
Abstract
The mechanisms of action responsible for the reported hypolipidemic activity of barley sprouts have yet to be elucidated. The objective of this study was to compare the content of saponarin (the sole flavonoid present in barley sprout leaves), hypolipidemic activity between barley sprout water extract (BSW) and barley sprout ethanol extract (BSE), and the associated relevance to hypolipidemic activity in 3T3-L1 preadipocytes. BSW elicited superior antiadipogenic effects when compared with BSE in MDI mixture [IBMX 0.5 mM + dexamethasone 1 μM + insulin 1 μg/mL]-treated 3T3-L1 preadipocytes. BSW attenuated MDI-mediated triacylglycerol (TAG) accumulation by inhibiting fatty acid synthase (FAS). FAS protein expression was markedly and dose dependently attenuated by BSW, with higher doses suppressing expression to a level equivalent to the controls. BSW also significantly attenuated MDI-mediated increases in the expression of genes involved in TAG synthesis as well as FAS in 3T3-L1 preadipocytes. High-performance liquid chromatography analysis indicated that BSW contains more than four times more saponarin than BSE. Further investigation of saponarin-mediated hypotriacylglycerolemic activity and related gene expression revealed that saponarin significantly inhibited TAG accumulation, which was attributed to reductions in TAG synthesis-related gene expression. Taken together, these findings provide a basis for further development of barley sprout extract for functional health food purposes.
INTRODUCTION
Increasing rates of obesity remain a significant health problem worldwide. Obesity is linked to higher susceptibility to hypertension, diabetes, hyperlipidemia, nonalcoholic fatty liver disease, and other metabolic disorders. 1 Once obesity is reached, lasting weight loss strategies generally have a low success rate. 2 Therefore, prevention is thought to be a more desirable approach to control.
As a young vegetable, sprouts are defined as being <10 cm in length, having 1–3 leaves after sprouting from seeds. 3 Interestingly, sprouting leads to remarkable increases in nutrients, including vitamins A, B, C, and E, minerals, 4 and phenolic compounds. 5 In addition, sprouts during the beginning of their growth phase contain even higher concentrations of nutrients. 6,7 Due to the higher content of nutrients and phenolic compounds, sprouts exhibit properties beneficial to human health. For instance, sprouts from Brassica oleracea species such as cabbage, kale, broccoli, and brussels have antioxidant properties, 8 while pseudocereals, amaranth, quinoa and buckwheat, alfalfa and soya bean seeds also exhibit antioxidant activity attributable to higher (increased) total phenolic content during the sprouting process. 4
Barley sprout (Hordeum vulgare L.) is defined as a young green leaf growing from a barley seed. Barley sprout consumed as a tea has received significant attention in recent years as a functional food material in several countries. 9 Germination generally causes improved protein content and digestion, increases sugar and B vitamin content, and decreases levels of phytates and protease inhibitors, 10 which is associated with a greater extent of activity of endogenous hydrolytic activity, 11 providing improved functionality compared with the seed form of the plant. Barley sprout extract exhibits antioxidant effects, which have been demonstrated in an alcohol-induced oxidative liver injury mouse model 12 by increasing NRF2 activity. In addition, barley sprout has been reported to have hypolipidemic, antidepressant, and antidiabetic effects. 13
The hypolipidemic activity of barley sprout has not been elucidated in terms of a focus on the potential mechanism mediated by triacylglycerol (TAG) inhibition, which is one of the major mechanisms involved in the development of obesity and related metabolic diseases, including nonalcoholic fatty liver disease. 1 The objectives of this study were to (1) compare the antiadipogenic activity of ethanol extract and water extract of barley sprout and examine related mechanisms of action, (2) compare content of the major flavonoid saponarin between two extracts, and (3) examine the hypotriacylglycerolemic activity of saponarin using 3T3-L1 preadipocytes as an adipogenesis model.
MATERIALS AND METHODS
Materials
Bovine calf serum (BCS), dimethyl sulfoxide (DMSO), saponarin, 3-isobutyl-1-methylxanthine (IBMX), dexamethasone, insulin, thiazolyl blue tetrazolium bromide (MTT), and Oil Red O (ORO) were purchased from Sigma-Aldrich (St. Louis, MO, USA). BCS, fetal bovine serum (FBS), Dulbecco's modified Eagle's medium (high glucose), Dulbecco's phosphate-buffered saline (DPBS), and antibiotic–antimycotic solution were purchased from Welgene, Inc. (Gyeongsan, Gyeongsangbuk-do, Republic of Korea). High-performance liquid chromatography (HPLC) grade water, acetonitrile, and acetic acid were purchased from Avantor Performance Materials (Center Valley, PA, USA).
Barley sprout extraction
Barley sprouts were obtained from Chamchaewon (Jincheon, Chungbuk, Republic of Korea). The barley sprouts were washed, and 1 kg was mixed with 70% ethanol or pure water. For ethanol extraction, the mixture was extracted for 2 h (vacuum-enrichment extraction, 70°C), and filtered by filter press followed by decompression concentration. For water extraction, the mixture was extracted for 2 h (vacuum-enrichment extraction, 95°C) and centrifuged (8000 g, 30 min). Both concentrates were then freeze dried and powdered (yield 1.8% and 1.9% for ethanol and water extraction, respectively). Both extract types were stored at −20°C until analysis.
Cell culture, treatment, and MTT assay
Mouse 3T3-L1 preadipocytes were purchased from ATCC (Manassas, VA, USA). Cells (7 × 104) were cultured in DMEM with high glucose with 10% BCS +1% antibiotics, and placed into an incubator (37°C, 5% CO2). After reaching 70–80% confluence, the cells were seeded into 96-well plates.
When confluence reached 100%, they were treated with either DMEM with high glucose medium +10% FBS +1% antibiotics or MDI mixture [IBMX 0.5 mM + dexamethasone 1 μM + insulin 1 μg/mL] added with or without sample extracts (barley sprout ethanol extract; BSE, barley sprout water extract; BSW, or saponarin). After 48 h incubation, the medium was changed to new medium containing insulin (1 μg/mL) only. After the incubation, the medium was again changed to insulin-containing fresh media, followed by a further 48-h incubation.
After observing the morphology of the adipocytes through a microscope, an MTT assay was performed. Medium was removed and washed with DPBS. MTT solution (0.5 mg/mL) was added to each well, and incubated at 37°C, 5% CO2 incubator for 3 h in the absence of light. The medium was then removed, DMSO was added to each well, and the plate was kept at room temperature with rocking for 5 min to produce formazan followed by reading the optical density (OD) value at 570 nm with a microplate reader (Molecular Devices, Sunnyvale, CA, USA).
Oil Red O staining
After reaching 70–80% confluence, the cells were seeded into 12-well plates, and incubated for 24 h. The 3T3-L1 cells were then treated with either DMEM with high glucose medium +10% FBS +1% antibiotics or MDI mixture (IBMX 0.5 mM + dexamethasone 1 μM + insulin 1 μg/mL) added with or without samples (BSE, BSW, or saponarin). After 48 h, the medium was changed to medium containing insulin (1 μg/mL) only.
After 48 h incubation, the medium was again changed to fresh media containing insulin alone, before incubation for another 48 h. Cells were then washed with DPBS and incubated with 3.7% formalin for 1 h, followed by washing and incubating at 60% in 2-propanol for 5 min. Cells were then stained with ORO solution for 20 min, followed by elution with 2-propanol, and absorbance (492 nm) of the dark-red elutes was measured (Molecular Devices).
Triacylglycerol
Cells were cultured and treated with the MDI mixture and insulin in the presence or absence of BSW (100 and 200 μg/mL) or saponarin (50 and 100 μM) as described in the previous section. On the last day, medium was collected to measure TAG content using a commercially available kit (Abcam, Cambridge, United Kingdom).
The experiment was performed according to the manufacturer's protocol. In brief, lipase was added to the collected medium and incubated for 20 min. Reaction mixture was added and incubated for another 60 min before OD was measured at 570 nm. TAG in the samples was quantified against a standard curve and normalized to total protein quantity, which was measured using a commercially available kit (Intron Biotechnology, Seongnam, Republic of Korea).
Western blot
Cells were treated as described above. After reaching confluence, the cells were treated with MDI complex in the presence or absence of 100 or 200 μg/mL BSW. Medium was changed into insulin-containing medium twice over a 2-day interval. Cells were downed, and the pellets were collected. Ripa buffer (Cell Signaling Technology, Danvers, MA, USA) was added for lysis. After centrifugation (20,000 g, 20 min, 4°C), the upper layer was collected and total protein was measured. Total protein (15 μg) was loaded onto 10% gels, before electrophoresis and transferred onto a nitrocellulose membrane.
The membranes were incubated with specific antibodies against fatty acid synthase (FAS), ACLY, and PPARγ (Cell Signaling Technology), followed by secondary antibodies (Bio-Rad, Hercules, CA, USA). Protein loading was verified by probing blots with antibody against HSP90 (Santa Cruz Biotechnology, Heidelberg, Germany). FAS, ACLY, PPARγ, and HSP90 proteins were detected using enhanced chemiluminescence substrate (Thermo Scientific, Rockford, IL, USA).
Quantitative real-time PCR
Cells were cultured and treated as described above. At day 6, the cells were washed with DPBS two times, and cell pellets were collected after centrifugation (2,000 g, 3 min). To extract total RNA, the cells were lysed with TRIzol reagent (Takara, Shiga, Japan), an equal volume of chloroform (EMD Millipore, Burlington, MA, USA) was added, and the upper layer was collected. Isopropanol was added to precipitate RNA. Reverse transcription for cDNA synthesis was performed using a cDNA reverse transcription kit (Takara Bio). Quantitative real-time PCR analysis was performed as described. 14 Primers were designed according to the GenBank database (Table 1). GAPDH was used to normalize the data.
Primer Sequences for Quantitative Real-Time Polymerase Chain Reaction
Saponarin analysis
Saponarin content of barley sprout extract was determined using HPLC according to the method described by Jin et al. (2011) with some modifications. 15 The barley sprout extract sample (∼400 mg) was added to a 50 mL conical tube containing 40 mL of DMSO and mixed using a vortex mixer for 2 min at room temperature. The sample was centrifuged (5000 g, 15 min) and filtered through a PVDF syringe filter (0.45 μm, Woongki Science, Seoul, Korea). A Waters HPLC System (Waters, Milford, MA, USA), equipped with an e2695 Separations Module and a 2998 photodiode array detector, was used for the analysis.
Saponarin was separated on a Capcell Pak C18 MG2 column (250 × 4.6 mm i.d., Shiseido, Tokyo, Japan) maintained at 40°C. Optimal separation was achieved by gradient elution using mobile phase (A) acetic acid/water (0.8:100, v/v) and (B) acetonitrile at a flow rate of 0.6 mL/min. The gradients (time, %B) were set as (0 min, 10), (2 min, 10), (30 min, 24), (60 min, 76), and (80 min, 10). The detector monitored the eluent at 336 nm, which is the wavelength of maximum absorption for saponarin.
Saponarin was identified by comparing its retention time (∼19.0 min) with that of the reference standard. The standard solutions at concentrations of 25–100 mg/L for calibration were prepared from a stock solution of 1000 mg/L by dilution. Each measurement was performed in triplicate. Saponarin content was calculated as mg per kg of barley sprout extract.
Statistical analysis
Data (mean ± SD) were analyzed using GraphPad Prism software (La Jolla, CA, USA). Student's t-test was used to compare group mean differences. Analysis was considered statistically significant at P < .05.
RESULTS AND DISCUSSION
Nontoxic concentrations of BSW are more effective than BSE at inhibiting lipid accumulation in 3T3-L1 preadipocytes treated with MDI
The MTT assays were performed using ethanol (BSE) and water (BSW) extracts of barley sprout in MDI-treated 3T3-L1 preadipocytes. Cells were exposed to different concentrations of BSE or BSW up to 400 μg/mL. While BSE decreased cell viability to <90% at 200 μg/mL, BSW did not affect cell viability up to 400 μg/mL (Fig. 1A). BSE beyond 200 μg/mL treatment reduced the viability of the cells, indicating that high-dose BSE may be cytotoxic to MDI-treated 3T3-L1 adipocytes.
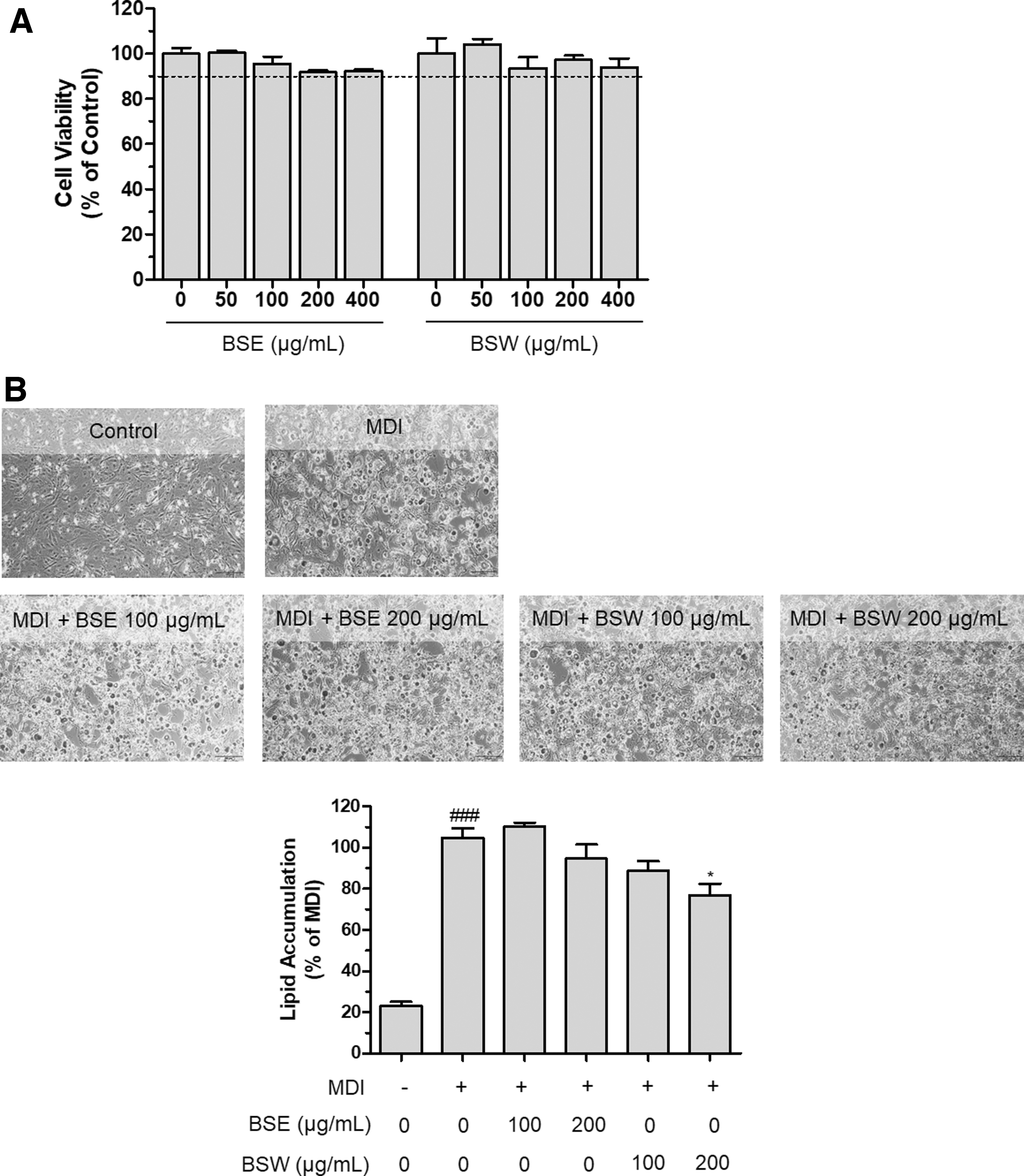
Cell viability was examined
We sought to examine the effect of BSE and BSW on adipogenesis. BSE and BSW were added to the medium every 2 days during differentiation of the 3T3-L1 preadipocytes, and the cells were stained with ORO on day 6 of differentiation. Stained cells were microscopically observed and quantified after dissolving the cells in 2-propanol. Compared with the non-MDI-treated controls, MDI-treated 3T3-L1 preadipocytes exhibited markedly increased lipid accumulation, which was unaffected by BSE, but reduced by BSW treatment (Fig. 1B, upper). After quantification of total lipids in these cells, MDI-mediated lipid accumulation was determined to be significantly reduced by BSW but not by BSE (Fig. 1B, lower). BSW at the high dose (200 μg/mL) significantly attenuated MDI-mediated lipid accumulation in 3T3-L1 preadipocytes.
Various health-promoting properties of barley sprout have been reported. For instance, barley sprouts have antioxidant and anti-inflammatory activities, which have been attributed to their high contents of total phenols, chlorophyll, and carotenoids. 11 Anti-inflammatory effects of BSE extract have also been reported in an alcohol-induced steatosis mouse model through reduction of hepatic TNF-α and increases in hepatic glutathione levels. 16
Meanwhile, few studies have focused on the antiadipogenic effects of barley sprouts. A recent study conducted by Kim et al. (2020) compared the adipogenic activity of BSE extract depending on germination conditions (light and nutrient treatment) and saponarin in 3T3-L1 adipocytes and HepG2 cells. 17 However, the mechanism by which the antiadipogenic activity of the extract and saponarin suppresses adipogenesis in the two different cell models was not examined. 17
BSW markedly attenuates FAS protein expression in MDI-treated 3T3-L1 preadipocytes
To further examine the antiadipogenic activity of BSW alone, the expression of adipogenesis-related proteins was measured by western blot. BSW reduced FAS protein expression that was otherwise increased by MDI treatment in 3T3-L1 preadipocytes (Fig. 2A). Whereas ACLY was likely attenuated by BSW, PPARγ was unchanged by BSW in these cells (Fig. 2A).
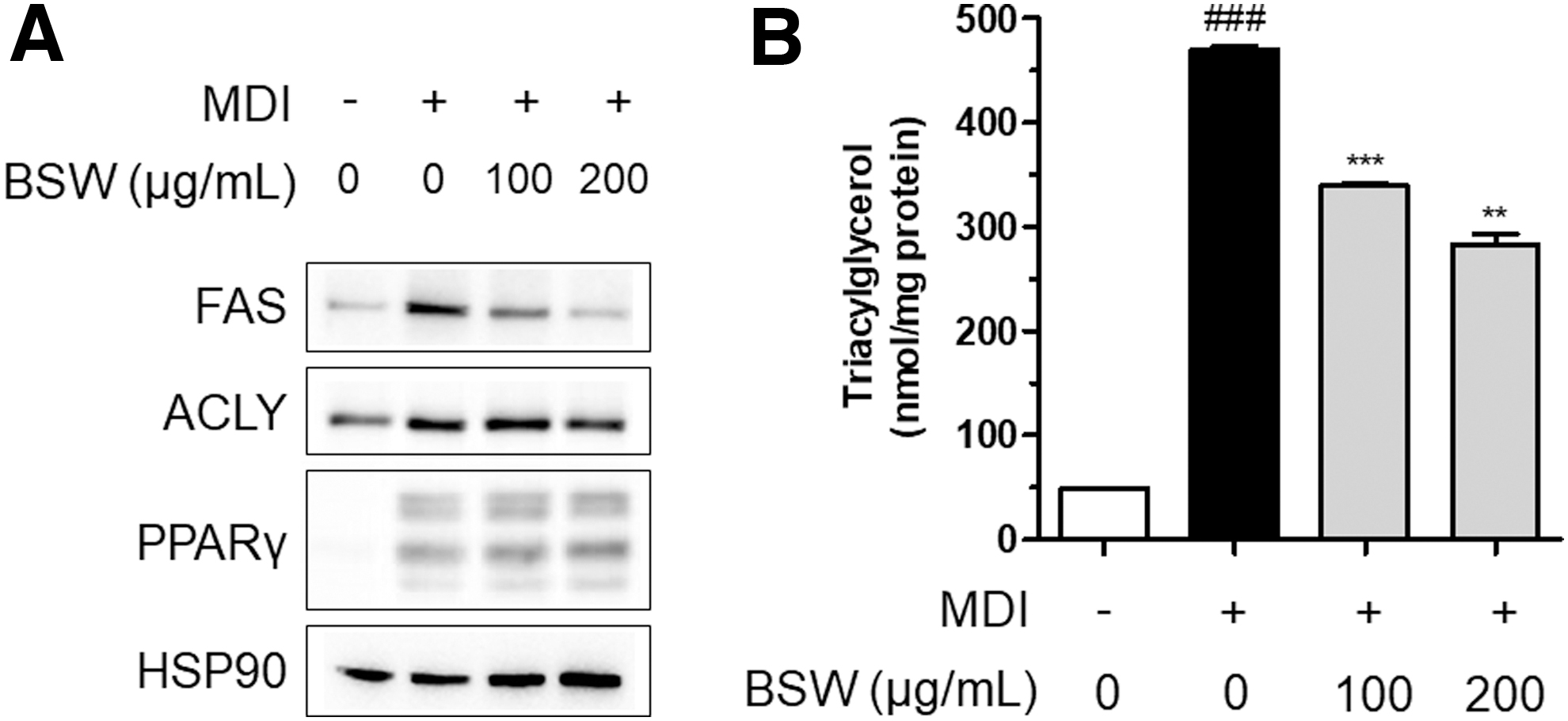
Expression of proteins including FAS, ACLY, PPARγ, and HSP90
TAG content was then measured using a commercially available kit, and the total level of TAG was normalized to the total protein content. Total TAG was significantly increased by MDI treatment, which was dose dependently and significantly attenuated by BSW treatment (P < .001; Fig. 2B). Taken together, these observations suggest that BSW-mediated reductions in lipid accumulation were attributable to reduced TAG accumulation, which was likely associated with FAS inhibition in MDI-treated 3T3-L1 preadipocytes.
FAS (encoded by Fasn) is an enzyme that catalyzes the de novo synthesis of fatty acids. The primary role of FAS involves the biosynthesis of saturated fatty acids, primarily producing palmitate, and other short fatty acids including stearate, myristate, and laureate using the substrates acetyl-CoA, malonyl-CoA, and NADPH. 18,19 FAS plays a key role in the pathogenesis of fatty liver and dyslipidemia. 20 Decreased FASN activity leads to reductions in lipid droplet size and stimulation of FFA and glycerol efflux by adipocytes. 21
Treatment with the FAS inhibitor C75 induced weight loss and improved hepatic steatosis in obese mice, 20 suggesting the possibility that FAS inhibition reduces TAG accumulation. Zhu et al. demonstrated that 22 downregulation of FASN expression using short hair-pin RNA (shRNA)-mediated RNA interference and C75 (FASN activity inhibitor) treatment both altered the expression of lipogenic genes, including GPAT AGAPT6 and DGAT2, and enhanced the expression of the lipolysis-related genes ATGL and HSL, which partly caused a reduction in TAG content. 22
BSW downregulates the expression of adipogenic genes
BSW-mediated FAS inhibition can suppress TAG accumulation, which may be related to the inhibition of TG synthesis-related gene expression in 3T3-L1 adipocytes. Hence, we aimed to measure the expression of genes related to TAG and fatty acid synthesis. FAS gene expression was significantly (P < .01) reduced by BSW at 100 and 200 μg/mL in MDI-treated 3T3-L1 preadipocytes (Fig. 3A). Another fatty acid biosynthesis gene, ACLY, that was otherwise increased by MDI treatment in 3T3-L1 preadipocytes, was also significantly reduced by BSW at 100 μg/mL (Fig. 3A).
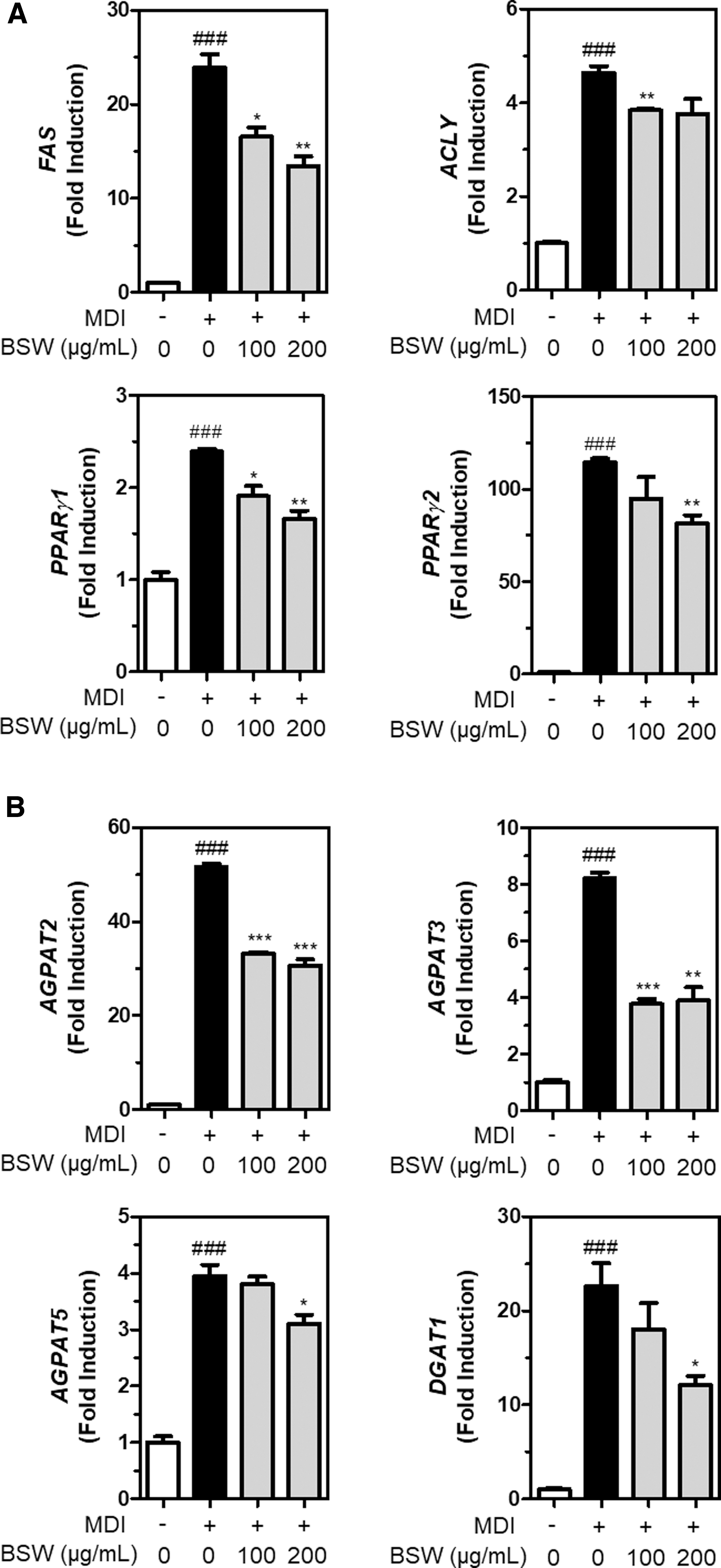
Expression of adipogenic genes including FAS, ACLY, PPARγ1, and PPARγ2
ACLY gene expression was generally reduced by BSW at 200 μg/mL, which was not statistically significant. We then examined whether BSW-mediated suppression of TAG accumulation was associated with decreased TAG biosynthesis-related gene expression. Interestingly, MDI-mediated increased expression of AGPAT2, 3, 5 and DGAT1 genes was significantly (P < .001) reduced by BSW supplementation in 3T3-L1 preadipocytes (Fig. 3B).
The synthesis and metabolism of TAG are significant in whole-body energy homeostasis in mammals, and dysregulation of TAG metabolism promotes the development of obesity, lipodystrophy, cardiovascular disease, insulin resistance, and type 2 diabetes. 23 Mature adipocytes need fatty acids and glycerol-3-phosphate for TAG synthesis and further storage. Free fatty acids that can cause steatosis originate from (1) circulating free fatty acid in the blood (59%), which is a major form of circulating free fatty acid; (2) de novo synthesis from acetyl-CoA in the liver (26%); and (3) diet (15%). 24 Meanwhile, glycerol-3-phosphate arises from glucose metabolism followed by glucose uptake from blood circulation through the glucose transporter GLUT4. 25
During TAG synthesis, the action of enzymes is important. TAG synthesis begins with the formation of lysophosphatidic acid (LPA) through acylation of the stereospecific numbering (sn)-1 position of glycerol-3-phosphate by the action of sn-glycerol-3-phosphate acyltransferase (GPAT). 26 LPA then converts to phosphatidic acid (PA) through acylation by the action of sn-1-acylglycerol-3-phosphate O-acyltransferase (AGPAT). PA converts to diacylglycerol (DAG), which then converts to form TAG by the action of diacylglycerol acyltransferase (DGAT). In this study, BSW inhibited FAS gene expression, followed by AGPAT and DGAT1 downregulation, thereby contributing to the suppression of TAG accumulation in MDI-treated 3T3-L1 adipocytes.
BSW contains relatively more saponarin than BSE
As one of the major compounds in barley sprouts, saponarin is the sole flavonoid in the primary leaves of the plant. 27 We compared the amount of saponarin in BSW and BSE by HPLC. Figure 4 showed chromatograms of saponarin, BSW and BSE. The BSW sample had more than four times the amount of saponarin than that found in BSE total extract (Table 2).
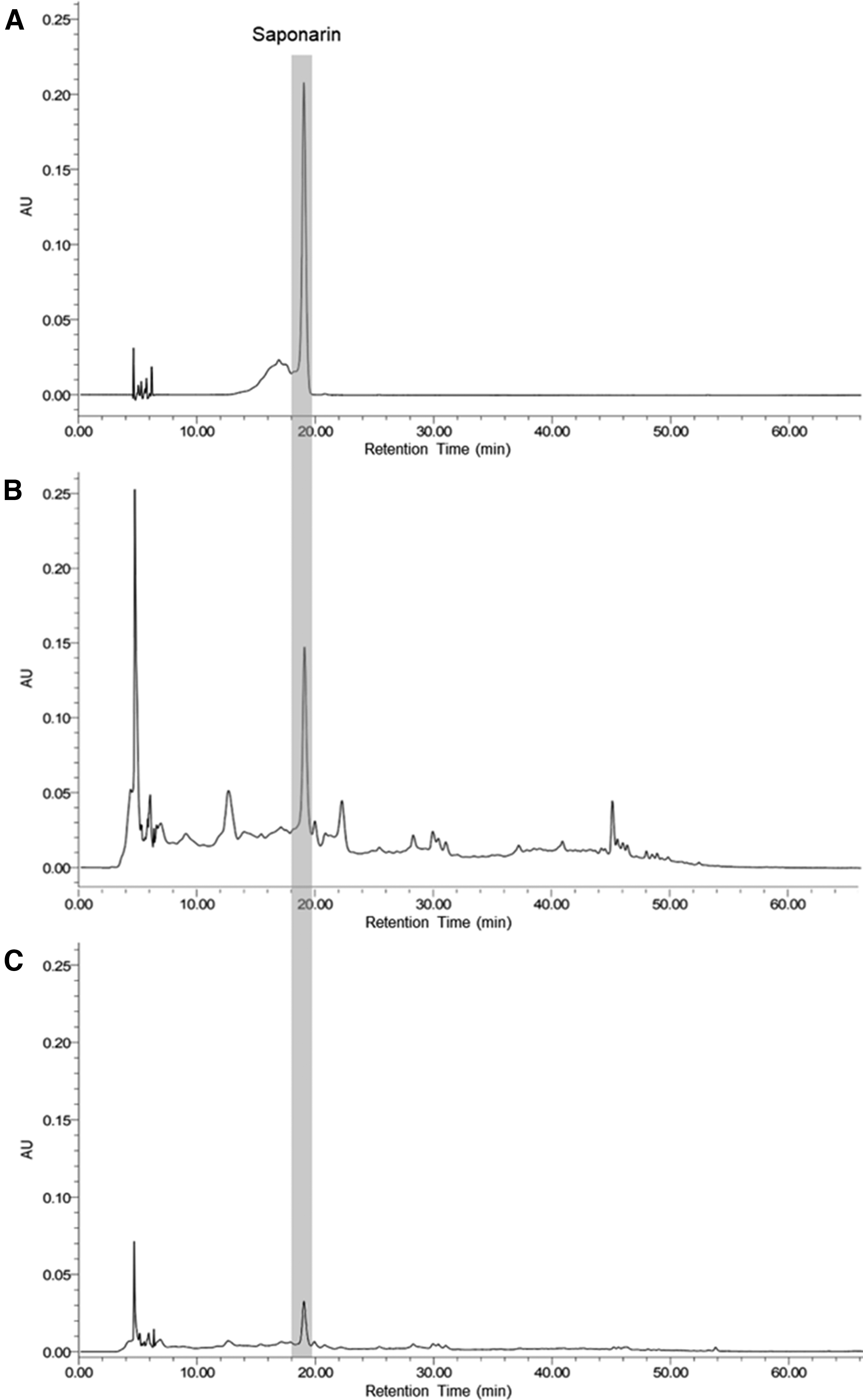
HPLC of reference standard of saponarin
Saponarin Concentration Found in Barley Sprout Water and Barley Sprout Ethanol (mg per kg Extract)
Indicates that there was a significant difference between BSW and BSE (P < .001).
BSW, barley sprout water; BSE, barley sprout ethanol.
Saponarin was first identified in barley sprout by Osawa et al., and the identification was confirmed by Markham & Mitchell. 28,29 Saponarin is abundant in barley sprout, as it mainly accumulates during the early stage of barley leaf development. 30
Barley sprouts contain nutrients and other functional compounds that can contribute to the prevention of various chronic diseases, including circulatory disorders, cancer, obesity, diabetes, arthritis, and high cholesterol, while exerting antioxidant and anti-inflammatory effects. 31 It is also abundant in nutrients and functional ingredients, including dietary fiber, protein, fat, vitamins A and C, minerals including calcium and chlorophyll, as well as the antioxidant enzymes superoxide dismutase and catalase.
In addition, barley sprouts contain flavonoids including lutonarin and saponarin as well as the functional compounds quercetin, kaempferol, catechin, and beta-glucan. 32 We found that BSW and BSE both yielded saponarin in this study. Interestingly, BSW contained a greater amount of saponarin than BSE.
Nontoxic levels of saponarin significantly inhibited lipid accumulation in MDI-treated 3T3-L1 preadipocytes
We next sought to investigate the antiadipogenic activity of saponarin. Cell viability was not affected by saponarin treatment up to 100 μM (Fig. 5A), and thus, 50 and 100 μM saponarin was added to MDI-treated 3T3-L1 preadipocytes to examine its antiadipogenic effects. MDI-mediated increases in lipid accumulation were significantly (P < .001) attenuated by saponarin supplementation (Fig. 5B, upper). ORO staining visually and statistically indicated that saponarin attenuates lipid droplet accumulation that was otherwise increased by MDI treatment (P < .0001; Fig. 5B, lower).
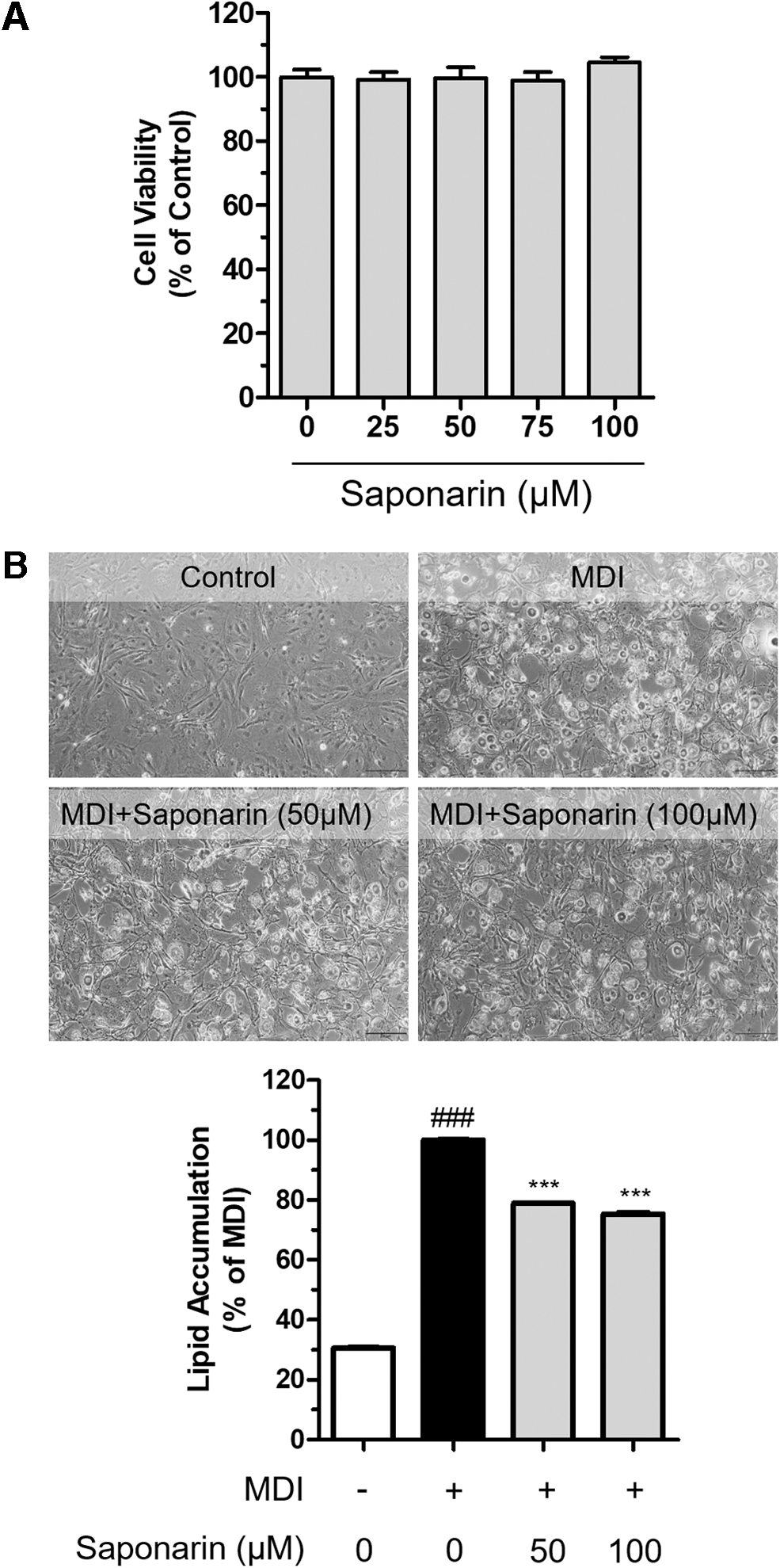
Cell viability was examined
Suppression of TAG accumulation by saponarin was associated with the downregulation of TAG synthesis-related genes
MDI-mediated increases in TAG concentration were significantly attenuated by saponarin 50 and 100 μM treatment in 3T3-L1 preadipocytes (Fig. 6A). TAG accumulation was reduced by high-dose saponarin (100 μM) to a similar extent observed in the non-MDI-treated controls (Fig. 6A). The expressions of TAG synthesis-related genes was then measured.
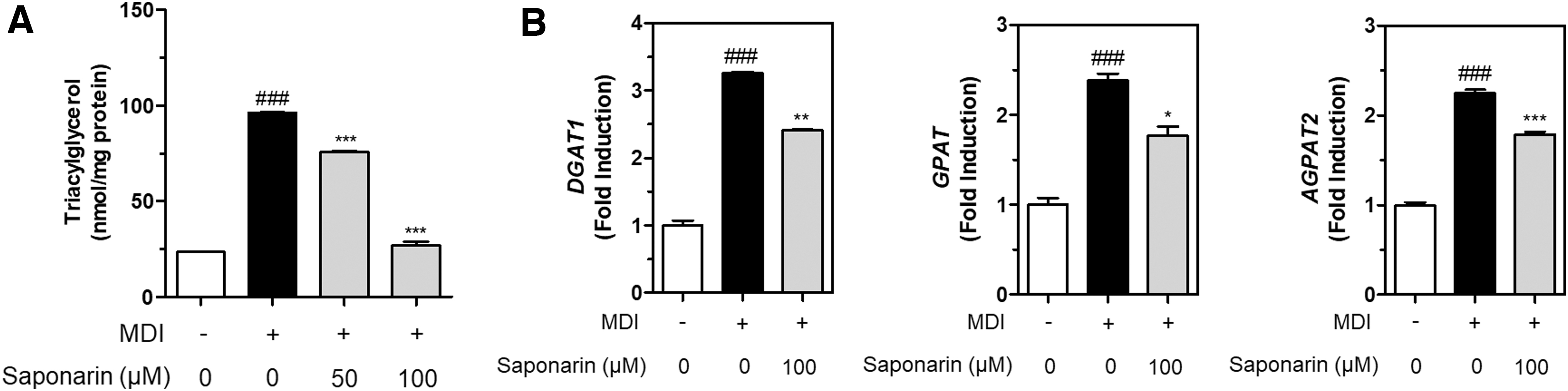
TAG concentration was measured and normalized to the total protein amount
3T3-L1 preadipocytes were treated with MDI and insulin to induce TAG accumulation, and high-dose saponarin (100 μM) significantly reduced DGAT1, GPAT, and AGPAT2 gene expression in the 3T3-L1 preadipocytes (Fig. 6B). This collectively implies that BSW mediates antiadipogenic effects, which are attributable to the suppression of TAG accumulation through the inhibition of DGAT1, GPAT, and AGPAT2 gene expression in MDI-treated 3T3-L1 adipocytes.
Saponarin (apigenin-6-C-glycosyl-7-O-glucoside) is a flavone glycoside. 27 Additional beneficial properties of saponarin relevant for human health have been reported. Saponarin regulates insulin sensitivity by suppressing gluconeogenesis and increasing glucose uptake through GLUT4 transactivation, which occurs through phosphorylation of AMPK in a calcium-dependent manner. 33
Saponarin also has anti-inflammatory activity that has been demonstrated in LPS-treated RAW264.7 cells through NFKB inactivation by inhibiting the mitogen-activated protein kinase ERK and p38 phosphorylation. 34 Saponarin in barley sprout has been suggested to be increased through blue light-mediated de novo synthesis. 17,27 Due to higher content of saponarin in BSW extract, sprouts germinated with blue light or cultivated at an early stage could be used in green-colored tea for health-promoting effects, including antiadipogenesis.
CONCLUSION
BSW exhibited greater antiadipogenic activity than BSE, which was likely attributable to the >4 times greater amount of saponarin present. We observed that BSW attenuated MDI-mediated TAG accumulation by inhibiting FAS. FAS protein expression was markedly and dose dependently attenuated by BSW treatment. Consistently, BSW significantly attenuated MDI-mediated increases in the expression of genes involved in TAG synthesis as well as FAS in 3T3-L1 preadipocytes.
Saponarin reduced TAG accumulation significantly and dose dependently, which was associated with TAG synthesis-related gene expression, including DGAT1, GPAT, and AGPAT2 in 3T3-L1 preadipocytes. Kang et al. have also demonstrated that barley (Hordeum vulgare L.) sprout young leaf water extract has antiobesity effects associated with reductions in adipogenic gene (PPARγ, FAS, C/EBPα, FABP4) expression in 3T3-L1 adipocytes as well as in diet-induced obese (DIO) mice.
However, this study has for the first time demonstrated that BSW exhibits antiadipogenic activity specifically through attenuation of TAG accumulation in 3T3-L1 adipocytes. We also investigated the hypotriacylglycerolemic activity of saponarin, the sole flavonoid present in the primary leaves of barley sprout. Despite the known hypolipidemic activity of barley sprout in vitro and in vivo, its antiobesity effects in humans remain inconclusive. 3 Further studies are warranted to examine the lipid lowering activity of barley sprouts in clinical settings.
Footnotes
AUTHOR DISCLOSURE STATEMENT
The authors have declared no conflict of interest.
FUNDING INFORMATION
This study was supported by the Main Research Program from the Korea Food Research Institute (KFRI), funded by the Ministry of Science and ICT (E0210601-02).