypan blue. About 5 × 104 cells were plated in each well of 96-well plates in 200 μL of complete α-MEM with 30 ng/mL M-CSF and RANKL for up to 6 days, with medium change on day 3. 20 The cultured cells were fixed with 2.5% glutaraldehyde (Polysciences, Warrington, PA, USA) in phosphate-buffered saline (PBS) and strained for TRAP using the leukocyte acid phosphatase kit according to manufacturer's instructions (Sigma-Aldrich). TRAP(+) multinucleated cells (more than three nuclei) were counted as osteoclast-like (OCL) cells.
Pit formation assay
The ability of OC to resorb bone was assessed by culturing them on the surface of ultraviolet-sterilized devitalized bovine cortical bone slices (4 × 4 × 0.2 mm) that were placed in the wells of a 96-well plate in α-MEM with M-CSF and RANKL (30 ng/mL) for up to 14 days. The medium was changed every 3 days. The bone slices were fixed with 2.5% glutaraldehyde in PBS and stained for TRAP. TRAP(+) multinucleated OCL cells were visualized by inverted light microscopy to confirm osteoclast differentiation. Bone slices were then sonicated in 0.25 NH4OH to remove OCL cells. The bone slices were stained with 1% toluidine in borax buffer to detect the resorption pits. To assess the resorption potential of OCL cells, the areas (μm2) of 100 or 125 individual pits per group were measured and recorded using the via-160 video image measurement system (Boeckeler Instruments, Tucson, AZ, USA). Mean of resorption areas per OC and standard error of mean were calculated.
Statistical analysis
Results were expressed as mean ± standard error. Two-way analysis of variance and two tailed t-tests were used to determine significant differences among groups. Differences were considered to be significant if P < .05.
Results
Blackcurrant treatment significantly decreased osteoclast differentiation
Delphinidin-3-glucoside and cyanidin-3-O-rutinoside are the major anthocyanins in blackcurrant, while cyanidin-3-O-rutinoside is the major anthocyanin in blackberry. To compare the effects of blackcurrant and blackberry anthocyanins on osteoclast differentiation, an osteoclast formation assay was performed. The results showed that blackcurrant anthocyanins decreased osteoclast differentiation to a greater extent than blackberry anthocyanins in a dose-dependent manner (Fig. 2). A bone formation assay was performed to further evaluate the effects of the anthocyanins from these two berries on osteoblastic differentiation by using AP staining. However, there was no evidence of a difference among groups.
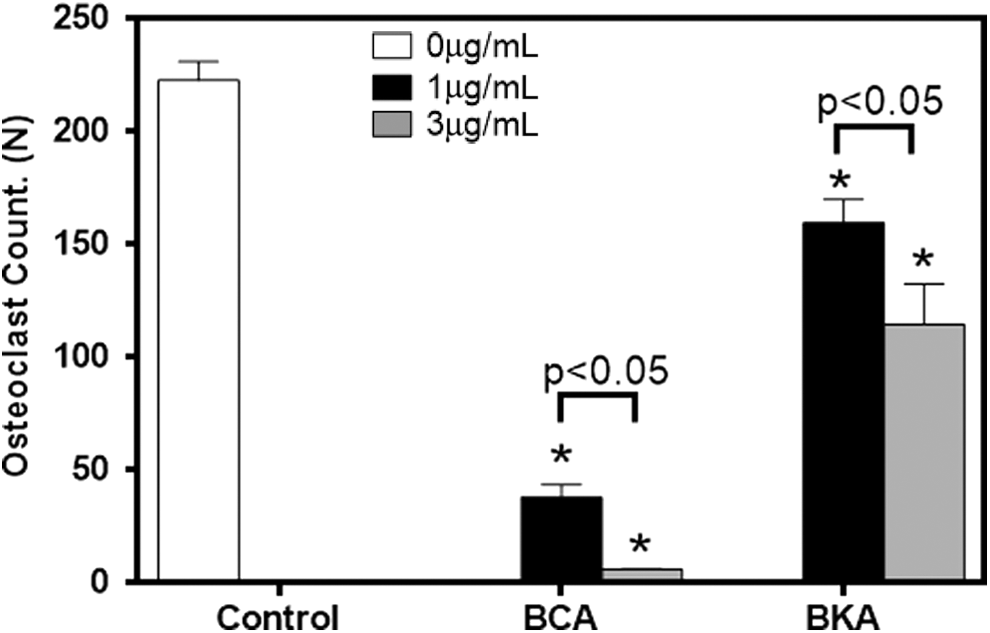
Effect of BCA and BKA on osteoclast formation. Values are reported as mean ± SEM. Data were analyzed by one-way ANOVA and Tukey's test. *Indicates significant effect of berry anthocyanins. P-value represents comparison among different treatment dosages. ANOVA, analysis of variance; BCA, blackcurrant anthocyanin; blackberry anthocyanin; SEM, standard error of mean.
Blackcurrant diet had no effect on ovariectomy-induced weight gain
To evaluate the effects of ovariectomy, mouse body weight and uterine weight were measured at the time of sacrifice. Body weight showed that ovariectomy significantly increased the body weight of mice in OVX and OVX+BC groups compared with Sham group at 4, 8, and 12 weeks (P < .05). In addition, there was a decrease in uterine weight in these two groups compared with Sham (P < .05), which further confirmed ovariectomy by uterine atrophy. Mice that did not exhibit uterine atrophy in OVX groups (n = 2) were eliminated from analysis. Blackcurrant treatment did not attenuate uterine atrophy (Table 1).
Values represent the mean ± SEM. No interaction between treatment and time was found by using two-way ANOVA. Multiple comparisons were performed by using Tukey's test.
Indicates significant effect of ovariectomy.
Indicates significant effect of blackcurrant supplementation (P < .05).
Sham, mice with sham operation; OVX, ovariectomized mice fed with basal diet; OVX+BC, ovariectomized mice fed with basal diet containing 1% anthocyanin-rich blackcurrant extract.
ANOVA, analysis of variance; SEM, standard error of mean.
Blackcurrant supplementation decreased ovariectomy-induced bone loss
To examine the effect of blackcurrant supplementation on bone, DXA analysis was performed. DXA analysis indicated that ovariectomy significantly decreased femur BMD over time, while blackcurrant supplementation started to improve BMD at 8 weeks (P = .055) and the effects were statistically significant at 12 weeks (P < .05) (Fig. 3). L1 BMD was also measured and showed a similar trend as that of femur in DXA analysis, however, there were no statistically significant differences.
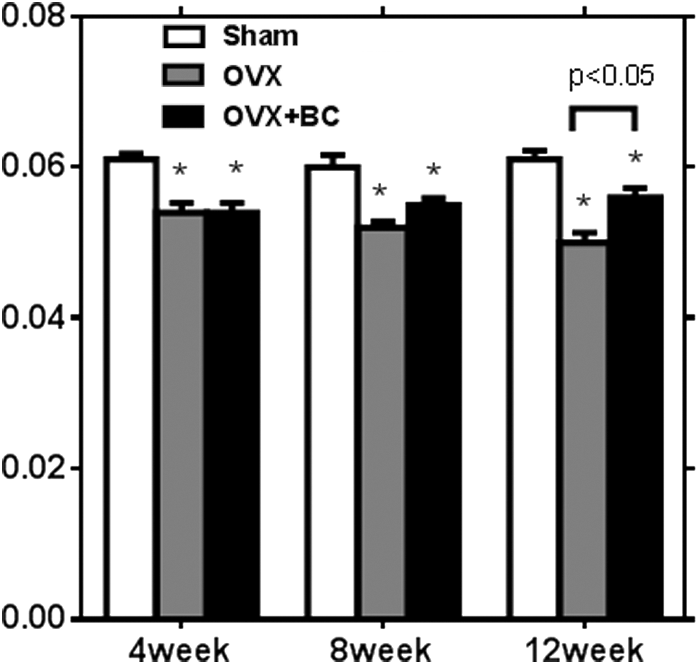
BMD in femur by DXA analysis. Values are reported as mean ± SEM for three time points. Data were analyzed by one-way ANOVA and Tukey's test. *Indicates significant effect of ovariectomy. P-value represents comparison between OVX and OVX+BC groups. BMD, bone mineral density; DXA, dual-energy X-ray absorptiometry; OVX, ovariectomized mice fed with basal diet; OVX+BC, ovariectomized mice fed with basal diet containing 1% anthocyanin-rich blackcurrant extract.
Since DXA represents both cortical and trabecular bone volume, we performed μCT analysis to determine whether blackcurrant supplementation alters the most metabolically active trabecular bone. Similar results were found in 3D μCT analysis (Fig. 4). According to the μCT image (Fig. 4a), blackcurrant anthocyanin supplementation showed a significant protective effect on both trabecular and cortical bones after 12 weeks of treatment. Ovariectomy significantly reduced trabecular bone volume at all time points we examined. This was due to a decrease in trabecular thickness and number, and increase in trabecular spacing. Blackcurrant supplementation has a significant mitigating effect on trabecular bone loss at 8 weeks (P = .057) and at 12 weeks (P = .053) due to increasing trabecular thickness and number, and decreasing trabecular spacing, which lead to partial reversal of ovariectomy-induced trabecular bone loss (Fig. 4b).
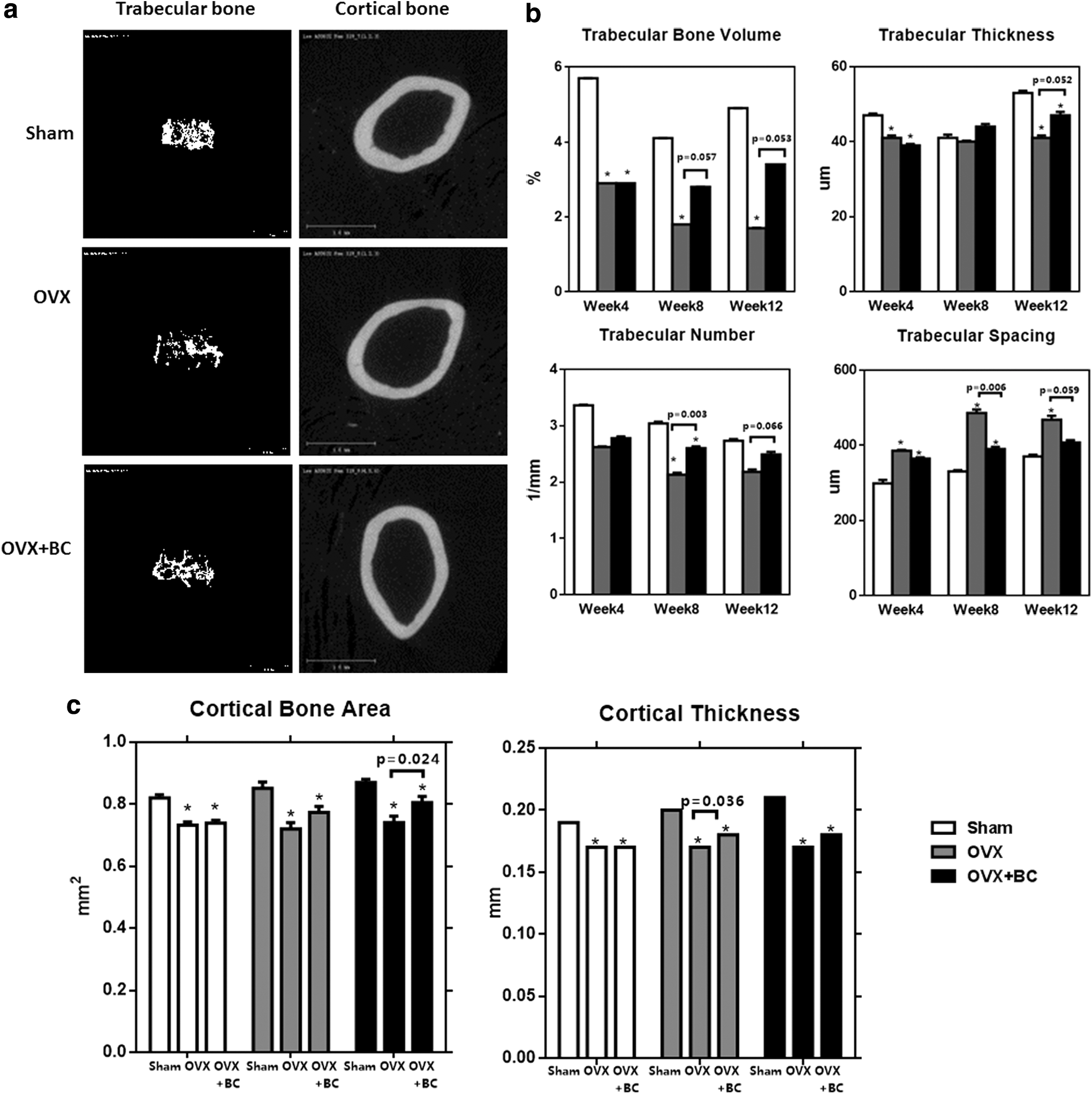
μCT analysis and images.
Since there was significant alteration in BMD, we also examined the cortical bone parameters by μCT as shown in Figure 4c. Ovariectomy significantly decreased cortical bone area and cortical thickness compared with sham at all time points examined. Blackcurrant supplementation partially reversed OVX-induced cortical bone loss at 12 weeks (P = .024) showing a trend toward the increase at 8 weeks compared with OVX. Further, there was significant difference in cortical thickness at both week 4 (P = .014) and week 8 (P = .036) in the two OVX groups.
To examine the effect of blackcurrant anthocyanin consumption on bone cells, static histomorphometry was performed. Since the greatest reduction in ovariectomy-induced bone loss was achieved by 12 weeks of treatment, based on the results from μCT and DXA analysis, only bones from mice supplemented with control or blackcurrant diet for 12 weeks were assessed by quantitative histomorphometric analysis. As shown in μCT, ovariectomy significantly downregulated trabecular bone volume compared with sham.
Blackcurrant supplementation partially reversed ovariectomy-induced trabecular bone loss, but this was not statistically significant. To examine whether the effect of blackcurrant supplementation was due to changes in OC or OB, we further examined the parameters of osteoblast and osteoclast in vivo. Blackcurrant supplementation or ovariectomy did not alter osteoblast or osteoclast parameters.
Although blackcurrant supplementation prevented ovariectomy-induced bone loss, results from quantitative histomorphometric analysis did not demonstrate changes in bone turnover by altering OC and OB numbers (Table 2). Thus, serum bone turnover markers (P1NP and CTX) were measured to assess changes in bone turnover. CTX, which represents bone resorption, increased in OVX mice at 8 weeks but decreased at 12 weeks, which were not significant (Fig. 5). There was no significant difference in the bone formation marker P1NP among the three groups over the time.
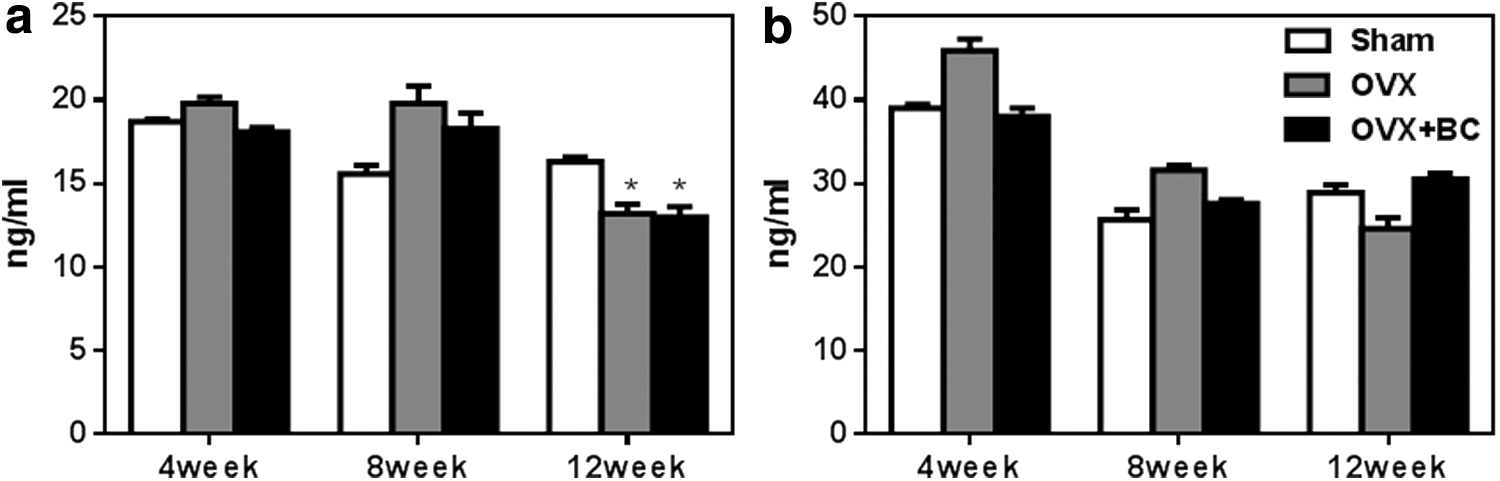
Serum biochemistry bone turnover markers.
Values represent the mean ± SEM. No interaction between treatment and time was found by using two-way ANOVA. Multiple comparisons were performed by using Tukey's test.
Tb.Th, trabecular thickness; Tb.Sp, trabecular spacing.
To determine whether blackcurrant anthocyanins affect osteoclast precursor cells in vivo, BM cells from Sham, OVX, OVX+BC mice were collected and allowed to differentiate into OCL cells ex vivo in the presence of M-CSF and RANKL for 5 days. BM cells from ovariectomized mice significantly increased osteoclast formation 1.5-fold compared with Sham controls (P < .05). Furthermore, BM cells from blackcurrant supplemented OVX mice downregulated TRAP(+) osteoclast formation compared to OVX at eighth week (P < .05) (Fig. 6a) indicating that in vivo supplementation of blackcurrant significantly affected osteoclastic potential of precursor cells to differentiate.
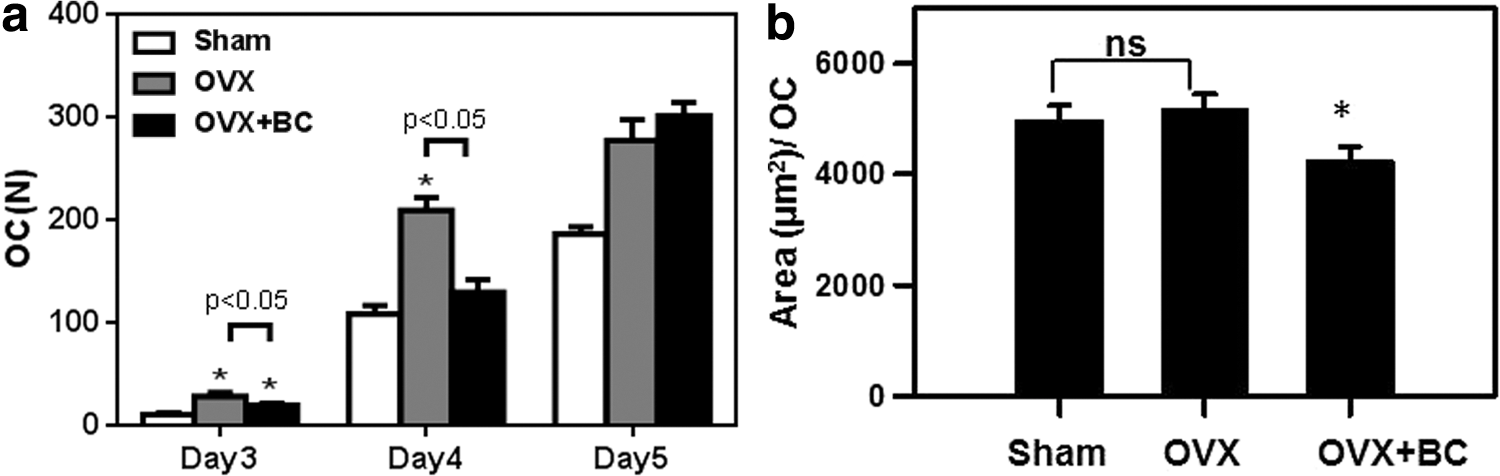
Osteoclasts activity (pit formation assay) and differentiation (TRAP assay). Week 8 osteoclast TRAP(+) number
We next examined whether blackcurrant supplementation affected bone resorption. BM cells from Sham, OVX, and OVX+BC were cultured on bone slices in the presence of M-CSF and RANKL as described and bone resorption area by individual osteoclast was evaluated. Interestingly, BM cells from ovariectomized mice did not affect bone resorption compared to those from Sham controls (Fig. 6b). However, BM cells from in vivo blackcurrant supplementation significantly decreased bone resorption.
Discussion
Osteoporosis is associated with inadequate bone mass and bone formation along with excessive bone resorption. 21,22 In this study, we attempted to examine whether blackcurrant anthocyanins can alleviate ovariectomy-induced bone loss. Both OVX and OVX+BC groups significantly increased in body weight compared to the Sham group, which was attributed to a postovariectomy estrogen deficiency as reported earlier. 23 There was no significant difference in body or uterine weight between OVX and OVX+BC groups indicating that blackcurrant extract may not mimic estrogen effect.
Osteoporosis is characterized by a slow decline in bone mass and impaired micro-architecture and leads to an increased risk of fracture. 24,25 It involves disruption of the balance between bone formation and resorption through uncoupling the balance between OB and OC. Since estrogen is important for maintaining bone density, its deficiency after menopause may accelerate bone loss. 26 Previous studies have shown that inflammation and oxidative stress play an important role in bone remodeling and osteoporosis development. 27 Our BMD findings suggest blackcurrant anthocyanins significantly protect against ovariectomy-induced bone loss over time. This preventive effect might be due to anthocyanins, the antioxidant components of blackcurrant through lowering oxidative stress and inflammation. 28 –30
The deterioration of the trabecular architecture is in agreement with decreased bone strength and increased fracture incidence in humans. 31 Thus, it is important to evaluate the effect of blackcurrant supplementation on the quality of bone structure in addition to BMD. μCT techniques assessing structural parameters in both trabecular and cortical bones provide better information on the bone microarchitecture. The μCT data indicate that blackcurrant anthocyanins can significantly improve these structural indices and repair the lost trabecular connectivity and further prevent the progression of trabecular architecture deterioration. Thus, long-term blackcurrant consumption may reverse postmenopausal bone loss and prevent the development of osteoporosis.
Bone turnover markers have been widely used to measure how drugs affect bone remodeling. 32 CTX and P1NP were selected as biochemical markers and tested in serum samples to assess bone resorption and formation. In theory, ovariectomy should alter both bone resorption and formation markers. However, these indices were not different among the three groups over time. This may be due to changes in metabolic bone turnover that occurred before the bone mass loss and this study might not capture the right time point. Aging as a confounding factor should also be considered. Although these mice were young at the beginning of the study, ovariectomy might disrupt their physiological balance and accelerate their aging process.
To study the mechanistic effects of blackcurrant on osteoclastogenesis, this study tested the osteoclastogenic activity of OC progenitor population from BM, which was cultured with M-CSF and RANKL. In OCL formation assay (TRAP), there was a linage development that OCL cells started osteoclastogenesis at third day and at fifth day reaching their peak of differentiation. OCL cells from OVX+BC had slightly different behaviors in these two collections, which made it hard to be interpreted. Nevertheless, there was a tendency to delay and even decrease the formation of OCL cells. Since cells were collected separately, there might have been uncontrolled factors that influenced the consistency of this trend. The ability to isolate highly purified primary BM populations plays an important role in determining the degree of osteoclastogenic lineage development and evaluating the factors that regulate their ability to form OCL cells. 33 Additionally, cells collected from mice treated for 12 weeks were older, which made aging a confounder again. At the same time, the resorption ability of OCL cells was further evaluated by measuring the resorption area on a bone chip. The cells collected from OVX+BC group had significant decrease in resorbing activity comparing to the other two groups. Therefore, blackcurrant anthocyanins might alleviate bone loss through suppressing osteocastogenesis and mediating osteoclast function.
In conclusion, our findings demonstrated that blackcurrant anthocyanins are effective in ameliorating bone loss caused by ovarian hormone deficiency. The effects of blackcurrant were apparent in both trabecular bone and cortical bone. Blackcurrant anthocyanins might work by inhibiting osteoclastogenesis and decreasing osteoclast activity. Further studies are needed to identify the major bioactive component of blackcurrant anthocyanins, the lowest effective dose, and the underlying mechanism through in vivo and in vitro studies.
Footnotes
Acknowledgment
This study was funded by the USDA/National Institute of Food and Agriculture (#2016-67018-24492) and University of Connecticut USDA Hatch Grants to Drs. Ock K. Chun and Sung I. Koo.
Author Disclosure Statement
No competing financial interests exist.