
Abstract
Lactoferrin (LF) has beneficial effects against various diseases. However, the effects of LF on liver fibrosis in systematic lupus erythematosus (SLE) are unknown. In this study, NZB/W F1 mice were utilized to investigate the effects of LF on SLE. Experiments reveal that LF significantly increases glutathione and 1,1-diphenyl-2-picryl-hydrazyl levels and significantly decreased malondialdehyde levels in both serum and liver in NZB/W F1 mice. LF also lowered matrix metalloproteinase-9 activity and liver inflammatory indices, such as aminotransferase and alanine aminotransferase. Notably, significantly decreased expression of fibrotic related molecules, including transforming growth factor (TGF)-β1, tumor necrosis factor-α, interleukin-1β, and TGF-β1 receptor, were observed in the livers of NZB/W F1 mice that had been treated with LF. Significantly, suppressed Smad2/3 signaling, α-smooth muscle actin, and collagen deposition were also detected. These findings reveal that LF has beneficial effects on SLE by increasing antioxidant activities and ameliorating liver inflammation and fibrosis, suggesting the therapeutic effectiveness of LF against SLE.
Introduction
S
Accumulating evidence reveals that liver fibrosis is involved in SLE and is associated with its pathogenesis. 7,8 Transforming growth factor (TGF)-β and Smads are known to be central regulators in chronic liver disorders, which contribute to fibrogenesis through inflammation. 9 Engagement with TGF-β–activated TGF-β type I receptor initiates the phosphorylation of Smad proteins, including Smad2 and Smad3. 10,11 The phosphorylated Smad2 and Smad3 (p-Smad2/3) proteins then interact with Smad4 and are then translocated to the nucleus to regulate the expression of fibrotic genes such as plasminogen activator inhibitor (PAI)-1. 12 The level of PAI-1 is significantly increased in a fibrotic liver, contributing to excessive accumulation of collagen and other extracellular matrix proteins. 13 The presence of α-smooth muscle actin (SMA) is also regarded as crucial to the fibrogenic response. 14 These processes lead to liver cirrhosis, which is ultimately fatal.
Lactoferrin (LF) is an 80 kDa iron-binding glycoprotein of the transferrin family that is widely distributed in colostrum and milk. 15 LF has central roles in a large range of biological activities, such as iron metabolism, antimicrobial activities, antioxidation, anticancer, and immune modulation. 16 –19 An earlier study reported that NZB/W F1 mice that were treated for a prolonged period with bovine LF exhibited a reduced frequency of the positive Coombs' reaction. 20 Other studies have found that orally administrated LF lowers the clinical score and prevents pathohistological changes in the spinal cord in rats with experimentally induced autoimmune encephalomyelitis. 21 These findings suggest that LF has a beneficial effect on autoimmune diseases, including SLE.
Along with liver abnormalities, decreased antioxidant activities and increased lipid peroxidation are also significantly correlated with elevated inflammatory cytokines and disease activity in SLE. 22,23 However, understanding of the effects of LF on the antioxidant activities and liver fibrosis in SLE is still limited. This study investigated whether LF increases antioxidant activities and ameliorates hepatic fibrosis in NZB/W F1 mice, focusing on the relevant underlying mechanisms.
Materials and Methods
Mice and diets
Female NZB/W F1 mice, a well-described lupus-prone mouse strain, 24 were purchased from Jackson Laboratory (Bar Harbor, ME, USA) and housed under supervision of the Institutional Animal Care and Use Committee at Chung Shan Medical University. The mice were housed in an animal room at 22°C with a 12-h light/12-h dark cycle under supervision of the Institutional Animal Care and Use Committee at Chung Shan Medical University (IACUC approval No. 1456). Chow diet, soybean oil, and cholesterol were purchased (TestDiet Division, PMI Nutrition International/Purina Mills LLC, Richmond, IN, USA). LF from bovine milk was purchased from Sigma-Aldrich (St. Louis, MO, USA). The dosages of LF used in this study are modified according to a previous study. 21 The control diet was composed of 93% rodent 5001 chow diet and 7% soybean oil (Ctrl). The cholesterol diet was composed of 92% rodent 5001 chow diet, 7% soybean oil, and 1% cholesterol (Chol). The cholesterol plus low-dose LF diet was composed of 91.5% rodent 5001 chow diet, 7% soybean oil, 1% cholesterol, and 0.5% LF (LLF). The cholesterol plus high-dose LF diet was composed of 91% rodent 5001 chow diet, 7% soybean oil, 1% cholesterol, and 1% LF (HLF). Forty 12-week-old female NZB/W F1 mice were divided into four groups (10 mice/group) and fed different diets ad libitum for 12 weeks. Body weights of the mice were measured weekly and no significant variation was detected among the four groups. Mice were sacrificed at the age of 24 weeks by CO2 asphyxiation and were fasted overnight before sacrifice. Heart blood and livers of the mice were obtained after CO2 sacrifice and stored at −80°C until use.
Measurement of glutathione levels
The glutathione level was determined as previously described. 25 Measurements of concentrations of reduced glutathione (GSH) were performed with GSH Assay Kit (Chemicon Inc., Temecula, CA, USA) according to the manufacturer's instructions. Briefly, fluorescence was measured at 380/461 nm in 90 μL of protein extracts and 10 μL of prepared monochlorobimane solution incubated in the dark for 2 h at 25°C using a 96-well fluorometric plate reader.
DPPH assay
The radical scavenging activity was determined using a 1,1-diphenyl-2-picryl-hydrazyl (DPPH) scavenging activity as previously described. 25 Briefly, a solution of 180 μL of 0.1 mM DPPH solution in ethanol was gently mixed with 20 μL sample in ethanol. The value of DPPH absorption was measured at 517 nm by a 96-well fluorometric plate reader. DPPH radical scavenging activity was expressed as % inhibition compared to the blank (ethanol).
Thiobarbituric acid reductase assay
Lipid peroxidation was measured using thiobarbituric acid reductase (TBAR) assay as previously described. 23 Briefly, to make a 10% homogenate, a volume of 200 mg serum or tissue was added to 2 mL of 1.15% KCl and mixing was performed with a homogenizer (Kontes Glass, Vineland, NJ, USA). A quantity of 3 mL of 1% phosphoric acid and 1 mL of 0.6% TBA solution were added to 0.5 mL of 10% tissue homogenate. The mixture was left in a boiling water bath for 45 min. After cooling, 4 mL of n-butanol was added and mixed vigorously. The butanol phase was separated by centrifugation at 500 g, and absorbance was measured spectrophotometrically at 535 nm. The difference was used as the TBAR value, and the results were calculated as nanomoles per gram of tissue.
Gel zymography
Matrix metalloproteinase (MMP)-2 and MMP-9 activities were analyzed by gelatin zymography assays as previously described. 26 Ten microliters of 10-fold diluted serum or 25 μg protein lysates of liver tissue from NZB/W F1 mice were separated onto an 8% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) gels containing 0.1% gelatin. Gels were washed for 30 min in 2.5% Triton X-100 to remove the SDS and then soaked in the reaction buffer containing 40 mM Tris-HCl, pH 8.0, 10 mM CaCl2, and 0.02% NaN3 for 30 min. The gels were incubated at 37°C for 24 h after removing to a fresh reaction buffer. Gelatinolytic activity was visualized by staining the gels with 0.5% Coomassie brilliant blue R-250, destaining with methanol-acetic acid water, and relative MMP levels were quantified by a gel documentation and analysis system (Appraise; Beckman-Coulter, Brea, CA, USA).
Enzyme-linked immunosorbent assay
The serum biochemical values of aspartate aminotransferase (AST) and alanine aminotransferase (ALT) were determined using commercial enzymatic kits (Cobas Mira, Roche, Basel, Switzerland) with the SPOTCHEM EZ SP-4430 automated analyzer (ARKRAY, Inc., Kyoto, Japan) according to manufacturer's instruction.
Preparation of tissue extract and determination of protein
All procedures were performed at 4°C in a cold room as previously described. 26 The liver tissues obtained from NZB/W F1 mice were homogenized in 600 μL PRO-PREP solution (iNtRON Biotech, Gyeonggi-do, Korea) by 30 strokes using a Dounce Homogenizer (Kontes Glass, Vineland, NJ, USA). The homogenates were centrifuged at 19,083 g for 10 min at 4°C and the supernatant was then stored at −80°C until use. The protein content in the supernatant was measured according to the procedures of a Bio-Rad Protein Assay Kit (Cat. No.: 500-0006; Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Western blotting
Western blotting was performed as described in our previous study. 25 Protein samples were separated in 10% or 12.5% SDS-PAGE and electrophoretically transferred to a nitrocellulose membrane (Amersham Biosciences, Piscataway, NJ, USA) described elsewhere. 27 After blocking with 5% nonfat dry milk in 1 × phosphate-buffered saline (PBS), antibodies against TGF-β1, TGF-β1 receptor, tumor necrosis factor (TNF)-α, interleukin (IL)-1β, p-Smad2/3, Smad2/3, specificity protein (Sp)-1, PAI-1, α-SMA (Santa Cruz Biotechnology, Santa Cruz, CA, USA), and β-actin (Upstates, Charlottesville, VA, USA) were diluted in PBS with 2.5% bovine serum albumin and incubated for 1.5 h with gentle agitation at room temperature. The membranes were washed twice with PBS-Tween for 1 h, and secondary antibody conjugated with horseradish peroxidase (HRP) (Santa Cruz Biotechnology) was added. Pierce's SuperSignal West Dura HRP Detection Kit (Pierce Biotechnology, Inc., Rockford, IL) was used to detect antigen–antibody complexes. Quantified results were performed by densitometry (Appraise, Beckman-Coulter, Brea, CA, USA).
Masson's trichrome staining
A small piece of liver tissue was collected and fixed in 10% buffered formalin. The sample was then embedded in paraffin, sliced into 5-μm-thick sections, and Masson's trichrome staining was performed, which stained collagen blue and hepatocytes red. Images were obtained using a Zeiss Axiophot microscope (Zeiss, Oberkochen, Germany).
Statistical analysis
All statistical analyses were performed using SPSS 10.0 software (SPSS, Inc., Chicago, IL, USA). One-way analysis of variance and the least significant difference test were used to differentiate the differences between treatments. A significant difference was used at the 0.05 probability level and indicated by an asterisk. Three independent experiments were repeated, and the values are expressed as the mean ± SEM.
Results
LF increases antioxidant activities and reduces hepatic inflammatory indices in lupus-prone mice
To investigate the effects of LF on the antioxidant capacity in lupus-prone mice, GSH levels were measured, and DPPH assay and TBAR assay were performed. According to the results thus obtained, GSH and DPPH levels in both the serum and liver samples from NZB/W F1 mice that received either a low or a high LF diet were significantly higher than in those mice that received a control or high-cholesterol diet (Fig. 1A–D). Both serum and liver malondialdehyde (MDA) levels were significantly lower in NZB/W F1 mice that had received either a low or a high LF diet than in those mice that received a control or high-cholesterol diet, respectively (Fig. 1E, F). Indices of liver inflammation were obtained by determining the liver MMP activity and serum levels of AST and ALT. Figure 2A presents the ratio of MMP-9 activity to MMP-2. Significantly reduced hepatic MMP-9 activity was detected in NZB/W F1 mice that received either a low or a high LF diet than in those mice that received a control or high-cholesterol diet. The lower panel of Figure 2A compares MMP-9 activity with MMP-2 activity. Similarly, both serum AST and ALT levels were significantly lower in NZB/W F1 mice that had received either a low or a high LF diet than in those mice that received a control or high-cholesterol diet (Fig. 2B, C). Since low and high LF diets had similar effects on NZB/W F1 mice, only the results of the following experiment concerning the mice that received the control, high cholesterol, or low-dose LF diet are presented.
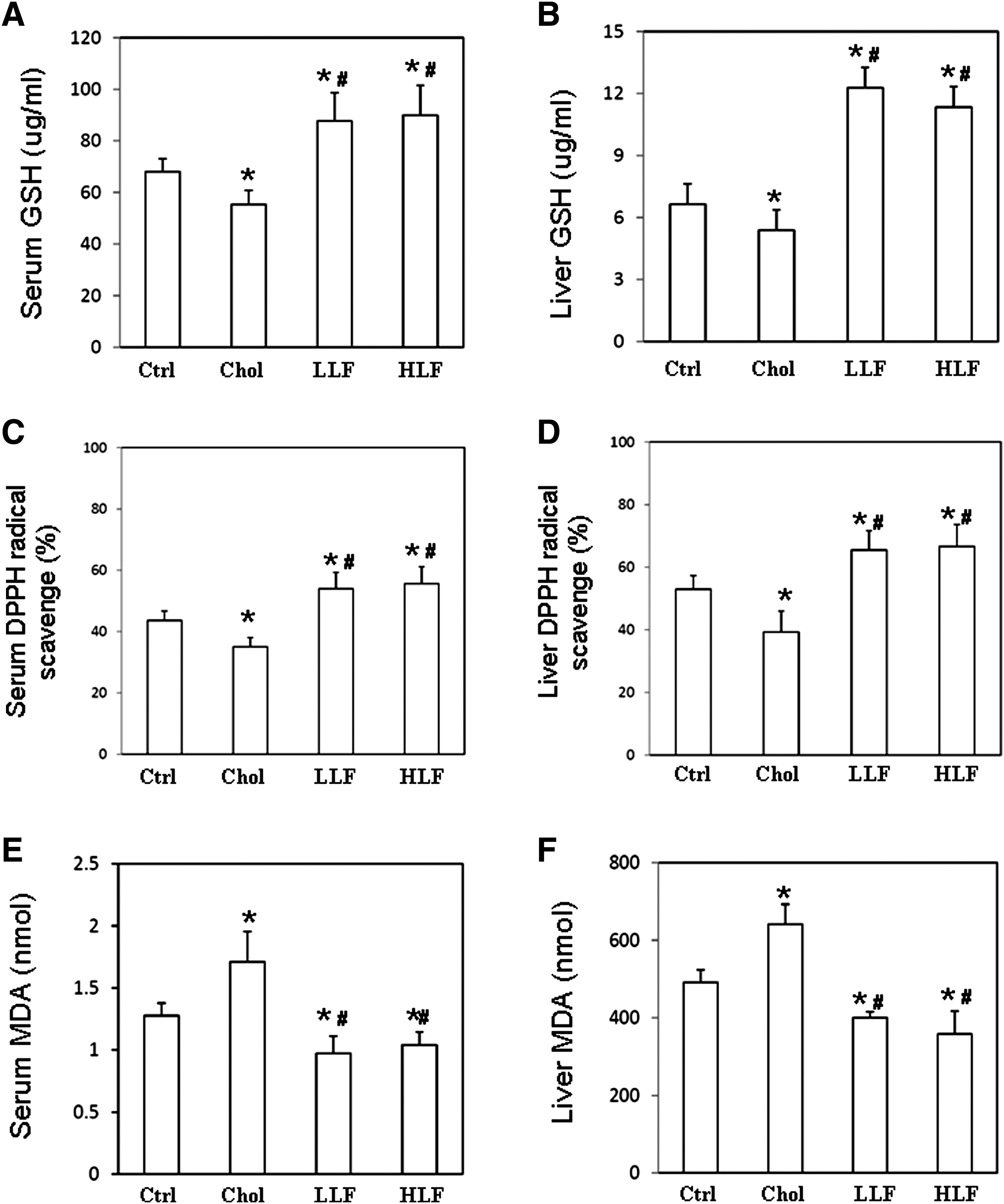
The serum and liver levels of
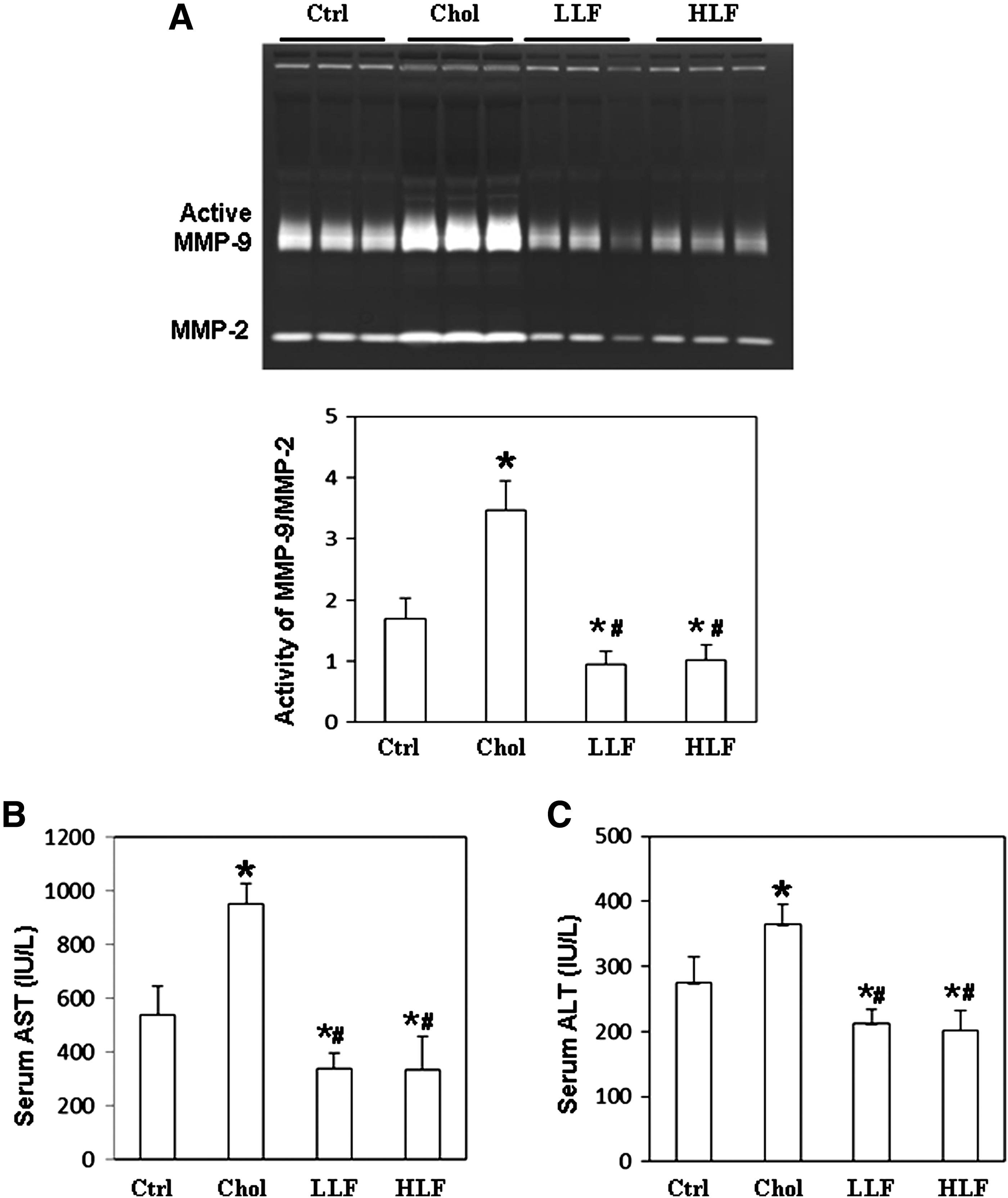
Detection of
LF reduces expressions of fibrotic related molecules in livers of lupus-prone mice
To investigate the effects of LF on hepatic fibrosis in NZB/W F1 mice, the expressions of fibrotic related cytokines, including TGF-β1, TNF-α, IL-1β, and the TGF-β1 receptor, were measured. Significantly reduced expressions of TGF-β1 and TGF-β1 receptor were detected in livers of NZB/W F1 mice that had received a low LF diet, rather than in those mice that had received a control or high-cholesterol diet (Fig. 3A). Figure 3B and C presents the TGF-β1 and TGF-β1 receptor, respectively, relative to that of β-actin. Significantly lower expressions of TNF-α and IL-1β were also detected in livers of NZB/W F1 mice that had received a low LF diet, but not in those mice that had received a control or high-cholesterol diet (Fig. 3A). Figure 3D and E compares the TNF-α and IL-1β, respectively, with that of β-actin.
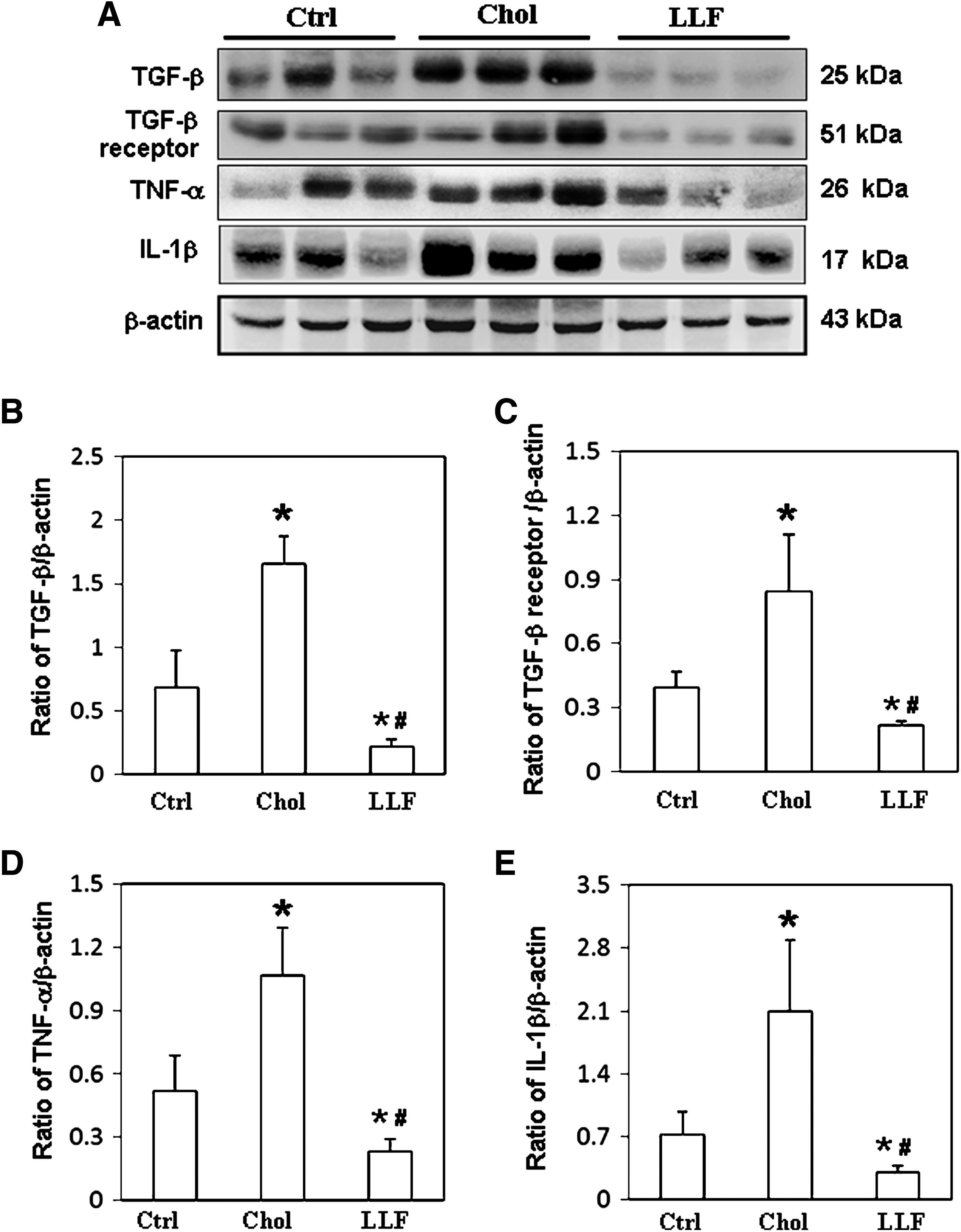
Expression of TGF-β1, TGF-β1 receptor, TNF-α, and IL-1β. Liver lysates obtained from the NZB/W F1 mice receiving control, high-cholesterol, and low-dose LF diet were probed with antibodies against
LF ameliorates hepatic fibrosis by reducing Smad2/3 signaling in lupus-prone mice
To investigate the mechanism by which LF affects hepatic fibrosis, the expressions of Smad signaling molecules were examined. Significantly lower p-Smad2/3 levels were detected in the livers of NZB/W F1 mice that had received a low LF diet than in those mice that had received a control or high-cholesterol diet (Fig. 4A). Figure 4B compares the p-Smad2/3 relative to that of Smad2/3. Similarly, a significantly lower expression of SP-1 was detected in the livers of NZB/W F1 mice that had received a low LF diet than in those mice that had received a control or high-cholesterol diet (Fig. 4A). Figure 4C compares the SP-1 relative to that of β-actin. The expressions of PAI-1, an inhibitor of fibrinolysis, and of α-SMA, a typical fibrosis marker, were also examined to clarify the effects of LF on hepatic fibrosis in NZB/W F1 mice. Significantly lower expressions of PAI-1 and α-SMA proteins were detected in the livers of NZB/W F1 mice that had received a low LF diet than in those mice that had received a control or high-cholesterol diet (Fig. 5A). Figure 5B and C compares the PAI-1 and α-SMA with that of β-actin. Furthermore, collagen deposition (blue area) in liver sections from NZB/W F1 mice that had received a low LF diet was markedly reduced compared to those mice that had received a control or high-cholesterol diet (Fig. 5D).
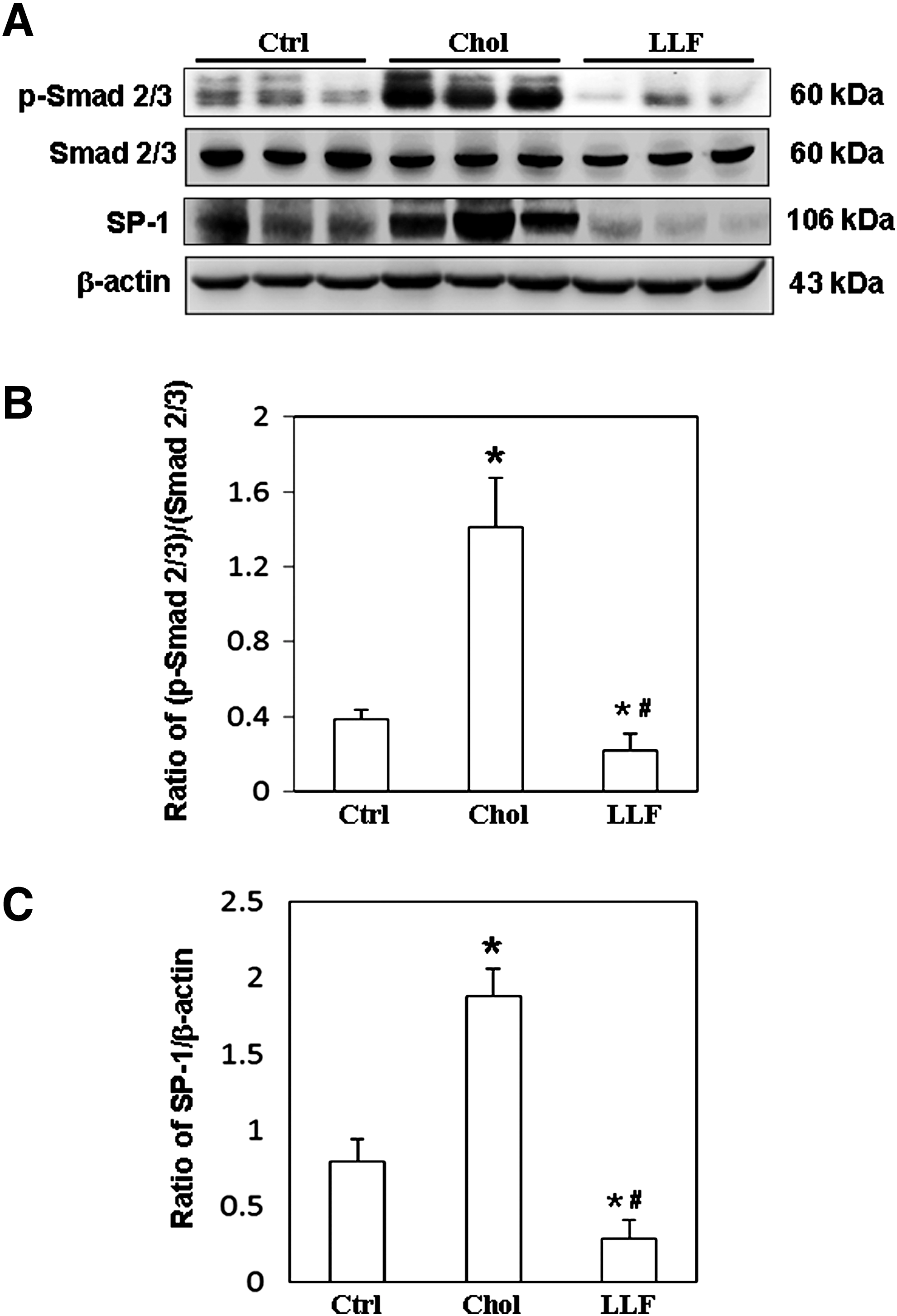
Expression of p-Smad2/3, Smad2/3, and SP-1. Liver lysates obtained from the NZB/W F1 mice receiving control, high-cholesterol, and low-dose LF diet were probed with antibodies against
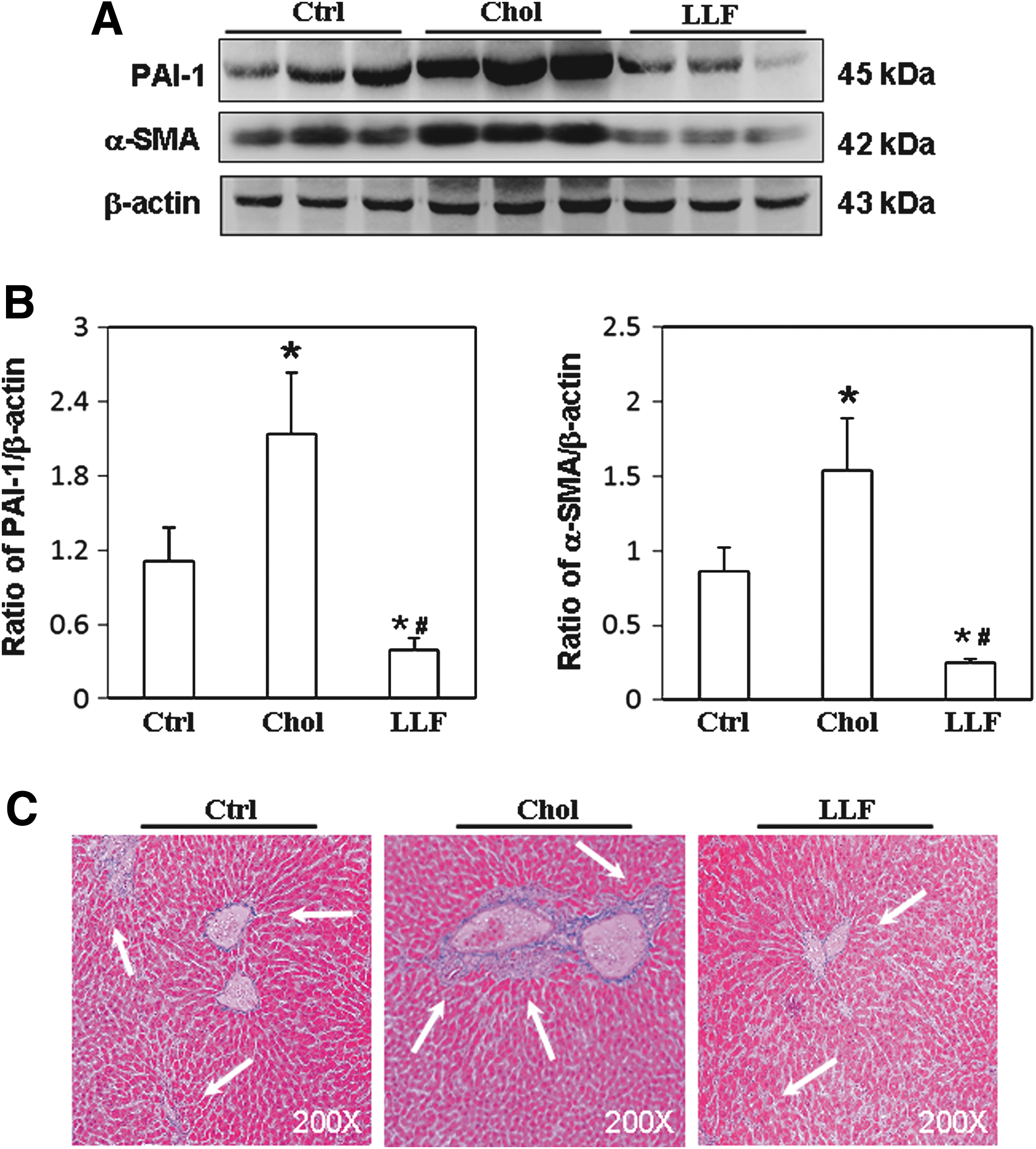
Expression of PAI-1 and α-SMA. Liver lysates obtained from the NZB/W F1 mice receiving control, high-cholesterol, and low-dose LF diet were probed with antibodies against
Discussion
Evidence reveals that oxidative stress is significantly higher in patients with SLE. Accordingly, significantly lower antioxidant enzyme activities and increased lipid peroxidation are detected in SLE patients and these effects are positively correlated with the disease activity. 28 Decreased levels of antioxidant molecules, such as GSH and DPPH, and an elevated level of MDA, the major reactive aldehyde that appears during lipid peroxidation, were detected in SLE patients compared to the control cases. 22 Accumulated oxidative stress is known to form adducts with proteins and significantly enhance their immunogenicity, resulting in immune system dysregulation and various pathological processes in SLE, including the increased expression of inflammatory cytokines. 23 In this study, LF significantly increased the activity of antioxidant molecules, including GSH and DPPH, and reduced the generation of MDA in both the serum and the liver of NZB/W F1 mice.
In addition, as demonstrated in our previous studies, a high-cholesterol diet not only induces the early onset of autoimmunity but also exacerbates disease activities in lupus-prone mice. 29,30 These studies suggest a pathological role of high cholesterol in hepatic fibrosis in SLE. In recent years, LF has attracted increasing interest owing to its protective effects against a variety of diseases, including SLE. 20,21 This study revealed that LF significantly increased antioxidative activities and attenuated fibrotic cytokines and signaling in the livers of NZB/W F1 mice that had received a high-cholesterol diet. These findings demonstrate an attenuating effect of LF on cholesterol-aggravated hepatic inflammation and fibrosis in NZB/W F1 mice. Although the mechanism requires further investigation, LF is considered to ameliorate the oxidative stress in SLE.
Liver injuries result in the recruitment of immune cells and the induction of inflammatory cytokines such as TGF-β1, TNF-α, IL-1β, IL-6, IL-10, and interferon-γ, leading to the regeneration of liver tissue and the deposition of the extracellular matrix by activated hepatic stellate cells. 31 The most profibrotic cytokine, TGF-β1, stimulates its own production by myofibroblasts and establishes an autocrine cycle of myofibroblast differentiation and activation. 32 TNF-α and IL-1β are also well-known pro-inflammatory cytokines that play a critical role in pulmonary fibrogenesis. 33,34 The gelatinases, including MMP-2 and MMP-9, participate in the PAI-1-regulated fibrinolytic activities toward basement membrane components, laminin and type IV collagen. 35 These processes lead to the deposition of an excess of extracellular matrix proteins and increase the distortion of normal liver architecture. Although LF can ameliorate liver fibrosis in dimethylnitrosamine-treated rats and lung fibrosis in mice that are exposed to hyperoxia, 36,37 little is known about the beneficial effects of LF on the liver fibrosis in SLE. This study is the first to report that LF ameliorates liver fibrosis, by decreasing gelatinase activity and fibrotic cytokines, and decreases hepatic damage indices, such as AST and ALT, in lupus-prone mice, which suggest that LF has potential as a remedy for treating hepatic inflammation and fibrosis in SLE.
LF is widely present in colostrum and milk and has a wide range of functions, including ion metabolism, antimicrobial, antioxidation, and anticancer functions, and immune modulation. LF binds iron in two distinct domains and apparently leads inactivation of these atoms in the redox cycling and hydroperoxide decomposition. 38 Although the precise mechanism needs further investigation, we speculate that the antioxidant activity of LF is probably through its regulatory effects in redox cycling and hydroperoxide decomposition. As presented in Figure 6, LF significantly increases antioxidant activities and suppresses hepatic fibrosis by decreasing the expression of fibrotic cytokines and the activation of Smad2/3, causing p-Smad2/3 to cooperate with Smad4 and SP-1 and to induce fibrosis-related proteins, including PAI-1 and α-SMA. This study is the first to demonstrate an ameliorative effect of LF on hepatic fibrosis in NZB/W F1 mice, by the inhibition of TGF-β/Smad fibrotic signaling, and suggests a therapeutic potential of LF against hepatic fibrosis in SLE.
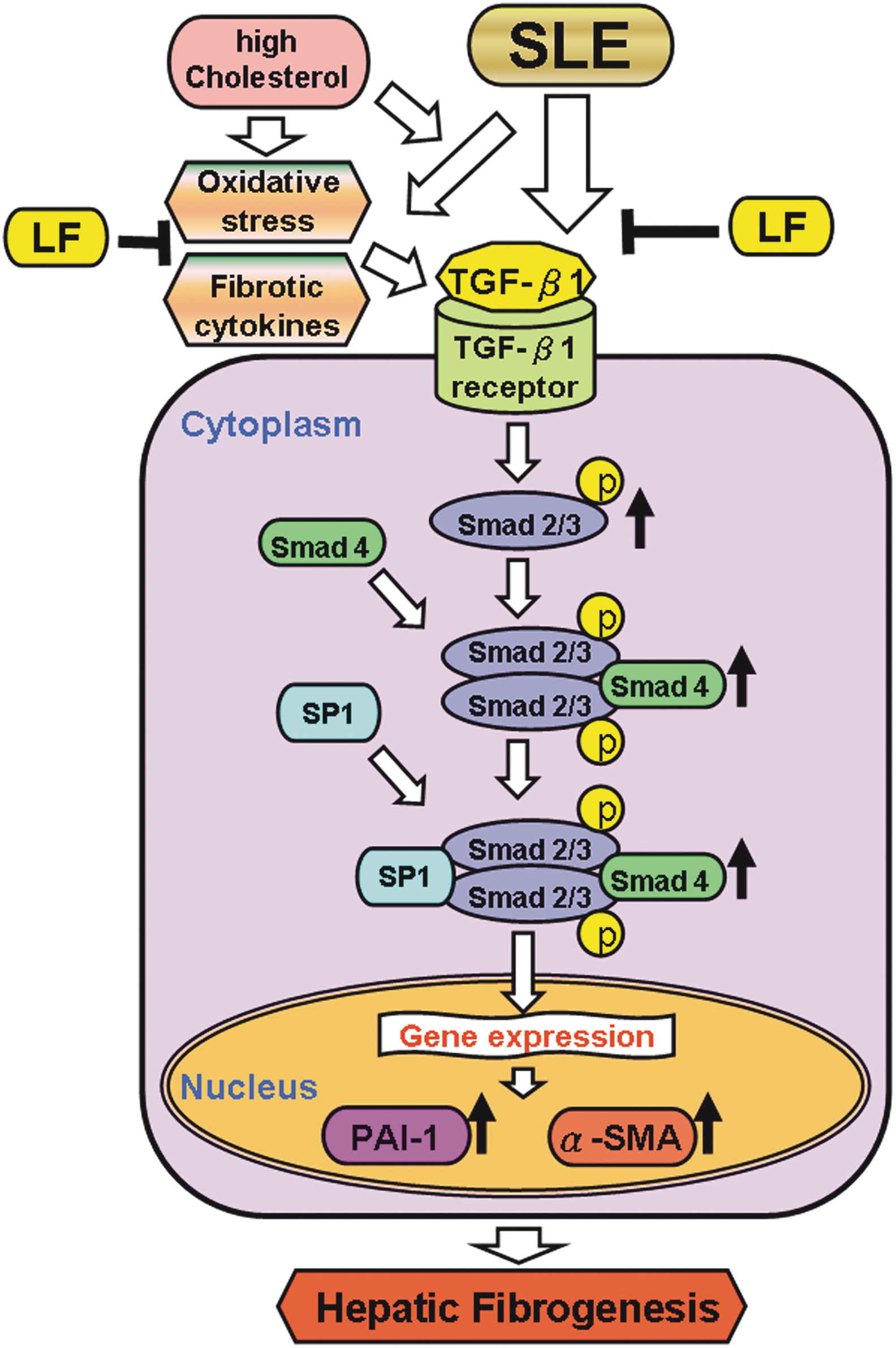
Schematic illustration of the possible mechanism of LF involved in the SLE-associated hepatic fibrosis. LF ameliorates oxidative stress, fibrotic cytokines, and TGF-β fibrosis signaling in livers of NZB/W F1 mice. SLE, systematic lupus erythematosus. Color images available online at
Footnotes
Acknowledgments
This work was funded by the Chung Shan Medical University and Chi-Mei Medical Center cooperative project (CSMU-CMMC-103-01 [CMCSMU10302]) and, in part, by the National Science Council grants, NSC-99-2632-B-040-007-MY3, from the National Science Council, Taiwan, Republic of China. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the article.
Author Disclosure Statement
No competing financial interests exist.