
Abstract
High blood pressure has been recognized as one of the main risk factors of cardiometabolic syndrome by health organizations. Hypertension is medicated by various classes of synthetic drugs; however, adverse effects have repeatedly been reported. Moreover, natural alternatives such as fish peptides have been effective in the treatment and prevention of hypertension. The aim of our study was to fractionate and identify antihypertensive peptides. Fractions were produced using different techniques of solid-phase extraction (SPE), pressure-driven ultrafiltration (UF), and electro04dialysis with UF membrane. According to our results, the hydrophobic fraction of SPE (IC50 5 μg) was the most potent anti-angiotensin converting enzyme (ACE) product. Findings of the study suggest that the separation technique plays an important role in the isolation efficiency of antihypertensive biopeptides. Importantly, the hydrophobic fraction's activity was retained through a static model of an in vitro gastrointestinal digestion system. In conclusion, polarity regardless of charge and size was the most important factor for anti-ACE activity of an Atlantic mackerel biopeptide. In addition, the presence of leucine at either of the extremities (C- or N-terminal) and/or leucine-rich motifs could well explain the hypotensive effect of our active fraction.
INTRODUCTION
According to the national cholesterol education program adult treatment panel (NCEP-ATP III) and the World Health Organization (WHO), hypertension, affecting almost one in four of the developed countries' population, is a criterion for the occurrence of metabolic syndrome and cardiometabolic syndrome. It is noteworthy that blood pressure is also an element of an individual's total cardiovascular risk. 1,2 Hypertension is strongly associated with the functional and structural cardiac and vascular deformities, thus being a significant cause of severe organ damage (heart, kidneys, brain, vasculature, and other organs) increasing the premature morbidity and mortality rate of individuals. 1 Even though angiotensin converting enzyme (ACE) (AKA dipeptidyl carboxypeptidase I and/or kininase II) regulates blood pressure, the state of prehypertension is an inflammatory-associated condition that would instigate a more severe condition if not prevented and treated by proper means in a timely manner. 3
ACE is a fairly large protein that converts angiotensin-I to angiotensin-II (a vasoconstrictor, in general, impairing endothelial function and renal hemodynamics) stimulating the release of aldosterone, thus increasing the pressure of blood initiating vascular and cardiac hypertrophy. 4 Many classes of drugs including ACE inhibitors, first introduced in 1981, have been in practice for the alleviation and deterrence of hypertension and the subsequent conditions. Yet, mild and or sever adverse effects have been reported for this group of compounds.
Studies have shown that synthetic anti-ACE compounds could efficaciously be substituted by natural biofunctional compounds in drug discoveries. Natural compounds alone or in combination therapies could noncytotoxically prevent or treat inflammation, hypertension, and ailments associated with hypertension. 5 –7 Among all compounds extracted from different terrestrial and or marine sources, fish protein and its hydrolysates have repeatedly demonstrated considerable ACE inhibitory activities in various in vitro, in vivo, and clinical studies. The topic has been thoroughly reviewed and discussed by the authors. 6 Few of the biologically active fish peptides are potent anti-ACE ingredients, as therapeutic peptides, of various commercial products intended for human consumption (e.g., Tensideal®, Molval®, Valtyron®, and PeptACE™ from mackerel, blue ling, sardine, and bonito, respectively). 8,9
Although not definite, low molecular mass and the presence of hydrophobic amino acids (HAA) and charged amino acids (CAA) at the C- or N-terminal and/or the penultimate position of those terminals would generally translate into potency of a fish biopeptide against high blood pressure and ACE activity. The presence of HAA in the sequence is a somewhat vital feature of a hypotensive fish biopeptide, yet its absence does not necessitate the ineffective activity of a protein similar to the concept of small molecular weight. For instance, many rather potent large hypotensive peptides (>1000 Da) have been successfully tested in various experimental settings as discussed in our recently published review article. These oligopeptides, originating from various fish types, have demonstrated considerable hypotensive in vitro and in vivo effects (salmon [FEDYVPLSCF], tilapia [KPLLLMQLLLLFR], tuna [GDLGKTTTVSNWSPPKYKDTP], bonito [IVGRPRHQG], bigeye tuna [WPEAAELMMEVDP], yellowfin sole [MIFPGAGGPEL], and pacific cod [LLMLDNDLPP]). 6
Antihypertensive peptides have successfully been extracted and separated by some common processes and techniques such as enzymatic hydrolysis, filtration, and chromatography. 6 Ultrafiltration (UF) membranes, with varying molecular weight cutoffs (MWCO) ranging from 1000 to 30,000 Da, have been used for rather selective separation of smaller size oligopeptides, which often exhibit higher bioactivities compared with the larger ones. Commonly for the purpose of better separation and isolation, fractions are processed by one or more chromatographic techniques post-UF. For instance, gel filtration and ion-exchange chromatography have been used for the separation of <1000 Da oligopeptides and charged peptides, respectively. Along the typical ones, some atypical procedures are also in use for the preparation of fish hypotensive biopeptides. For example, isoelectric focusing electrophoresis has been used by Auwal et al. for the separation of anti-ACE hepta-, hexa-, and penta-peptides from the hydrolysate of stone fish (Actinopyga lecanora). 10
On the one hand, filtration, with low selectivity, in recent years has been used for inexpensive, commercial, and large-scale separation of peptides and value-added products from a variety of crude hydrolysates. On the other hand, highly selective and expensive chromatography techniques have been used for profitable separation of, generally, in-demand therapeutic and pharmaceutical-grade peptides from hydrolysates. Yet, either of the abovementioned conventional separation techniques, compared with processes such as electrodialysis with UF membrane (EDUF), has obvious shortcomings limiting their practice for biopeptide production in pilot and industrial scales. Nonetheless, EDUF could reasonably and selectively isolate biopeptides (based on charge and size) from a range of lysates. 6 To the knowledge of authors, EDUF has not been yet used for the separation of charged anti-ACE fish, specifically mackerel biopeptides. 6
In our previous article, we have tested and reported the immunomodulatory effects of fractions produced from Atlantic mackerel (Scomber scombrus) protamex hydrolysate by EDUF, UF, and solid-phase extraction (SPE) based on their different properties of molecular weight, charge, and hydrophobicity. Our observations were indicative of the anti-inflammatory and proinflammatory effects of SPE and EDUF (particularly pH3 recovered fractions), respectively. 11 We, in continuation of the previous article, for the first time intended to screen EDUF and UF fractions along with SPE fractions for the separation of gastrointestinal digestion stable ACE inhibitory peptides.
The objectives of our study were (1) screening all the resultant fractions of EDUF, UF, and SPE for their hypotensive effects in an in vitro setting, (2) examining the susceptibility of the selected fraction to digestion in a static model for bioavailability prediction of the Atlantic mackerel's antihypertensive biopeptide in a hostile environment of the intestine, and (3) lastly, characterizing the selected fraction for structure–activity relationship-based assumptions.
MATERIALS AND METHODS
Materials
Chemicals and consumables
Chemicals and consumables were obtained from various producers and providers as follows.
Protamex from Novozymes A/S (Bagsværd, Kobenhavn, Denmark), column cartridge Sep-Pak® Vac 20 cc (5 g) and 6-aminoquinolyl-N-hydroxysuccinimidyl carbamate (AQC) from Waters Corp. (Milford, MA, USA), enalapril, N-hippuryl-His-Leu hydrate (HHL) 99%, ACE from rabbit lung, ≥2.0 U/mg protein (modified Warburg-Christian), 1,4-dioxane anhydrous (99.8%), bile extract porcine, pancreatin from porcine pancreas (8 × US pharmacopeia), intestinal acetone powders from rat, pepsin from porcine gastric mucosa ≥2500 U/mg protein, and trypsin from bovine pancreas from Sigma-Aldrich (Oakville, ON, Canada), hydrochloric acid (HCl) and Alfa Aesar™ cyanuric chloride (NCCl)₃ 98% from Thermo Fisher Scientific (Montreal, QC, Canada), sodium chloride crystalline (NaCl), sodium hydroxide pastilles (NaOH), and orthoboric acid (H3BO3) ACS from VWR life sciences (Montreal, QC, Canada), monopotassium phosphate (KH2PO4), sodium bicarbonate (NaHCO3), and potassium chloride (KCl) from EMD Millipore Sigma (Etobicoke, ON, Canada), multipurpose-type ED cell (100-cm2 of effective area) from Electrocell AB (Taby, Sweden), Neosepta CMX-SB (cationic) and AMX-SB (anionic) membranes from Astom Ltd. (Tokyo, Japan), polyethersulfone 4200-cm2 surface area spiral-wound 1000 Da MWCO and ultrafiltration 20,000 Da MWCO membranes from Synder Filtration (Vacaville, CA, USA).
Hydrolysate
Hydrolysate was prepared according to the method of Beaulieu et al., which has been detailed in our previous work. 14 Our material was provided by Merinov, the Quebec Fisheries, and Aquaculture Innovation Centre (Gaspé, QC, Canada). In brief, Atlantic mackerel was ground and subsequently processed by protamex for enzymatic hydrolysis. The material was then centrifuged and membrane filtered. Resultant fraction weighed <10,000 Da. 11
Fractionation methods
Fractionation by SPE
The SPE technique was used to produce relatively hydrophobic peptides according to the method of Offret et al. with modifications. 12 Briefly, 12.5 g of the mackerel hydrolysate was thoroughly dissolved in 50 mL of deionized water, under constant agitation, and loaded onto column cartridge (Sep-Pak C18, 5 g sorbent, 55–105 μm particle size) set on Waters multiposition cartridge manifold. Sample solution was eluted out of a column in the increasing acetonitrile (ACN) order of 10%, 55%, and 70% and fully dried using SpeedVac (Thermo Scientific Savant, Fisher, MA, USA), at 45°C under 12 mmHg pressure, and freeze drier. Dried samples were stored at −30°C.
Fractionation by pressure-driven UF
Mackerel hydrolysate was additionally fractionated based on the size of peptides as described in our previous work. The MWCO of the polyethersulfone spiral-wound membrane (surface area of 4200-cm2) was 1000 Da. Operational parameters were ambient room temperature, pressure of 25 psi, and 5 L of 2.4% protein feed (w/v). Recovered fractions, namely retentate (>1000 Da) and permeate (<1000 Da), were lyophilized and stored at −30°C.
Fractionation by EDUF
Mackerel hydrolysate was further processed by the electroseparation method, as described in our previous work, to produce peptides based on their charge and molecular weight. We made use of multiple membranes, a cation exchange membrane, an anion exchange membrane, and two cellulose acetate UF membranes (MWCO 20,000 Da) in this process to simultaneously generate positively and negatively charged fractions (<20,000 Da). Effect of pH was simultaneously assessed on the separation efficiency of these types of biopeptides.
Operational parameters were as follows: 4% protein feed solution (2 L), two 2 g of KCl L−1 (4 L) as the recovery compartment solutions and 20 g of Na2SO4 L−1 as the electrode rinsing solution (2 L), flow rate of 2 L min−1 for feed and KCl recovery compartments and 4 L min−1 for electrode rinsing solution, pressure of 2 psi in recovery and feed solutions, anode/cathode voltage difference of 15 V, run time of 3 h, pH of 3, 9, and/or 6, and ambient room temperature. Nine fractions were collected, freeze-dried, and stored at −30°C.
Characterization of newly generated mackerel hydrolysate SPE subfractions
Total nitrogen and protein content
The nitrogen and protein contents of material were evaluated by combustion method using a Leco Truspec Micro CHNS microanalysis analyzer. Protein contents of the samples were calculated based on the protein factor of 6.25 and the total nitrogen, organic and inorganic, of fractions.
Total amino acid analysis
Fractions were tested for total amino acid quantity according to a previously standardized method. 13,14 Samples were hydrolyzed and derivatized preanalysis. Analysis was done using a reversed-phase ultraperformance liquid chromatography (RP-UPLC) system with AccQ-Tag™ Ultra C18 column.
RP-UPLC and mass spectrometry analyses
We performed RP-UPLC analyses by a 1290 Infinity II UPLC system (Agilent Technologies, Santa Clara, CA, USA) with binary pump (G7120A), multisampler (G7167B), in-line degasser, and variable wavelength detector (VWD G7114B) adjusted to 214 nm. We diluted the fraction to 0.5% protein and filtered through a 0.22-μm polyvinylidene difluoride filter into a glass vial. Subsequently, the sample was loaded (10 μL) onto a C18 column (Acquity UPLC CSH 130 1.7-μm, 2.1 × 150 mm i.d.; Waters Corp.). We operated the column at a flow rate of 400 μL min−1, 45°C. A linear gradient consisting of solvent A (LC-MS-grade water with 0.1% formic acid) and solvent B (LC-MS-grade ACN with 0.1% formic acid) was applied, with solvent B going from 35% to 90% in 30 min, holding for 5 min, and then back to initial conditions.
We used a hybrid ion mobility quadrupole time of flight (TOF) mass spectrometer (6560 high-definition mass spectrometry [IM-Q-TOF]; Agilent, Santa Clara, USA) to detect and measure the relative abundances of peptides. All liquid chromatography with tandem mass spectrometry (LC-MS/MS) experiments were acquired using Q-TOF. Signals were recorded in a positive mode at an extended dynamic range, 2 GHz, 3200 m/z with a scan range between 100 and 3200 m/z. Nitrogen was used as drying gas (13.0 L min−1, 150°C) and as nebulizer gas at 30 psig. The voltages were set at 3500, 300, and 400 V for capillary, nozzle, and fragmentor, respectively. The instrument was calibrated using an ESI-L low-concentration tuning mix (Agilent Technologies). Data acquisition and analysis were completed by Agilent mass hunter software package (LC/MS data acquisition, version B.08.00 and qualitative analysis for IM-MS, version B.07.00 service pack 2 with BioConfirm software). Spectrum mill MS proteomics workbench Rev B.05.00.180 was used for additional search.
For identification of the fraction, we looked into various protein databases (mackerel, S. scombrus, pelagic fish, and Percomorphaceae).
ACE inhibition
In vitro antihypertensive activity of all the recovered fractions was analyzed using a spectrophotometry method previously developed by Hayakari et al., using HHL as a substrate and a colorimetric reagent of hippuric acid (HA), namely 2,4,6-trichloro-S-triazine (TT). 15 Briefly, 20 μL of ACE (250 mU mL−1 in borate buffer) was added to 20 μL of sample, followed by the addition of 80 μL of phosphate buffer and incubation of 10 min at 37°C (water bath). One replicate of each sample, concurrently, was incubated for 10 min at 95°C to inactivate ACE at the base (negative control). Postincubation, 40 μL of the ACE substrate (HHL 6.25 mM in borate buffer) was added to each test tube, followed by a 60-min incubation at 37°C.
To inactivate the further action of ACE, test tubes were incubated for 10 min at 95°C. For yellow color development, 480 μL of phosphate buffer and 360 μL of TT reagent were carefully added to all the test tubes. Two hundred microliters of each tube was pipetted into the wells of a 96-well microplate and the absorbances were read at 382 nm by a microplate reader. An ACE inhibitor drug, enalapril, was used as positive control. The percentage of ACE inhibition was calculated and documented.
Stability of selected ACE inhibitory fraction
Effect of gastric and duodenal enzymes (noncontinuous)
Stability of the selected fraction to gastrointestinal enzymes was tested according to a previously described method of Enari et al., with modifications. 16 Sample (14 mg) was individually incubated with the enzymes (1% [w/w] pepsin in 7 mL of 0.1 M HCl-KCl solution [pH 2.0], 1% [w/w] trypsin in 7 mL of 0.1 M phosphate buffer solution [pH 7.5], and/or 1% [w/w] pancreatin in 7 mL of 0.1 M phosphate buffer solution [pH 7.5]) at 37°C for 120 min. Reaction was stopped by heating at 95°C for 15 min. Throughout the process, a shaking water bath of 50 RPM was utilized for incubation of the sample. For adjusting the pH of pepsin solution, NaOH (2 N) was used. Samples were tested for anti-ACE activity according to the method described above.
Effect of in vitro gastrointestinal-simulated digestion (continuous)
The stability of selected anti-ACE fraction was challenged by an in vitro simulated gastrointestinal digestion static model, according to a previously described method with modifications. 17 Briefly, the sample (20 mg) was dissolved in 10 mL of phosphate buffer (10 mM), pH 6.8, and incubated for 2 min at 37°C (oral condition). Right after, acidic condition (pH 1.5–2) was produced by addition of 5 mL of HCl-KCl (1 M) to the solution mixture. To mimic gastric conditions, we used pepsin 1% (w/w) (substrate: enzyme ratio 100:1) in 5 mL of HCl-KCl (1 M) solution and incubated the sample solution at 37°C for 60 min (gastric condition). Ten milliliters of NaHCO3 (1 M, pH 7.5) was added for pH adjustment.
Duodenal condition was simulated by the addition of 10 mL of mixture solution of 1% (w/w) pancreatin, 1% (w/w) trypsin, and 35% (w/w) bile extract in phosphate buffer (10 mM, pH 8.2) (duodenal condition) with a final pH of 7.5. After 30 min of incubation at 37°C, 1% (w/w) intestinal acetone powder solution in phosphate buffer (10 mM, pH 8.2) was added to mirror the intestinal condition. Solution was incubated for another 30 min at 37°C. Reaction was stopped by heating the solution (95°C) for 15 min.
Throughout the process, a shaking water bath of 50 RPM was utilized for incubation of the sample. Control was prepared by mixing the inactivated enzymes (by heating at 95°C for 15 min) with the peptide solution. Samples were drawn (200 μL) and centrifuged (2000 g for 30 sec) from each condition to perform activity tests. ACE inhibition analysis was performed as described above. The experiments were performed in triplicates.
Statistical analyses
All experiments had a completely randomized design. Collected data were tested for normal distribution by the Anderson–Darling normality test and analyzed by one way-ANOVA, using Minitab (version 18). Data in tables are presented as mean ± standard deviation. The error bars on the graphs represent the standard deviation of two or three independent studies. Different letters on bars, as obtained by Tukey's multiple comparison test at 95% confidence intervals (P < .05), were used to indicate statistical significance among the different groups.
RESULTS AND DISCUSSION
Isolation efficiency
Isolation efficiency of the SPE process is noted in Table 1. SPE yielded about 4% to 13% of crude fractions with gradient hydrophobicities. The highest and lowest yields, irrespective of the bioactivity concept, were from 10%ACN and 70%ACN, correspondingly (Table 1). Fractions were 86%–91% protein rich. Regarding the EDUF process, as reported in our previous work, most of the <20,000 Da positively charged peptides, weight-based, were produced in EDUF pH3 (3.1% of the initial hydrolysate feed) followed by pH6 and pH9. Accordingly, most of the <20,000 Da negatively charged peptides were obtained from EDUF pH6 and pH9 (1.2%–1.3% of the initial hydrolysate feed) followed by pH3. Regardless of the peptides' property and bioactivity, about 22% (of the initial feed) <1000 Da oligopeptides were recovered in the pressure-driven UF process.
Solid-Phase Extraction Process Yield Based on Protein Dry Weight
Protein percentages of the SPE fractions were measured by Leco and MicroLeco instruments. Protein contents of each fractions (%) are dry weight based. Recovery percentages of the fractions are based on the starting and final material's protein weight. Each cell represents mean of three measurements ± standard deviation. Data were normally distributed according to the Anderson–Darling normality test.
Columns with different superscripts letters, as obtained by ANOVA (Tukey's multiple comparison test), are used to indicate statistical significance (P ≤ .05).
ACN, acetonitrile; SPE, solid-phase extraction.
In our previous work, we concluded that EDUF and SPE processes were the techniques of choice for separation of immunomodulatory peptides, pro- and anti-inflammatory fractions, respectively.
Characterization of selected fractions
Total amino acid analysis
Results of the unprocessed hydrolysate and SPE fractions are presented in Table 2. Total amino acid composition of EDUF and UF fractions were discussed in our earlier research work. Rationally, SPE-70%ACN and SPE-10%ACN ought to be the most and least HAA-rich fractions. In accord, SPE-70%ACN had 33% more total HAA than its most hydrophilic counterpart fraction, SPE-10%ACN. Nonetheless, the same fraction had 25% less CAA than the SPE-10%ACN. Interestingly, most hydrophobic fractions had about approximately twofold the branched chain amino acids (BCAA) compared with SPE-10%ACN. As presented in the following section, we observed noticeable anti-ACE potency of the unprocessed mackerel hydrolysate (IC50 of 8.8 μg of protein) compared with the hydrophobic fraction (IC50 of 5.0 μg of protein). Protamex-digested Atlantic mackerel hydrolysate had about 81% more CAA, and 33% and 46% less HAA and BCAA than SPE-70%ACN, respectively. Yet, its IC50 value, statistically, was only marginally different from the hydrophobic fraction.
Total Amino Acid Composition (g 100 g− 1 of Protein) of Mackerel Hydrolysate and Solid-Phase Extraction Fractions
Total amino acid contents of the mackerel hydrolysate and the SPE fractions were measured by the RP-UPLC system and AccQ-Tag™ Ultra C18 column. Each cell represents the mean of observations ± standard deviation of duplicate measurements. Data were normally distributed according to the Anderson–Darling normality test.
Columns with different superscripts letters, as obtained by ANOVA (Tukey's multiple comparison test), are used to indicate statistical significance (P ≤ .05).
Membrane-filtered hydrolysate.
SPE fractions in the increasing order of hydrophobicity.
BCAA, branched chain amino acids; CAA, charged amino acids; EAA, essential amino acids; HAA, hydrophobic amino acids; RP-UPLC, reversed-phase ultraperformance liquid chromatography.
Among charged EDUF fractions, cationic and particularly anionic peptides of pH9 were the most potent ones with IC50 values of 11.5 and 9.4 μg of protein, yet the effects could not exceed the hydrolysate's activity. The anionic peptide fraction of pH9, as presented in our earlier work, was relatively richer in BCAA, HAA, and essential amino acids (EAA), while it only composed of balanced non-EAA and CAA compared with its counterpart fraction of pH3 and pH6. Cationic fraction of pH9 had a balanced amount of EAA, non-EAA, and BCAA although it contained less of total HAA compared with cationic peptide fractions of pH3 and pH6. The hydrolysate of grass carp with substantial in vitro and in vivo antihypertension activity was rich in negatively CAA. 18 It is of note that positively charged peptides of various origins, alone or in combination therapies, could significantly attenuate hypertension. 19 –22
Conclusively, aside from compositional and structural properties of anti-ACE fish biopeptides, total CAA content of the fraction could possibly be as important as its HAA content for the hypotensive effects of Atlantic mackerel extracted peptides.
RP-UPLC and mass spectrometry analyses
All the possible sequences identified and predicted in the selected fraction of SPE-70%ACN have positive GRAVY values up to 4.5, with relatively small molecular sizes, demonstrating and confirming the hydrophobic character of the highly potent hypotensive peptidic material (Table 3 and Fig. 1). These peptide sequences originated from 19 different precursor proteins such as NADH and formate dehydrogenase, nucleoside and ABC transporter permease, ATPase, argininosuccinate lyase, S-malonyl transferase, transcriptional regulator, and cytochrome b. An antimicrobial subfraction of the same fraction recently has been characterized by our team and the potent material was characteristically leucine and isoleucine rich (LILLILLLLKLLLLLI, LLILLLLKLLLLLI, and LLILLLLLLILLLILLPF). 12 Fragments of these oligopeptides have also been identified in the selected hypotensive fraction.
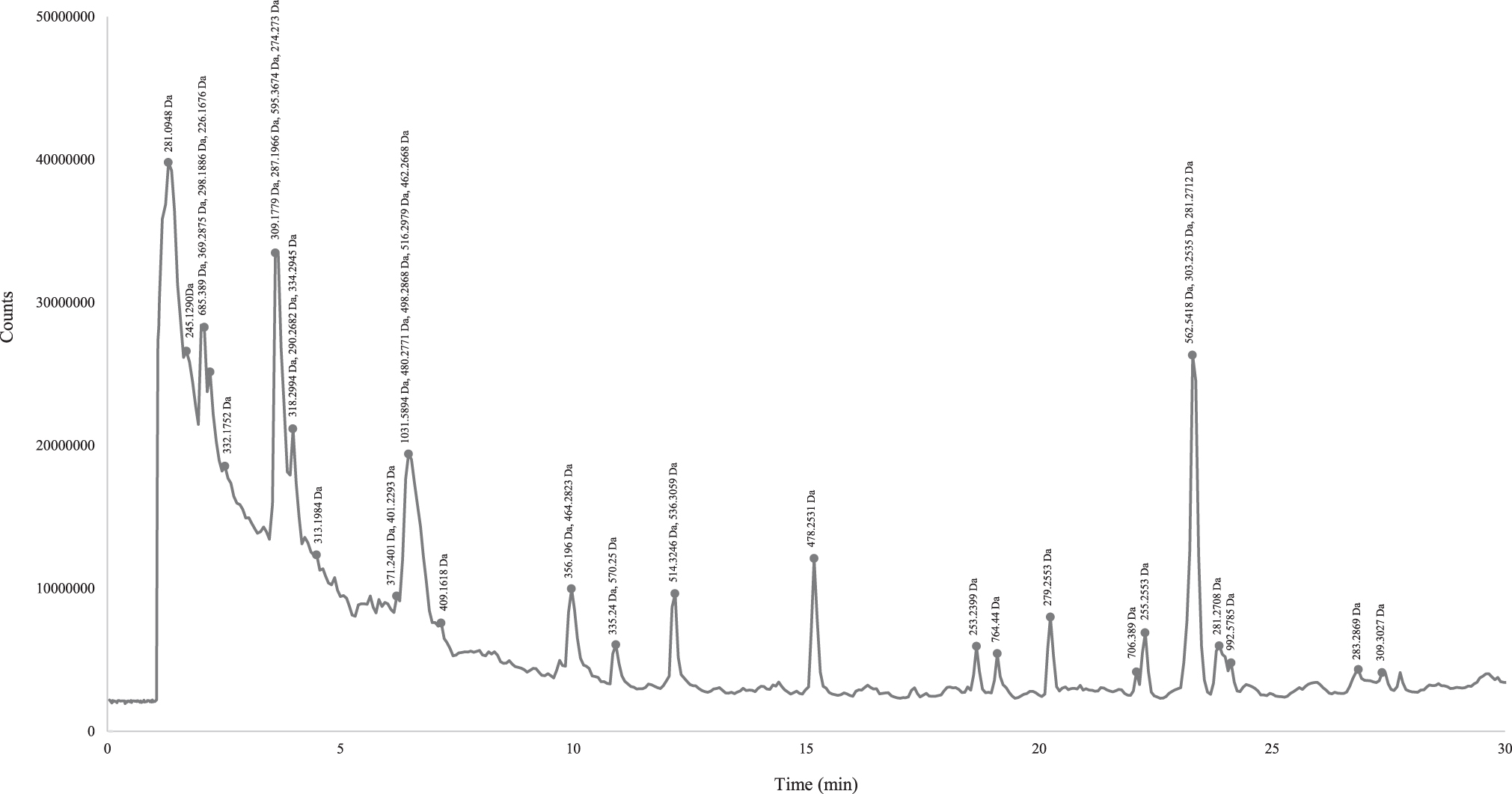
Chromatogram of SPE-70%ACN fraction and compounds' molecular weight associated with each peak. ACN, acetonitrile; SPE, solid-phase extraction.
Properties of Major Oligopeptides of Solid-Phase Extraction-70%ACN Fraction
The fraction was analyzed by RP-UPLC, and MS. Spectrum Mill MS Proteomics Workbench Rev B.05.00.180 was used for additional search. Each row consists of properties of a predicted sequence based on various protein databases (mackerel, Scomber scombrus, pelagic fish, and Percomorphaceae).
Decades ago, the presence of leucine and isoleucine at either of the extremities of an antihypertensive sequence has been stated vital for the bioactivity of a peptide. 23 Fish hypotensive peptides such as AHLLLL, LGPLGHQ, LNLQDFR, MVGSAPGVL, LLMLDNDLPP, ELLGFV, QDLLFR, MILLLFR, NELLLFR, and NLGALLFR are evidently leucine rich. 6 The antihypertensive dipeptides such as IF, FL, and VL, specifically of fish origin, are also fragments of some possible sequences of the fraction meeting the Lipinski's rule of five, thus being potential drug-like candidates (Table 3). 6 Moreover, examining the data about the motifs of possible sequences (submitted as Supplementary Data), we noticed that there are no shared sequences between the most and the least effective anti-ACE fractions (SPE hydrophobic and pH3 negatively charged EDUF fractions, respectively).
Interestingly, FPF (homologous with motifs of cytochrome b (mitochondrion) [Notacanthus chemnitzii] and ABC transporter permease [Cedecea neteri]) are a shared possible sequence between hydrophobic and pH9 negatively charged EDUF fractions. The latter was the most potent hypotensive fraction among EDUF separated material. The tripeptide, FPF, has been previously reported for its anti-ACE and anti-dipeptidyl peptidase 4 (DPP-IV) activities. 24,25 The peptide according to the study of Liu et al. has three potential binding sites to DPP-IV enzyme (Arg125, Glu205, Glu206). 26 Motifs of this tripeptide, FP, and PF, have been part of many biopeptides with a wide range of bioactivities, as discussed in our review articles. 5 Information regarding pH3 negatively charged and pH9 negatively charged EDUF fractions has been submitted as Supplementary Data to our previous research article.
ACE inhibitory activities of fractions
Results of anti-ACE potency of all the fractions and their corresponding IC50 values are presented in Table 4 and Figure 2. Anti-ACE effects of enalapril were logarithmic. Inhibition activity of the samples was rather logarithmic, and the effects for some of the tested fractions were slightly dose independent (Fig. 1). The most hydrophobic fraction, in comparison with the most hydrophilic one, was about three-times more potent. Even though the most hydrophobic fraction of SPE had the smallest IC50 value among all in this study, overall, it was statistically identical to its midhydrophobic (55%ACN) counterpart (Table 4). Least inhibitory fractions were negatively charged EDUF pH3 and negatively charged EDUF pH6 peptides being almost five- and eightfold less effective than the hydrolysate and SPE-separated hydrophobic material.
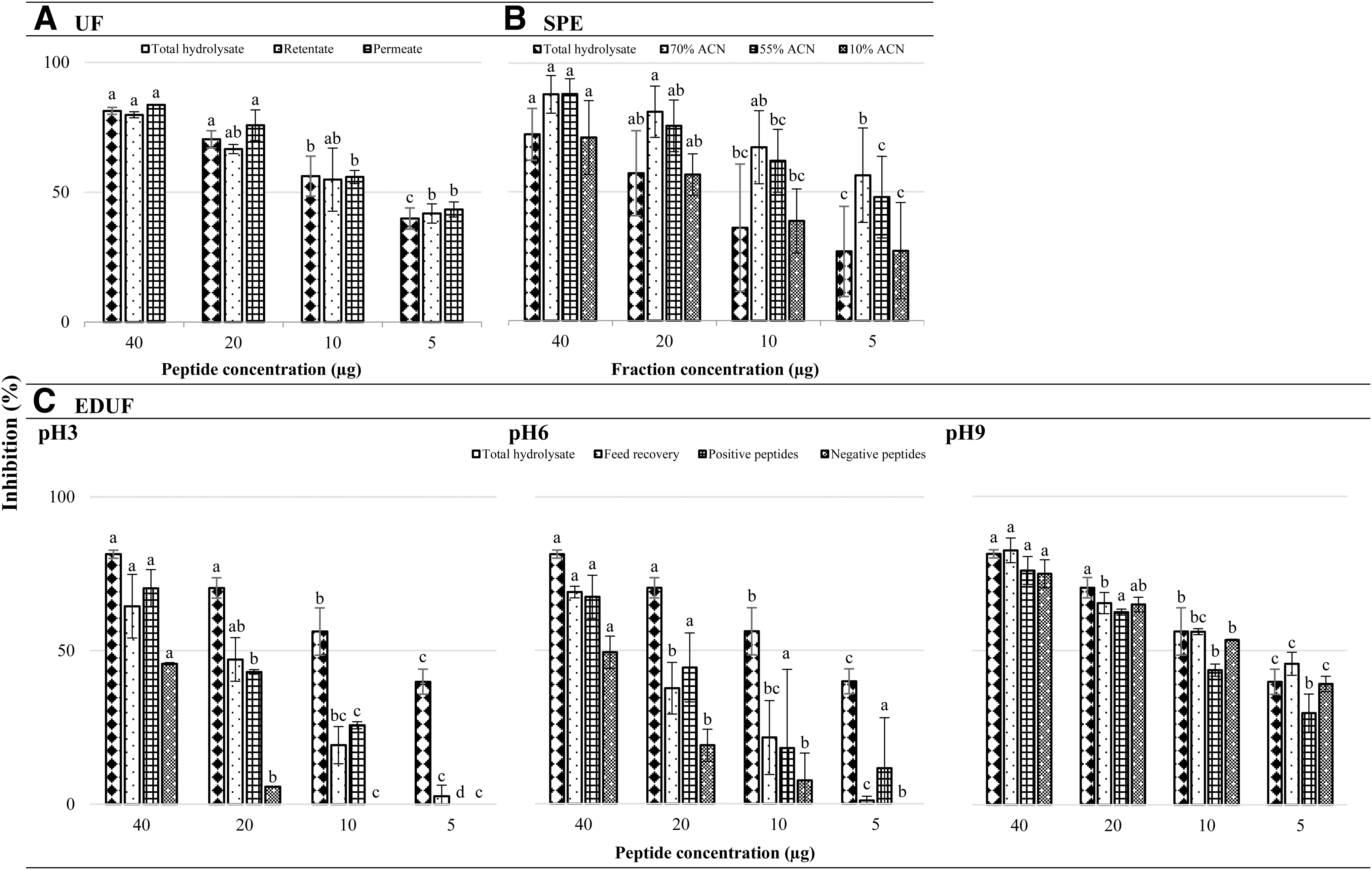
Percent in vitro ACE inhibitory activity of four different concentrations (μg of protein or fraction) of mackerel hydrolysate and its fractions
In Vitro Angiotensin Converting Enzyme
ACE inhibition potential of Atlantic mackerel hydrolysate and its fractions was measured by a spectrophotometry method (using a substrate: hippuryl-histidyl-leucine and a colorimetric reagent of hippuric acid: 2,4,6-trichloro-S-triazine). IC50 column represents the mean of observations ± standard deviation of two to six independent studies, triplicate measurements. Data were normally distributed according to the Anderson–Darling normality test.
Columns with different superscript letters, as obtained by ANOVA (Tukey's multiple comparison test), are used to indicate statistical significance (P ≤ .05).
ACE, angiotensin converting enzyme; EDUF, electrodialysis with UF membrane; UF, ultrafiltration.
In general, the EDUF technique and the implemented operational parameters were not as successful in the generation of anti-ACE peptides. However, notably the condition of pH9 (IC50 8.9–11.5 μg of protein) statistically did not decrease the efficacy of hydrolysate (IC50 8.8 μg of protein). Regarding the production of small-molecular mass oligopeptides, although size-based fractionation of hydrolysate did not improve the effectivity of material, yet comparably did not reduce the potency of fractions. These observations highlight the fact that size and charge of peptides may not be as vital for anti-ACE properties of mackerel-derived fractions. However, hydrophobicity seemed to play a rather significant role in the hypotensive effects of the sample. In accordance, hydrophobicity was likewise essential for the in vitro anti-inflammatory effect of 70%ACN fraction. The presence of HAA in the sequence of a peptide has repeatedly been documented as relatively important for the effective interaction of biopeptide with the active sites of ACE. 6
Interestingly, a fairly hydrophobic derivative oligopeptide of 70%ACN fraction, KVEIVAINDPFIDL (mass of 1584 Da, and hydrophobicity of +15.86 kcal mol−1), has previously demonstrated sa ignificant antibacterial activity against a range of bacteria, yet most effective on Listeria spp. with the minimum inhibitory concentration value of 207.5 μg mL−1. 12 Previous studies have shown the importance of C-terminal hydrophobic moiety for anti-inflammatory activity of natural antimicrobial peptides. 27 Finally, hydrophobicity is somewhat essential for the anti-inflammatory and antihypertensive activity of a peptide.
Stability of the selected hypotensive fraction to gastrointestinal digestion, continuous and noncontinuous
The most potent fraction (SPE-70%ACN fraction) was assessed for its susceptibility to gastrointestinal digestion, and the results of material stability are presented in Tables 5 and 6. Bioactivity of the test material was retained in both simulated continuous (through oral, gastric, duodenal, and intestinal phase) and noncontinuous digestion assays and the variation was not statistically significant compared with the undigested sample. In the continuous model, at the end of digestion, both digested and nondigested (control) fraction inhibited about 50% of the ACE activity. It was also evident that none of the digestive enzymes (pepsin, trypsin, and/or pancreatin) could further hydrolyze the fraction to smaller oligopeptides. A sample size of 40 μg fraction (34.3 μg of hydrophobic protein) postdigestive enzyme hydrolysis could quench the activity of 5 mU ACE by about 90%.
Stability of Solid-Phase Extraction-70%ACN Fraction to Gastrointestinal Proteases
The sensitivity of SPE-70%ACN fraction to three different gastrointestinal enzymes was tested. Final peptide concentration in the well was 34.3 μg. ACE inhibition column in the table presents the percentage inhibitory effect of digested fraction against ACE. Undigested fraction's anti-ACE activity (as control) was insignificantly different from the enzyme-digested samples. Table represents the mean of observations ± standard deviation of three independent studies, triplicate measurements. Data were normally distributed according to the Anderson–Darling normality test.
Columns with superscript letter, as obtained by ANOVA (Tukey's multiple comparison test), are used to indicate statistical significance (P ≤ .05).
Stability of Solid-Phase Extraction-70%ACN Fraction to In Vitro Continuous Gastrointestinal Digestion
The sensitivity of SPE-70%ACN fraction to gastrointestinal digestion (representative of digestion in mouth, stomach, and small and large intestines) was tested using a static model. The table represents the mean of observations ± standard deviation of three independent studies, triplicate measurements. ACE inhibition activity of both digested and control samples did not significantly differ from one another. Data were normally distributed according to the Anderson–Darling normality test.
Columns with superscript letter, as obtained by ANOVA (Tukey's multiple comparison test), are used to indicate statistical significance (P ≤ .05).
Sample solution subjected to gastrointestinal digestion.
Sample solution prepared with inactivated enzymes simultaneously tested with the main sample (as control).
Comparable results were documented for the nondigested hydrophobic fraction. In addition, in many studies, gastrointestinal (GI) digestion has been shown to improve the activity and increase the potency of peptides, yet such result was not achieved in our study. For instance, further hydrolysis of cobia hydrolysate with digestive enzymes enhanced its antihypertensive activity up to 61%. 28 It is of note that in the study of Offret et al., Atlantic mackerel GAPDH-related antimicrobial peptide (a derivative of the same fraction) lost its integrity and activity postgastric phase in the dynamic GI TIM-1 model, while SPE-70%ACN fraction was digestion resistant in a simulated static GI model. 12 It has been revealed that the presence of proline and tryptophan, generally at the C-terminus, is required for the stability of a fish peptide to GI digestion and digestive proteases, including pepsin, trypsin, and chymotrypsin. 6,16
To the knowledge of authors, a comparable finding about the GI stability of Atlantic mackerel-extracted hypotensive biopeptide has not yet been documented to date and we are the first to report such an observation. In the next phase, the authors have planned to test the in vivo effectivity of selected fractions on metabolic syndrome-associated factors. 29
CONCLUSION
In this study, of the three processes, the chromatographic technique of SPE was the most efficient practice for the separation of GI digestion-stable anti-ACE material of mackerel hydrolysate origin. Moreover, according to our results, pressure-driven UF and/or EDUF pH9, if optimized, may also be fairly suitable for the purpose. In the present work, hydrophobicity feature of the hypotensive peptides topped charge and size properties of the Atlantic mackerel fractions, thus being two- and up to eight-times more potent compared with the unprocessed hydrolysate and its processed fractions, correspondingly.
In agreement, it has previously been noted that characteristics of the final product and subsequently the activity of hypotensive fish biopeptides not only depend on the nature of digestion but also on the type of the isolation/purification technique. The highly potent hydrophobic fraction, identified in this study, has the capacity for further purification for the isolation of drug-like substances that may be used as drug leads in a drug design and discoveries for antihypertensive medication. Moreover, in-depth examinations of the effect of fraction in a well-controlled cell, and in vivo animal models or human intervention studies concomitant with structure–activity relationship experiments are required for definite conclusions.
Footnotes
ACKNOWLEDGMENTS
The authors would like to thank Louis-Charles Rainville and Nadine Renaud (Merinov, the Quebec Fisheries, and Aquaculture Innovation Centre [Gaspé, QC, Canada]) for providing the Atlantic mackerel hydrolysate and for their technical support.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This project was made possible by the Natural Sciences and Engineering Research Council of Canada (NSERC) under the name of Strategic Partnership Grants for Project grant STPGP/479527 – 2015.
SUPPLEMENTARY MATERIAL
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.