
Abstract
Uric acid is the final product of purine metabolism in human. The increase of serum uric acid is tightly related to the incidence of hyperuricemia and gout. Also, it has been reported that the intake of purine-rich foods like meat and seafood is associated with an increased risk of gout. Therefore, the reduction of purine absorption is one of therapeutic approaches to prevent hyperuricemia and gout. Currently, probiotics are being studied for the management of hyperuricemia and gout. In this study, we aimed to investigate the effect of Lactobacillus brevis MJM60390 on hyperuricemia induced by a high-purine diet and potassium oxonate in a mouse model. L. brevis MJM60390 among 24 lactic acid bacteria isolated from fermented foods showed the highest ability to assimilate inosine and guanosine in vitro and typical probiotic characteristics, like the absence of bioamine production, D-lactate production, hemolytic activity, as well as tolerance to simulated orogastrointestinal conditions and adherence to Caco-2 cells. In an in vivo animal study, the uric acid level in serum was significantly reduced to a normal level after oral administration of L. brevis MJM60390 for 2 weeks. The activity of xanthine oxidase catalyzing the formation of uric acid was also inhibited by 30%. Interestingly, damage to the glomerulus, Bowman's capsule, and tubules in the hyperuricemia model were reversed by supplementation with this strain. Fecal microbiome analysis revealed that L. brevis MJM60390 supplementation enhanced the relative abundance of the Rikenellaceae family, which produces the short-chain fatty acid butyrate and helps to maintain good gut condition. Therefore, these results demonstrated that L. brevis MJM60390 can be a probiotic candidate to prevent hyperuricemia.
Introduction
Uric acid is the metabolic final product of purines in the human body, and increased serum uric acid levels cause hyperuricemia and gout. 1,2 Generally, uric acid in human plasma ranges from 4 to 7 mg/dL, and concentrations above or below this range are considered abnormal and generally referred to as hyperuricemia or hypouricemia, respectively. 3 A prolonged uric acid excess at levels exceeding 7 mg/dL tends to cause a deposit of crystalline uric acid at the surface of joints or cartilage. Accumulation of urate crystals in the joints or soft tissue can cause inflammation and extreme pain, which is clinically diagnosed as gout.
The global burden of this condition is substantial and is increasing. In South Korea, the number of people with gout is increasing rapidly since 2010. According to the National Health Insurance Corporation, the number of gout patients increased by 36%, from 338,302 in 2015 to 459,429 in 2019. 4
The reasons for “gout-level” hyperuricemia are complex and poorly understood. Obesity, 5 cerebrovascular and cardiovascular diseases, 6 chronic kidney disease, 7 and nephropathy in diabetic patients 8,9 are known to be related with increased uric acid levels in the serum. Alcohol is also closely related to gout because it induces excessive synthesis of uric acid in the body and prevents it from being released into the urine. 10 In addition, the intake of various purine-rich foods, like meat and seafood, can increase the risk of gout. 11,12 Food rich in adenine and hypoxanthine significantly increased the uric acid level in serum compared with control. 13 Therefore, patients with hyperuricemia were asked to control their diet.
However, due to the limited information available regarding the correlation between different purine components and food materials, it is difficult for the patients to manage their diet. 14 Moreover, condiments, which are widely used in cooking and food processing contain purines, and the kinds and contents differ greatly. The purine content in some of these food additives ranged from 684.8 mg/100 g in fresh soup powder to 3182.70 mg/100 g in chlorella. 15
The epidemiologic studies suggested that both increased synthesis and decreased excretion of uric acid are related to hyperuricemia. 16 Therefore, the current pharmacologic agents for lowering serum uric acid are developed based on the inhibition of its synthesis, 17 augmentation of excretion, 18 or its catabolism. 19 However, all of these agents need to be taken for a long time, which increases the risk of side effects, such as abdominal pain, nausea, diarrhea, skin rash to severe hypersensitivity, and others. 20 It is necessary to find an effective, safe, and long-term strategy to control the hyperuricemia.
Probiotics are “live microorganisms that, when administered in adequate amounts, confer a health benefit on the host.” They are generally regarded as safe, and long-term use of probiotics is well accepted. 21 According to the guidelines of Korea Food and Drug Administration (KFDA), safety assessment should be carried out for the development of probiotic strains. The safety assessment includes antibiotic susceptibility, hemolytic activity, toxin and D-lactate production, etc.
The beneficial effect of probiotics include the maintenance of gut microbiota composition, modulation of immune response, management of metabolic disorders, etc. Many research studies have revealed that these beneficial effects are related to the strain itself and modulation of intestinal microbiota. 22,23 A previous study found that Lactobacillus gasseri PA3 can reduce the intestinal absorption of purine and lower uric acid levels. 24 The purines are taken up and incorporated by some lactobacilli. 25,26
In the present study, we screened and isolated a lactobacillus strain with high purine-degrading activity, and the antihyperuricemia activity was assessed in potassium oxonate-induced hyperuricemic mice supplemented with a high-purine diet (HPD). The possible mechanism for lowering serum uric acid level was also investigated. Furthermore, the probiotic activity and in vitro safety were characterized.
Materials and Methods
Screening of purine-degrading lactobacillus strains
A total of 24 lactobacillus strains were tested for their purine-degrading ability. The lactic acid bacteria (LAB) strains were inoculated into de Man, Rogosa, and Sharpe (MRS) medium (Difco, USA) and cultured at 37°C for 48 h. After incubation, 1 mL of culture broth was centrifuged at 4000 g for 10 min. After washing three times with 1 mL of 0.85% NaCl, the cells were resuspended in 375 μL of 1 mM inosine-1 mM guanosine solution, and incubated at 37°C for 120 min. After that, the solution was centrifuged at 4000 g for 10 min. Next, 270 μL of the supernatant were taken, and mixed with 30 μL of 0.1 M HClO4 solution to stop the reaction. The residual inosine and guanosine in the solution were determined using high-performance liquid chromatography (HPLC) as described below.
HPLC was performed by using a YMC ODS-A column (S-5 μm, 12 nm, 250 × 4.6 mm) connected to a binary HPLC pump (Waters 1525, Milford, MA, USA), and the column temperature was maintained at 37°C. The isocratic elution was performed with a NaClO4-H3PO4 solution (0.1 mM NaClO4, and 0.187 M H3PO4 in dH2O) at a flow rate of 1 mL/min. Inosine, guanosine, and adenosine were detected at 254 nm by the retention times of 21.8, 17.3, and 11.8 min, respectively, and were quantified by comparing with the standard.
The degradation rate and speed of guanosine or inosine by different LAB strains were calculated according to the following formula: V = (C-X)/120, D = [(C-X)/C] × 100%, where V is degradation speed (g/L/min); C denotes content of inosine or guanosine (g/L); X represents the residual content of inosine or guanosine (g/L); and D indicates the degradation rate (%).
Strain identification based on 16S rRNA sequence and phylogenetic analysis
Genomic DNA was isolated using a commercial kit (GeneAll, Republic of Korea). The 16S rRNA sequence amplified by using a universal primer, 27F and 1492R, were sequenced and blasted in the Ezbiocloud database (ChunLab, Inc., Republic of Korea). For phylogenetic analysis, the 16S rDNA sequences of closely related strains were multiple aligned, and the phylogenetic tree was constructed with neighbor-joining method using MEGA X software. 27
Safety assessment of candidate LAB in vitro
In vitro safety assessments were carried out for hemolytic activity, D-lactate production, biogenic amine production, and susceptibility to antibiotics.
Hemolytic activity of MJM60390 was tested on blood agars, which contain 5% defibrinated sheep blood (Hardy Diagnostics, A10). MJM60390 was streaked on the blood agar and incubated at 37°C for 24 h. Hemolytic activity was observed with transmitted light, and determined by the photochromic properties of the colonies.
For the D-lactate production assay, the candidate strain was inoculated with 5 mL of MRS liquid medium, and incubated at 37°C for 48 h. D-lactate in the supernatant was analyzed by using the D-Lactic Acid Assay Kit (Megazyme, K-DATE) according to the manufacturer's protocol.
Bile salt deconjugation was determined through standard plate assay with minor modification. 28 The candidate strains were inoculated into the MRS agar plate supplemented with 0.5% taurodeoxycholic acid sodium salt hydrate (Sigma, USA), and incubated at 37°C for 48 h. Bile salt deconjugation was determined by the visible halo zone and the opaque white precipitate around the colonies.
The biogenic amine production of the candidate LAB was determined using decarboxylase medium supplemented with different amino acids as described previously. 29 The amino acid used in this study were L-histidine, L-tyrosine, L-ornithine, L-phenylalanine, and L-lysine. The candidate strains were inoculated on the plates, and incubated at 37°C for 16 to 24 h. The formation of purple color around the colonies was analyzed.
The susceptibility of the candidate strains to the antibiotics
The minimum inhibitory concentration (MIC) of different antibiotics was determined using the twofold broth microdilution method. 30 Antibiotics used for tests included chloramphenicol, ampicillin, tetracycline, gentamycin, kanamycin, streptomycin, erythromycin, clindamycin, and vancomycin as recommended by the European Food Safety Authority (EFSA). 31 The MIC cutoff values of various antibiotics, given by EFSA for Lactobacillus brevis, were used to determine the antibiotic susceptibility of MJM60390.
Orogastrointestinal transit assay
Orogastrointestinal transit (OGT) assay was performed in OGT solutions as previously described with minor modification. 32 The LAB strains with initial concentration of 109 CFU/mL were exposed to oral stress by treating the bacterial cells in an oral stress solution (g L−1: NaCl, 6.2; KCl, 2.2; CaCl2, 0.22; NaHCO3, 1.2) containing 150 mg L−1 lysozyme for 10 min. The oral stress solution was removed by centrifugation at 1800 g for 5 min, and remaining cells were exposed to gastric stress solution by treating the cells in a gastric electrolyte solution containing 0.3% pepsin at pH 3 for 1 h. Then, the gastric stress solutions were removed by centrifugation, and the remaining cells were incubated for 120 min in an intestinal electrolyte solution (g L−1: NaCl, 5; KCl, 0.6; CaCl2, 0.25) containing 0.1% pancreatin and 0.3% bile Oxgall at pH 7. Cells incubated in phosphate-buffered saline (PBS) without any stress were used as controls.
The cell viability was determined at each step by plating the cells on MRS agar plates, followed by incubation at 37°C and counting the number of colony-forming units (CFU) after 48 h.
Caco-2 cell culture and adhesion assay
Human enterocyte-like Caco-2 cells were purchased from Korea Research Institute of Bioscience and Bioscience Center (KCTC) and cultured in Dulbecco's Minimum Essential Medium (DMEM) (Invitrogen, Germany) supplemented with 20% (v/v) fetal bovine serum (Gibco, Waltham, MA, USA), 100 U/mL penicillin, and 100 mg/mL of streptomycin (Gibco) at 37°C in a CO2 incubator maintained at 5% CO2. The Caco-2 cells were seeded at a concentration of 2 × 105 cells/well in a 12-well plate and incubated for 24 h to obtain a polarized monolayer and 80% confluence. The medium was changed every day.
The adhesion of the candidate strain to Caco-2 cells was determined as described previously with minor modification. 33 Briefly, Caco-2 monolayers were washed three times with PBS, and the medium was replaced with antibiotic-free DMEM. Each well was inoculated with bacteria (final concentration, 1 × 108 CFU/mL), and incubated for 2 h at 37°C in 5% CO2. After incubation, the monolayers were washed three times with PBS, followed by the addition of 200 μL of 1% Triton X-100 (Sigma-Aldrich). An aliquot of 1 mL of the homogenate was serially diluted and plated on MRS agar and incubated at 37°C for 48 h. The percentage of bacteria that adhered to the cell was calculated by dividing the number of remaining bacteria grown on MRS agar by the number of initial inoculations. All of the experiments were performed in triplicate.
Antimicrobial activity of the culture filtrate of MJM60390
The antimicrobial activity of the candidate strain was determined against human, animal, and food-borne pathogens by using paper disc agar diffusion assay. The test strains included Staphylococcus aureus, Listeria monocytogenes KACC 10764, Bacillus cereus KACC 11240, Bacillus subtilis KSP12, Salmonella typhi KCTC 2514, Salmonella choleraesuis KCTC 2932, Pseudomonas aeruginosa KCCM 11802, Salmonella gallinarum KCTC 2931, and Escherichia coli O1 KCTC 2441.
For the antimicrobial test, MJM60390 was incubated at 37°C for 24 h in MRS broth, and 50 μL of fresh culture filtrate was loaded in a paper disc and dried. Then the paper disc was placed on the agar plate, which has been mixed with the pathogenic strains at a concentration of 106 CFU/mL. The plates were incubated at 37°C for 24 h. The antimicrobial activity of the culture filtrate was determined by measuring the diameters of inhibition zones. All the tests were performed in triplicate.
Proliferation assay
To test the utilization of purines and nucleosides by MJM60390, a chemically defined growth medium containing the pyrimidines thymine, cytidine and 2′-deoxyuridine (DM-py) 24 supplemented with purines or nucleosides was used for the proliferation assay. DM medium containing different purines or nucleosides (final concentration, 400 μM) were inoculated with LAB at the concentration of 2 × 108 CFU/mL, and incubated at 37°C for 0, 2, 4, 6, and 8 h under anaerobic conditions. The growth of LAB was monitored at OD600 using a spectrophotometer (Tecan, infinite M200PRO, Austria). DM medium without any purines or nucleosides was inoculated with the same amount of LAB and used as the control.
Animals and treatment
Male C57BL/6J mice (7 weeks-old, n = 8/group) were purchased from RaonBio (Gyeonggi, South Korea). All animals were housed in standard plastic cages, and maintained under a 12-h light–12-h dark cycle at constant temperature and humidity (22°C ± 2°C and 55% ± 5%, respectively) with free access to food and water.
The protocols for animal experiment were approved by the Committee on the Ethics of Animal experiments of the Myongji Bioefficacy Research Center in Myongji University (MJIACUC-2019003), and the mice were maintained in accordance with the NIH Guidelines for the Care and Use of Laboratory Animals.
After acclimatization for 1 week, the mice were divided into five groups: (1) control group (Control), (2) hyperuricemia group (Hyperuricemia), (3) allopurinol-treated group (Allopurinol), (4) L. gasseri MJM60662-treated group (MJM60662) and (5) L. brevis MJM60390-treated group (MJM60390). To suppress the activity of uricase, potassium oxonate (300 mg/kg B.W. per day)-carboxymethylcellulose sodium (GMC-Na) solution (3 g/L) was intraperitoneally injected into the mice in groups 2 to 5 daily. Meanwhile, the mice in group 2 to 5 were supplied with HPD containing additional 570 g yeast extract in the normal diet (AIN-93 M; RaonBio), while the control group was supplied with normal diet.
For treatment, LAB strains were cultured fresh daily, washed, and resuspended in saline (0.85% NaCl) for use. The MJM60662 and MJM60390 groups were administered with L. gasseri MJM60662 (3 × 109 CFU/mice/day) and L. brevis MJM60390 (3 × 109 CFU/mice/day), respectively, once daily for 14 days by oral gavage, and the control group and hyperuricemia group were administered saline alone. The allopurinol group was exposed to allopurinol (5 mg/kg B.W.; Sigma). The body weight was recorded on days 0, 7, and 14.
Serum biochemical analysis
The blood samples were collected from the heart on the days 0, 7, and 14. The collected samples were immediately moved to ice and centrifuged at 2000 g for 15 min at 4°C. The serum was separated, and stored at −80°C until analyses. The levels of uric acid, creatinine and blood urea nitrogen (BUN) were measured using a biochemical blood analyzer (FUJIFILM DRI-CHEM NX500i, Tokyo, Japan). Xanthine oxidase (XO) levels in the serum of each group were measured using the XO activity assay kit (Sigma-Aldrich, Saint Louis, MO, USA). The concentration of urate transporter 1 (URAT1) in the serum was measured using the SLC22A12 Elisa Kit (MyBioSource, USA).
Kidney sectioning and histopathological assessment
At the end of the experiment, mice were euthanized with isoflurane 3% for anesthesia induction and cervical dislocation. The intestine was dissected, and the kidney from each animal was immediately removed, rinsed, and fixed in 4% paraformaldehyde (Sigma-Aldrich, Germany), followed by the paraffin-embedding and H&E staining.
For quantification of changes in glomerulus morphology and hypertrophy, the glomerular image was digitally sectioned into 20 segments, and each segment contained 2 glomeruli. The glomeruli, in which the vascular pole was evident, were selected and the area was assessed using the “draw closed polygon” tool in CaseViewer by 3DHISTECH (Budapest, Hungary). All areas were recorded and analyzed.
Fecal sample analysis
After scarification, the intestine was immediately dissected, and the feces were collected in a tube and stored at −80°C before use. For metagenome analysis, DNA was extracted with the Exgene™ Stool DNA Mini Kit (GeneAll, South Korea) according to the manufacturer's protocol. The V3-V4 region of the bacterial 16S rRNA gene was amplified using barcoded universal primers, 341F and 805R. 34 Microbiome profiling was conducted on the 16S-based Microbial Taxonomic Profiling platform of EzBioCloud Apps (ChunLab, Inc.).
For extraction of short-chain fatty acid (SCFA) in the fecal sample, 0.5 g of each fecal samples was thawed, suspended in 2 mL of water, and homogenized for 5 min. The pH of the suspension was then adjusted to 3 with 5 M HCl, and then kept at room temperature for 10 min with occasional shaking. The suspension was then centrifuged at 4690 g for 20 min, and the supernatant was used for gas chromatography–mass spectrometry (GC-MS) analysis (Thermo Fisher Scientific). The content of SCFAs in each sample was then calculated by comparing with the standard.
Statistical analyses
All data were expressed as mean ± standard deviation. The statistical differences between control and experimental groups were analyzed using one-way analysis of variance, or a two-sample t-test to determine statistical significance. All statistical analyses were carried out in GraphPad Prism software (version 8.0). P values of 0.05 or less between groups were considered statistically significant.
Results
Screening of LAB strains degrading inosine and guanosine
To screen potential probiotic strains, a total of 24 strains were tested for the assimilation of inosine and guanosine by HPLC (Table 1). As a result, the assimilation rate of guanosines by these strains ranged from 11.8% to 100%, with seven strains assimilating more than 50% of guanosine in 2 h. For inosine, the assimilation rate ranged from 11.1% to 99.8%, and eight strains assimilated more than 50% of inosine. Although some strains prefer guanosine, and others prefer inosine, most strains utilized both guanosine and inosine at the same level. Among the tested strains, strain MJM60390 showed the strongest assimilation ability of both guanosine (100%) and inosine (99.8%), and thus it was selected for further study. L. gasseri MJM60662 was isolated from yogurt, which showed antihyperuricemia activity, 35 and used as a control in this study.
High-Performance Liquid Chromatography Analysis of Purine Assimilation by Candidate Lactic Acid Bacteria
Values are means, n = 3. Agua, the assimilation rate of guanosine by LAB strains; Vgua, the assimilation speed of guanosine by LAB strains; Aino, the assimilation rate of inosine by LAB strains; Vino, the assimilation speed of inosine by LAB strains.
Highest similarity based on the 16S rDNA sequence.
LAB, lactic acid bacteria.
Identification of MJM60390 by phylogenetic analysis
BLAST search of the Ezbiocloud database showed that the 16S rRNA full sequence of MJM60390 showed 99.98% (difference of 1/1499 bp) similarity with L. brevis ATCC 14869T. Phylogenetic analysis of the 16S rRNA gene sequence of MJM60390 compared with various Lactobacillus species revealed a close relationship with L. brevis (Fig. 1).

Neighbor-joining tree based on the 16S rRNA gene sequences showing the phylogenetic relationship of strain MJM60390 and its related taxa. The accession numbers in the parentheses were obtained from NCBI. Numbers at nodes indicate percentage levels of bootstrap support based on a neighbor-joining method of 1000 replications. The evolutionary distances were computed using the Kimura 2-parameter method. Scale bar indicates 0.002 substitutions per nucleotide position.
In vitro safety assessment of MJM60390 of hemolytic activity, D-lactate production, biogenic amine production, and susceptibility to antibiotics
Strain L. brevis MJM60390 tested negative for hemolytic activity on blood agar, D-lactate production, bile salt deconjugation, and bioamine production on the decarboxylase agar media supplemented with various amino acids (Table 2). Strain L. brevis MJM60390 showed susceptibility to antibiotics (except kanamycin B) according to the cutoff value recommended by EFSA (2012) (Table 2).
In Vitro Safety Assessment of L. brevis MJM60390 and L. gasseri MJM60662
MIC value (mg/L) for the antibiotics recommended by EFSA, 2012.
R, resistant; —, no activity.
EFSA, European Food Safety Authority; MIC, minimum inhibitory concentration.
Tolerance to OGT
L. brevis MJM60390 showed good tolerance to the simulated oral, gastric, and intestinal stress in vitro. Both L. brevis MJM60390 and L. gasseri MJM60662 showed good resistance to the lysozyme under oral stress. The initial numbers of cells subjected to the stress were 9.08 ± 0.02 log CFU/mL for L. brevis MJM60390 and 8.90 ± 0.03 log CFU/mL for L. gasseri MJM60662, respectively. The viability after oral, gastric, and intestinal stress for L. brevis MJM60390 was 8.57 ± 0.21, 8.67 ± 0.08, and 7.55 ± 0.07 log CFU/mL, respectively. The viability of L. gasseri MJM60662 was 8.23 ± 0.04, 7.35 ± 0.02, and 7.35 ± 0.02 after oral, gastric, and intestinal stress, respectively. Taken together, L. brevis MJM60390 showed a reduction of 1.53 log units of CFU (P < .005) at the end of the orogastrointestinal (OGI) assay compared with the initial bacteria number, and L. gasseri MJM60662 exhibited a 1.55 log unit reduction (P < .005) after the assay (Fig. 2).
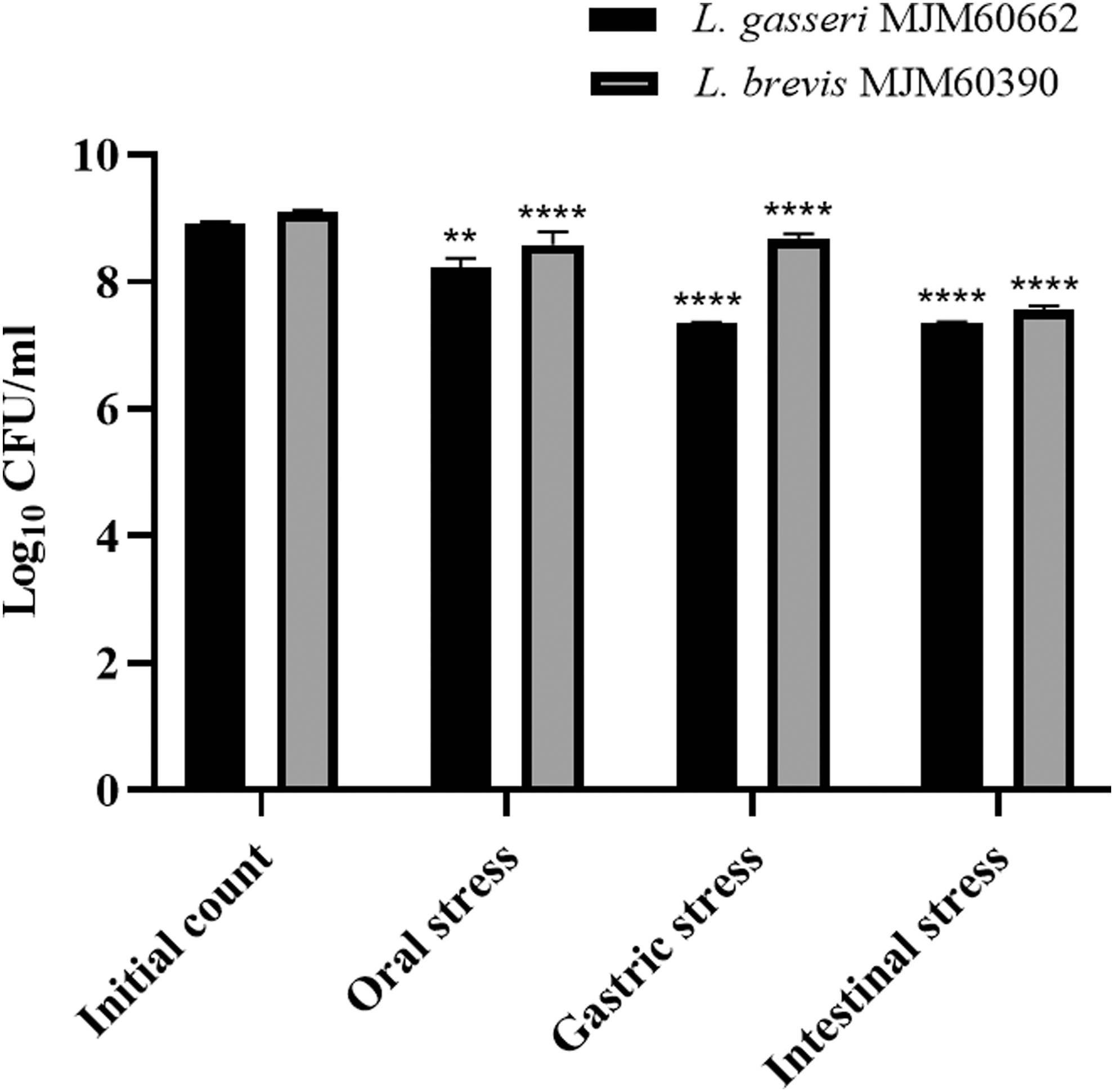
The tolerance of Lactobacillus brevis MJM60390 and Lactobacillus gasseri MJM60662 to the biological barriers of OGT. **P < .01, ****P < .0001, compared with the initial count. OGT, orogastrointestinal transit.
Adherence of MJM60390 to Caco-2 cells
The adherence of L. brevis MJM60390 and L. gasseri MJM60662 to Caco-2 cells was calculated by the percentage of bacteria remaining attached to the Caco-2 cells after washing compared with the inoculated number of bacteria. The adherence capacity of L. brevis MJM60390 was 8.87%, whereas that of L. gasseri MJM60662 was 7.20% (Fig. 3). There was no significant difference between these strains. All values are mean (n = 4).
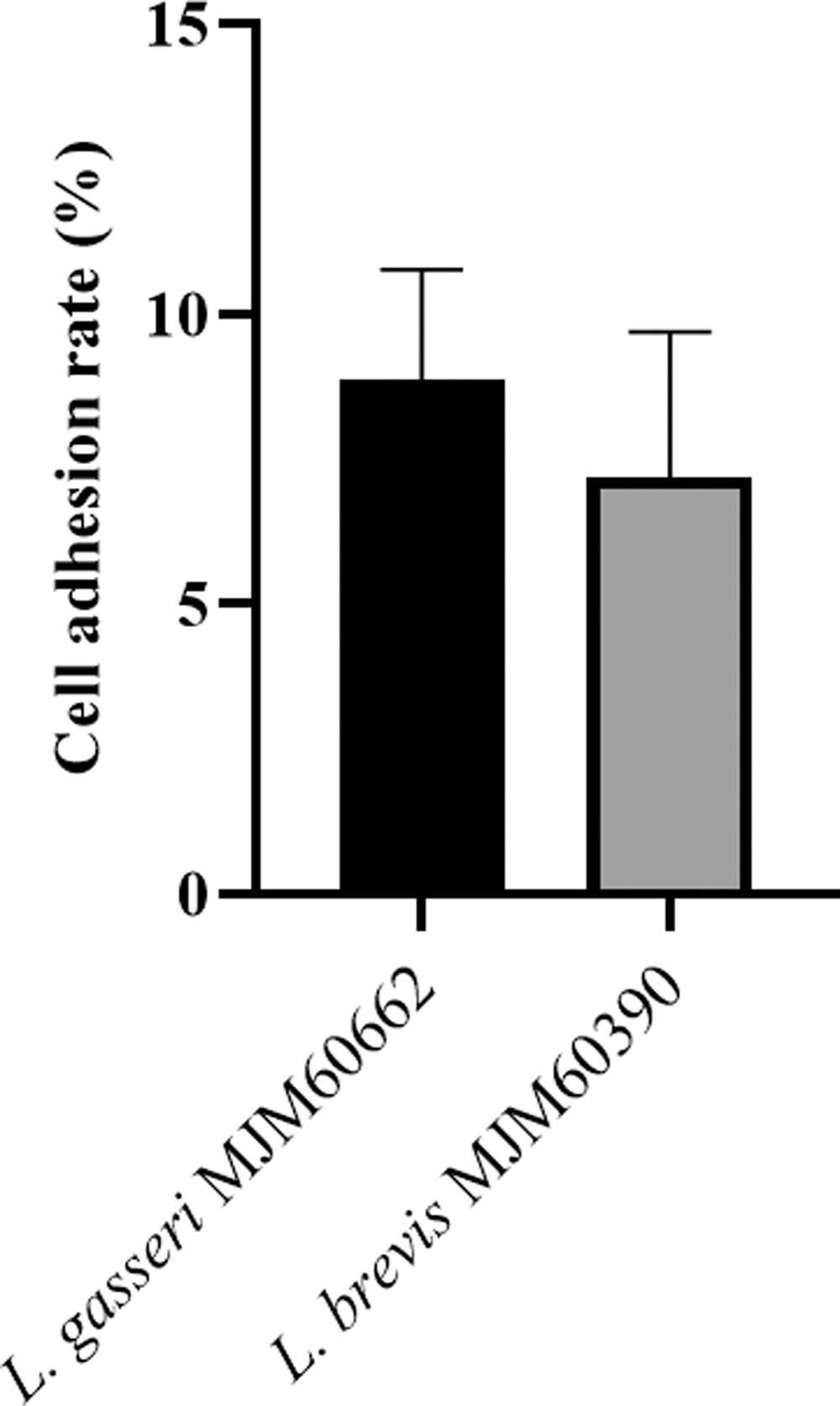
The adherence of L. brevis MJM60390 and L. gasseri MJM60662 to Caco-2 cells. Adhesion rate of L. brevis MJM60390 and L. gasseri MJM60662 to Caco-2 cells was calculated as the percentage of cells that remain attached to the Caco-2 cells when compared with the initial plate count of the added bacterial cells.
Antimicrobial activities of candidate LAB strain
L. brevis MJM60390 showed inhibitory activity against some intestinal and food-borne pathogens (Table 3). The culture filtrate of L. brevis MJM60390 inhibited the growth of Salmonella galinarum and L. monocytogenes more strongly than Pseudomonas aeruginosa, L. monocytogenes, and E. coli. L. gasseri MJM60662 inhibited P. aeruginosa the most among the tested strains.
Antibacterial Activity of the Culture Filtrate of Candidate Strains
KCTC, Korea Research Institute of Bioscience and Bioscience Center.
Proliferation assay
Both L. brevis MJM60390 and L. gasseri MJM60662 showed greater proliferation when cultured in the presence than in the absence of adenine, adenosine, and adenosine monophosphate (AMP). The cell growth of L. brevis MJM60390 was the highest in the presence of AMP, followed by adenosine and adenine (Fig. 4A). In contrast, L. gasseri MJM60662 preferentially utilized adenine rather than adenosine and AMP (Fig. 4B).
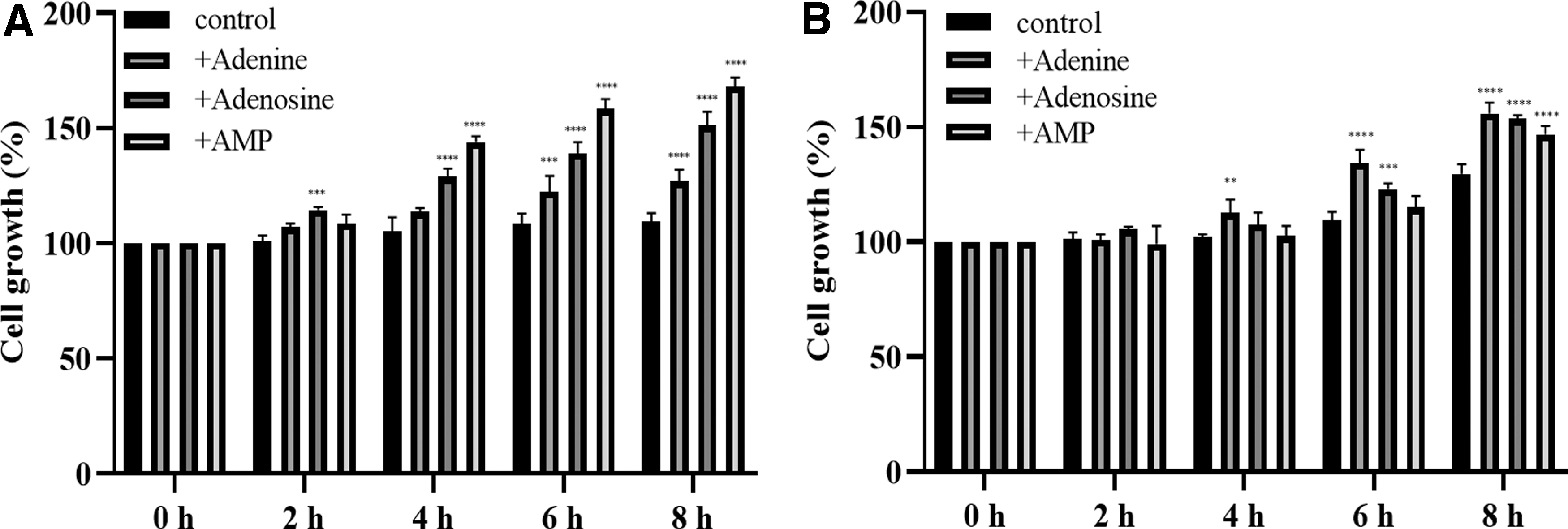
Cell proliferation in the presence of adenine, adenosine, and AMP. The growth of L. brevis MJM60390
Effect of MJM60390 on the body weight and the serum uric acid level
In the animal study, hyperuricemia was induced by intraperitoneal injection of potassium oxonate and a HPD. Mice with hyperuricemia were treated with L. brevis MJM60390 and L. gasseri MJM60662 for 14 days (Fig. 5A). The control group was fed with normal diet, and the other groups were fed a HPD to increase the purine uptake from food. Hyperuricemia group served as the symptom control group, and the allopurinol-treated group was used as the positive control. Body weight and blood samples were measured on days 0, 7, and 14.
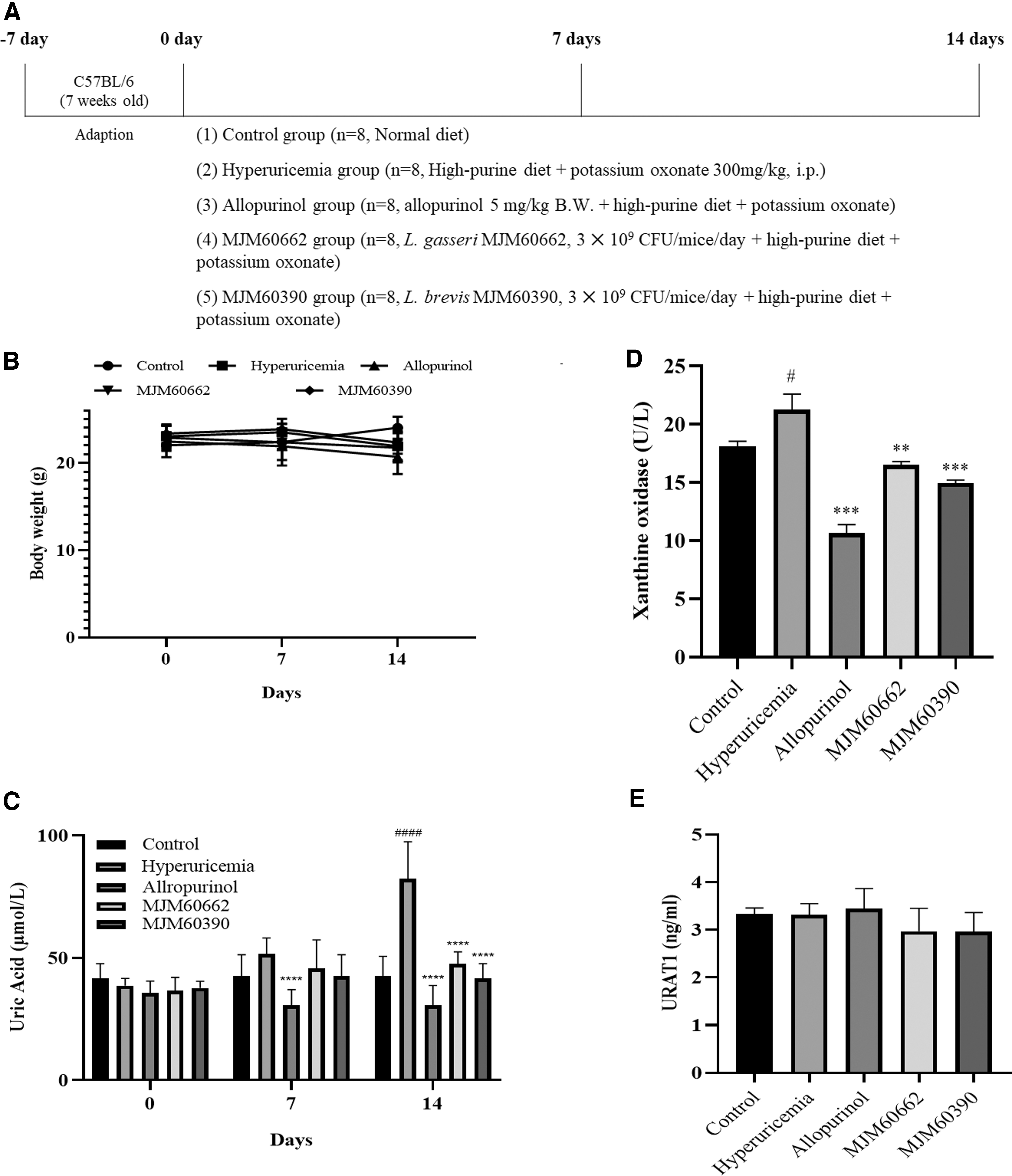
The effect of LAB administration on the uric acid concentrations in a hyperuricemia model.
As shown in Figure 5B, the body weight gain was not significantly different among all groups (Fig. 5B). The uric acid level did not differ on day 0 and 7. Only the allopurinol-treated group showed a significant decrease in uric acid level on day 7. The uric acid level of hyperuricemia group gradually increased, and reached 82.29 μmol/L on day 14. The uric acid level in hyperuricemia group increased almost twofold compared with control (42.63 μmol/L), indicating the successful establishment of hyperuricemia in mice. The serum uric acid levels on day 14 were 42.63, 82.29, 30.73, 47.58, and 41.64 μmol/L in control, hyperuricemia group, allopurinol group, MJM60662-treated group, and MJM60390-treated group, respectively.
Administration of L. brevis MJM60390 had significantly decreased the serum level of uric acid by 50% when compared with hyperuricemia control, and the uric acid level was reduced to the level in normal control (Fig. 5C).
XO and URAT1 level in serum
XO is an enzyme that converts hypoxanthine into xanthine, and xanthine into uric acid. The amount of uric acid is increased by high levels of XO. The XO level was 18.07 U/L in the control group, 23.47 U/L in hyperuricemia group, 10.65 U/L in allopurinol group, 16.49 U/L in MJM60662 group, and 14.94 U/L in MJM60390 group (Fig. 5D). Although the decrease was less compared with allopurinol, both MJM60662 and MJM60390 groups showed lower XO levels than the hyperuricemia group, and XO levels in MJM60390 group are 1 U/L lower compared with the MJM60662 group (Fig. 5D).
URAT1 protein facilitates the reabsorption of uric acid in the kidney. The URAT1 levels in the control, hyperuricemia, allopurinol, MJM60662, and MJM60390 groups were 3.31, 3.38, 3.71, 3.26, and 3.31 ng/mL, respectively. There was no statistically significant difference among them (Fig. 5E).
Effect of LAB on kidney by histological observation and biochemical analysis
Approximately 70% of uric acid is currently excreted from the kidneys and, therefore, the increase in serum uric acid levels can alter the kidney structure, including glomerulus, Bowman's capsule, and tubules. 36 Our study showed a decrease in the Bowman's capsule space and dilatation of the proximal tubule cells. These morphological changes were more serious in the hyperuricemia group, and attenuated in the allopurinol groups, which showed an increase in the Bowman's capsule space and altered circumference of each glomerulus. MJM60390 group showed an increase in the Bowman's capsule space more similar to the control group (Fig. 6A).

Kidney H&E staining and creatinine, urea nitrogen level in serum.
To avoid any subjective evaluation of the changes in kidney pathology, we calculated the glomerular tuft area of each sample. This area reflects hypertrophy of each glomerulus, in which the hypertrophic structure is larger than in nontreated animals. 37,38
The hyperuricemia group showed a glomerular tuft area of 3712.6 ± 1358.5 μm, 2 representing an increase of 25.3% compared with the control group (2882.9 ± 1123.4 μm2). In the case of allopurinol and MJM60662 groups, the glomerular area of 3296.1 ± 1480.71 μm2 and 3411.9 ± 1325 μm2 represent a decrease of 11.2% and 8%, respectively, compared with the hyperuricemia control group. Finally, the MJM60390 group showed a 2962.6 ± 1027.3 μm2 of glomerular area, which represented 20% of decrease compared with hyperuricemia group.
This result confirmed the effectiveness of L. brevis MJM60390 to attenuate the morphologic changes in kidney observed in mice treated for hyperuricemia and the increase in the percentage of accurate organ function (Fig. 6B).
Creatinine and BUN are indicators of kidney function. The creatinine level was increased in the hyperuricemia group, especially in the allopurinol group (Fig. 6C). There was no significant difference in urea nitrogen level in the serum (Fig. 6D).
SCFA contents in the fecal sample
The concentrations of SCFA in the feces were measured in each group. Quantities of acetic acid, propionic acid, and butyric acid were measured by GC-MS. Acetic acid levels detected in the control group, hyperuricemia group, allopurinol group, MJM60662 group, and MJM60390 group were 92, 69, 81, 72, and 76 mmol/L, respectively. The levels of propionic acid in these groups were 15, 44, 30, 72, and 76 mmol/L, respectively. Butyric acid levels in these treated groups were 13, 19, 17, 19, and 26 mmol/L, respectively. Mice supplemented with L. brevis MJM60390 generated the highest levels of butyric acid in their feces (Fig. 7).
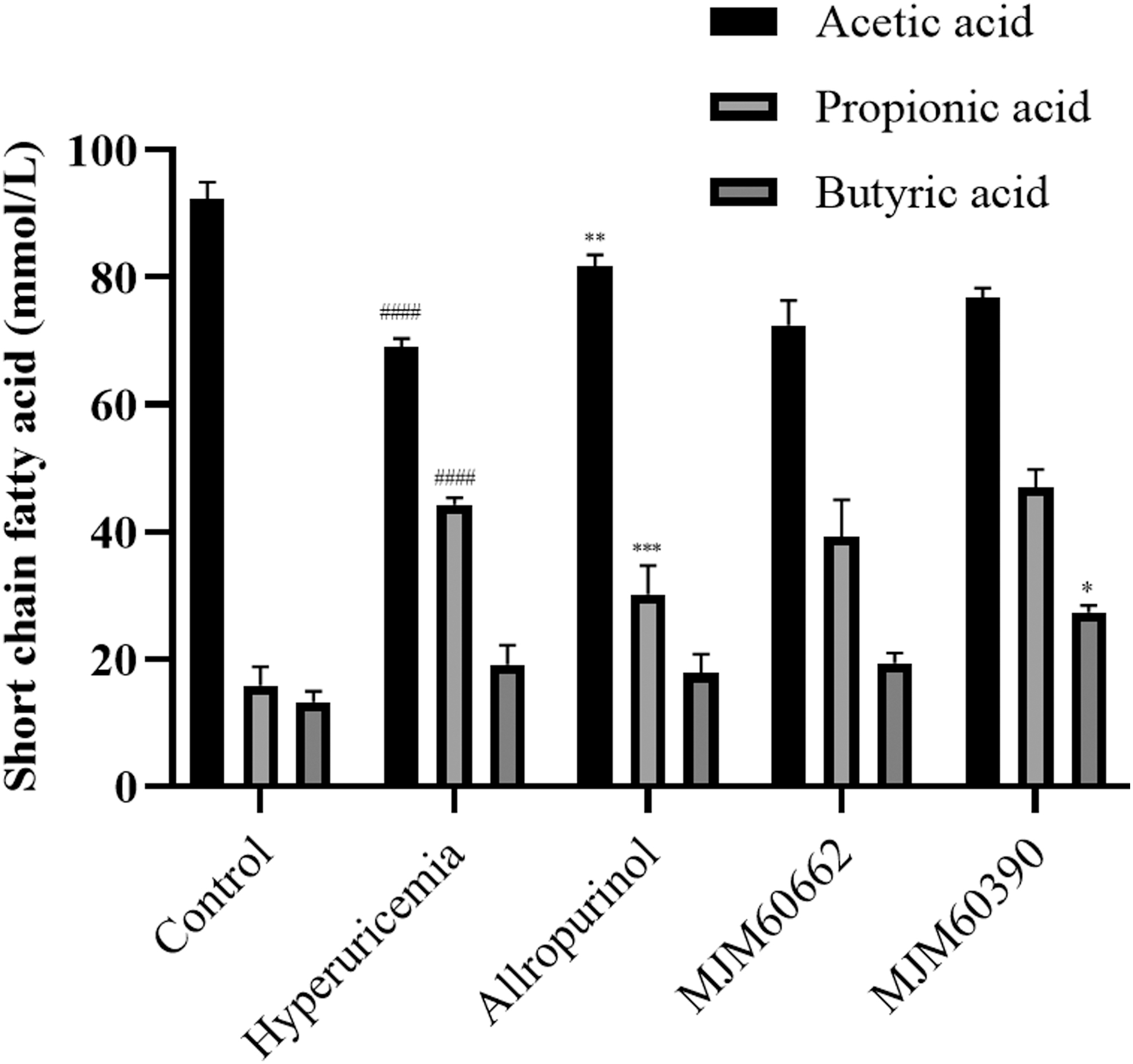
The SCFA concentration in the fecal samples. The amounts of acetic acid, propionic acid, and butyric acid were measured in the feces of each group. MJM60390: mice treated with L. brevis MJM60390; MJM60662: mice treated with L. gasseri MJM60662. Data are shown as mean ± SDs (n = 3). #### P < .0001, compared with control group; *P < .05, **P < .01, ***P < .001, compared with hyperuricemia group. SCFA, short-chain fatty acid.
Influence of LAB on gut microbiota
To identify specific taxa related to probiotic supplementation, the relative abundance of bacterial profile was assessed at the phylum and family levels. Mice from the hyperuricemia-induced groups (including hyperuricemia, allopurinol, MJM60662, MJM60390 groups) showed notable shifts in gut microbial composition compared with the control group (Fig. 8A, B).
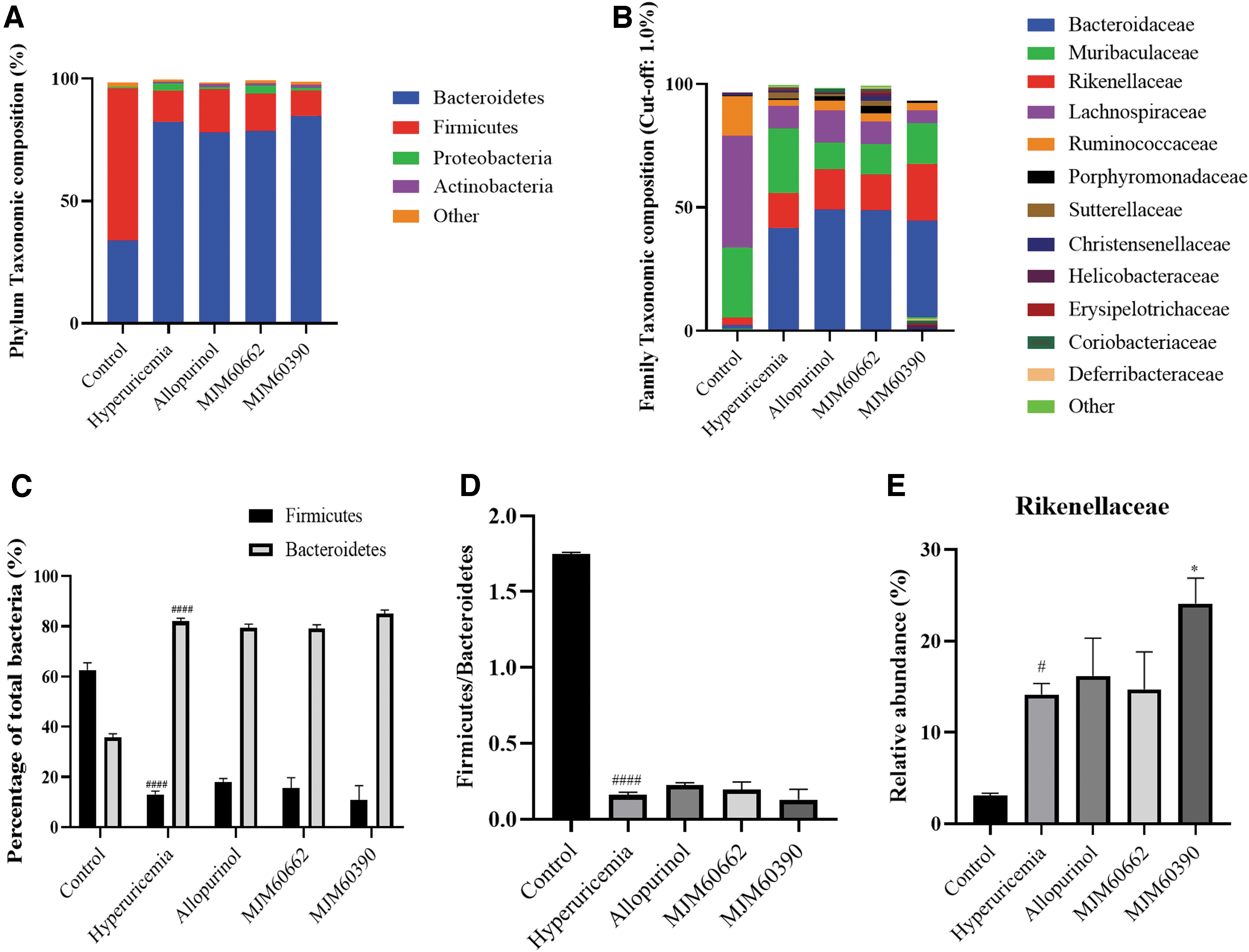
Supplementation of L. brevis MJM60390 modulated the composition of the gut microbiota.
Firmicutes and Bacteroidetes were known as predominant gut microbiota in mice. Compared with the control, the relative abundance of Firmicutes reduced, while Bacteroidetes increased in the hyperuricemia-induced groups (Fig. 8C). The hyperuricemia-induced group had a significantly lower Firmicutes/Bacteroidetes ratio compared with the control group, which was not reversed by probiotic treatments (Fig. 8D). At the family level, Lachnospiraceae, Muribaculaceae, and Ruminococcaceae were the predominant families in the control group, while Bacteroidaceae, Rikenellaceae, and Muribaculaceae were the most abundant in the hyperuricemia-induced group (Fig. 8B). It is worth noting that the relative abundance of Rikenellaceae was higher in the MJM60390 group than in other hyperuricemia-induced groups (Fig. 8E).
Discussion
Due to the changes in lifestyle, such as increased consumption of meat and processed foods, the incidence of hyperuricemia has increased, resulting in significant health concerns worldwide. 39 A high level of uric acid in serum (hyperuricemia) has been reported to be associated with many metabolic diseases. However, hyperuricemia is commonly asymptomatic. Even if uric acid crystals existed in the joints, the rate of subjective judgment as negative was up to 21.2%. 40
Lack of prompt treatment can lead to urgent surgery or hospitalization. Therefore, maintaining the normal level of serum uric acid is important to prevent the development of these diseases. However, it is not recommended to prescribe drugs to reduce the uric acid levels for hyperuricemia patients without symptoms. Therefore, it is very important to find a safe and alternative way to manage the hyperuricemia.
Probiotics are commonly regarded as safe and have been reported to show beneficial effect in humans. Many recent studies have revealed their important role in the management of various diseases, especially in the metabolic disorders. Some previous studies have demonstrated that the administration of probiotics can attenuate hyperuricemia through different mechanisms.
LAB isolated from Chinese sauerkraut can degrade purine compounds, and lower the serum uric acid level in a hyperuricemia rat model. 41 L. gasseri PA3 was found to incorporate nucleoside and purine derivatives into the cell, and thereby reduced their absorption in rats. 42 A recent study reported Limosilactobacillus fermentum JL-3, a bacterial strain isolated from fermented vegetables, ameliorates hyperuricemia in mice by degrading uric acid. 39 However, the mechanism by which probiotics reduce hyperuricemia is still being studied, and not completely understood.
In the present study, we screened 24 LAB strains based on their ability to degrade guanosine and inosine in vitro. Our results showed that most of the candidate strains can assimilate both guanosine and inosine. However, in a few strains, purine assimilation was selective (Table 1).
Lactobacillus plantarum MJM60404 showed a moderate guanosine assimilation rate of 72.6%, and a very low inosine assimilation rate of 11.1%. In contrast, Enterococcus faecium MJM60401 assimilated 16.1% of guanosine, and 63.9% of inosine. The assimilation rates differ even for similar species. L. brevis MJM60390 can almost completely assimilate both guanosine and inosine, whereas L. brevis MJM60391 showed only moderate assimilation rate of guanosine (44.8%) and inosine (32.5%). These results indicate that the purine decomposition ability is strain specific rather than species specific. L. brevis MJM60390 isolated from fermented cereal was selected for its superior rate of purine assimilation. A commercial strain, L. gasseri MJM60662, which showed antihyperuricemia activity, served as the control in this study.
One mechanism by which LAB degrade purines is that LAB use purines for growth, reducing their absorption in the host. In a proliferation assay using adenine, adenosine, and AMP as substrates, both L. brevis MJM60390 and L. gasseri MJM60662 showed a higher growth rate in the presence than in the absence of the purine compounds. L. brevis MJM60390 prefers AMP, whereas L. gasseri MJM60662 prefers adenine. These results confirmed the differential utilization of purine compounds by Lactobacillus spp. (Fig. 4). The preference of different purines by different strains may be due to the presence of different transporters.
Effective probiotics should be nonpathogenic and nontoxic. 43 Safety tests utilized included hemolytic activity, D-lactate production, bile salt deconjugation, biogenic amine production, and antibiotic susceptibility determinations. Lactobacilli contain DL-lactate racemase, which convert L-lactate to D-lactate that cannot be metabolized by human. The accumulation of D-lactate may cause acidosis in toddlers, children, or patients with short bowel syndrome. 44 Some Lactobacilli can produce biogenic amines, which can have toxic effects in humans. 45 Some probiotics deconjugate bile salts for survival in the intestinal environment.
However, excessive bile salt deconjugation may inhibit the fat emulsification, and the deconjugated bile salt may affect other intestinal microorganisms to produce toxic secondary metabolites. In this study, the L. brevis MJM60390 strain did not produce any D-lactate nor biogenic amine, and no bile salt deconjugation was detected (Table 2). Furthermore, hemolytic activity was negative by the L. brevis MJM60390 strains. Moreover, L. brevis MJM60390 showed susceptibility to most antibiotics tested, except for kanamycin. The resistance to aminoglycoside antibiotics is considered intrinsic and nontransferrable for Lactobacilli.
Survival in the gastrointestinal tract and colonization on the intestinal epithelium are very important for successful probiotic activity. In the simulated OGT assay, L. brevis MJM60390 maintained high viability and only 1.5 log unit CFU (P < .05) reduction was observed at the end of the assay, whereas the reference strain L. gasseri MJM60662 showed a 2-log unit CFU reduction (Fig. 2). Caco-2 cells are commonly used as a model to study the adherence characteristics of putative probiotic strains. In this study, L. brevis MJM60390 showed a cell adhesion rate of 7.2%, which is not very high compared with other experiments; however, the reference strains showed a cell adhesion rate of 8.87%. No significant difference was found between these two strains (Fig. 3).
Moreover, the cell-free supernatant of L. brevis MJM60390 showed inhibitory activity against some enteric and food-borne pathogens (Table 3). These results demonstrated the probiotic effects of L. brevis MJM60390, and suggested it has efficacy as a probiotic supplement.
The antihyperuricemic activity of L. brevis MJM60390 was further investigated in a mouse model. Hyperuricemia was induced through i.p. injection of potassium oxonate and supplementation of HPD. The uric acid level was increased twofold in the hyperuricemia group, and the administration of L. brevis MJM60390 resulted in a significant decrease in the serum uric acid level. Furthermore, the XO was reduced by 30%. XO catalyzes the oxidation of hypoxanthine to xanthine, and can further catalyze the oxidation of xanthine to uric acid. However, the levels of URAT1, a urate transporter related to reabsorption of uric acid, remained unchanged. In this study, oral administration of L. brevis MJM60390 decreased serum XO concentrations, which may contribute to the uric acid-lowering effect (Fig. 5).
Uric acid reduction can improve renal function, ameliorate the proteinuria, and tubulointerstitial damage in type 2 diabetic mice. 46 Blocking uric acid-induced intrarenal inflammation, may contribute to the improvement. In this study, hyperuricemia mice developed glomerular hypertrophy as observed through H&E staining of kidney. Glomerular hypertrophy leads to compression of renal capillaries described as glomerular endotheliosis. 47 The MJM60390 group showed a greater decrease in the glomerular tuft area than the hyperuricemia group (Fig. 6), suggesting that L. brevis MJM60390 prevented glomerular hypertrophy induced by hyperuricemia. L. brevis MJM60390 is a probiotic that can safely lower uric acid levels and prevent hyperuricemia without causing kidney toxicity.
SCFA, including acetate, propionate, and butyrate are the main metabolites produced by the bacteria in the colon through the fermentation of dietary fibers and resistant starch. SCFA supply energy and nutrition to the colonocytes; more importantly, SCFA were reported to play an important role in the management of many metabolic diseases, such as diabetes and obesity. Among the SCFA, butyrate is the major and preferred metabolic substrate for colonocytes, at least 60–70% of energy required for the proliferation and differentiation are provided by the butyrate. Furthermore, butyrate was reported to play an important role in the management of hyperuricemia. 48 In this study, administration of L. brevis MJM60390 increased the butyrate production (Fig. 7), which may be involved in the management of hyperuricemia.
Many studies have shown that gut microbiota is an important factor associated with hyperuricemia. The increased serum levels of uric acid may affect the gut microbiota and alter the composition and diversity of intestinal microbiota. Consistent with previous studies, the hyperuricemia group showed a decrease in the relative abundance of Firmicutes and an increased relative abundance of Bacteroidetes (Fig. 8A). Oral administration of L. brevis MJM60390 cannot restore the changes in bacterial diversity.
However, the relative abundance of Rikenellaceae was increased in the MJM60390 group (Fig. 8B). Strains belonging to family Rikenellaceae are one of the few strains that produce butyrate among the strains belonging to Bacteroidetes, which explains the increased butyrate synthesis in the MJM60390 group. Most of the butyrate is produced by Firmicutes, but among Bacteroidetes, about 10 species of microorganisms are known to produce butyrate. 49
Uric acid is the final metabolic product of purines in the human body and increased uric acid concentrations in serum cause hyperuricemia and gout. The intake of purine-rich foods, such as meat and seafood, is associated with an increased risk of gout.
In this study, 24 LAB isolated from fermented food were screened for the degradation of inosine and guanosine in vitro. Among them, strain MJM60390 was selected because it had the greatest ability to assimilate inosine and guanosine from the solution, and was identified as L. brevis based on the 16S rDNA sequence. During probiotic characterization, L. brevis MJM60390 did not produce bioamines and no hemolytic activity was observed. L. brevis MJM6390 showed susceptibility to antibiotics and exhibited antimicrobial activity against some intestinal pathogens. In addition, L. brevis MJM60390 tolerated simulated OGI conditions and adhered to Caco-2 cells.
In a potassium oxonate and HPD-induced hyperuricemia animal study, serum uric acid concentrations were significantly decreased to a normal level after oral administration with L. brevis MJM60390 for 2 weeks. XO was also decreased by 30%. Hematoxylin and Eosin staining indicated that the morphological changes in the glomerulus, Bowman's capsule, and tubules in hyperuricemia were reversed after treatment with L. brevis MJM60390. Fecal sample analysis showed that the butyrate content was increased by treatment with L. brevis MJM60390. Metagenomic DNA analysis showed that L. brevis MJM60390 supplementation enhanced the relative abundance of the Rikenellaceae family, a butyrate-producing bacteria.
In conclusion, these data suggested that L. brevis MJM60390 can ameliorate hyperuricemia by decreasing XO production and regulating microbiota, and therefore may be useful as a probiotic candidate for managing hyperuricemia. However, the MJM60390 was administered from day 0; it is rather a prophylactic effect than a therapeutic treatment. It is necessary to investigate the therapeutic effect in a future study and ultimately in human clinical trials.
Footnotes
Data Availability
The raw data used to support the findings of this study will be made available by the authors, without undue reservation, to any qualified researcher.
Ethics Approval
All protocols for animal experiment were approved by the Institutional Animal Care and Committee (IACUC) of Myongji University (MJIACUC-2019003) and thus conducted in accordance with the NIH Guide for the Care and Use of Laboratory Animals.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was carried out with the support of “Cooperative Research Program for Agriculture Science and Technology Development (Project No. PJ01562101)” Rural Development Administration, Republic of Korea.