
Abstract
Epidemiologic studies suggest an inverse correlation between coffee consumption and the occurrence of neurodegenerative diseases, but the role of caffeine and roasting degree are still matter of debate. The objective of this work was to evaluate the effects of caffeinated (light, medium, and dark roast) and decaffeinated instant coffee samples in acetylcholinesterase (AchE) inhibition and antioxidant assays, as well as in animal models of Parkinson's disease. Caffeinated coffees inhibited the AchE in much smaller concentrations than decaffeinated coffee. All coffee samples showed antioxidant capacity without relation with the caffeine content. Blockade in the haloperidol-induced catalepsy was observed with caffeinated coffee, but not in the decaffeinated sample. The medium-roast coffee reduced the number of rotations of rats after methamphetamine administration on the 6-hydroxydopamine unilateral lesion of the medial forebrain bundle. However, the coffee treatment did not avoid the loss of dopaminergic neurons on substantia nigra pars compact and only the smallest dose of coffee was able to avoid the decrease of dopamine levels in the lesioned side of the striatum. Altogether, these results suggest that coffee exerts moderate pro-cholinergic and pro-dopaminergic effects and caffeine seems to be the main factor responsible for these effects.
Introduction
Coffee is one of the most popular beverages worldwide, exhibiting a complex source of multiple bioactive constituents, including alkaloids, diterpenes, chlorogenic acids, among others. 1,2 Epidemiologic studies suggest an inverse correlation between regular coffee consumption and neurodegenerative diseases, such as Parkinson's disease (PD) and Alzheimer's disease (AD). 3 –5
PD is characterized mostly by the loss of dopaminergic neurons from the substantia nigra pars compacta (SNpc) and by α-synuclein deposition, in the form of Lewis bodies. 6 Clinical symptoms include akinesia or bradykinesia, impaired balance, postural changes, and in many cases, cognitive impairment. AD is mainly characterized by accumulation of amyloid-beta peptide (neuritic plaques) and neurofibrillary tangles with progressive degeneration of cholinergic neurons in neocortex, limbic system, and several other brain areas leading to progressive loss of cognitive functions. 7 Oxidative stress and neuroinflammation are considered important factors associated with AD and PD development, which makes the use of antioxidants and agents with anti-neuroinflammatory effect a potential strategy to reduce the risk of such diseases. 8
The supposed neuroprotective activity of coffee is attributed mainly to caffeine, a competitive adenosine receptor antagonist. There is a close structural and functional association between adenosine and dopamine (DA) receptors in some areas, especially in the basal ganglia, where adenosine negatively modulates the dopaminergic system, 9 making the use of adenosine antagonists a relevant strategy for PD. Furthermore, the possible neuroprotective effect of other coffee constituents cannot be ignored.
The current medicines used in PD and AD are able to improve the quality of life, but cannot avoid the progressive neuron degeneration, making it important to identify potential protective factors such as diet or life style, that could reduce the risk of developing such neurodegenerative diseases, or drugs which could avoid the disease progression. This study aimed to evaluate the prophylactic/therapeutic potential of decaffeinated and caffeinated instant coffee samples submitted to different roasting degrees, assessing their antioxidant and anticholinesterase activities in vitro and their effects on experimental models of PD.
Materials and Methods
Drugs
Coffee bean water-soluble extracts were purchased from Nestlé®, in four different presentations (light, medium and dark roast coffee, and decaffeinated coffee). Rutin, 2,2-diphenyl-1-picrylhydrazyl (DPPH), 5,5-dithiobis-2-nitrobenzoic acid (DTNB), acetylcholinesterase (AchE), acetylthiocholine iodide, thiobarbituric acid, trichloroacetic acid, caffeine, and 6-hydroxydopamine (6-OHDA) were purchased from Sigma (St. Louis, USA). Apomorphine was obtained from Tocris (St. Louis, USA) and methamphetamine was donated by the Brazilian Federal Police. Haloperidol, rivastigmine, anesthetic, antibiotic, and anti-inflammatory drugs were acquired in local pharmacies.
Phytochemical profile and caffeine quantification
Quantitative analysis of caffeine contents in coffee samples was performed by means of high pressure liquid chromatography (HPLC) analysis using a photodiode array detector (HPLC-PDA) as previously described. 10 The coffee samples were also analyzed by HPLC-electrospray ionization tandem mass spectrometry (ESI-MS/MS) to detect the main compounds. Spectral UV data from all peaks were collected in the spectral range 240–400 nm, and chromatograms were recorded at 270 (caffeine) and 330 nm (caffeoylquinic acid derivatives and feruloylquinic acids derivatives).
Animals
Male Swiss mice 3–4 months old (30–50 g) and male Wistar rats 2 months old (250–400 g) were provided by CEDEME/UNIFESP. Animals were kept in rooms with controlled temperature and 12 h light/dark cycle, with water and food ad libitum. All procedures were made under the approval of the University's ethical committee (No. 1637/09).
Antioxidant assays
DPPH radical scavenging activity
The radical scavenging activity was measured using the DPPH radical according to the method previously described. 11 Coffee samples and rutin (final concentrations 0.625–40 μg/mL) were incubated in a microplate with DPPH methanolic solution for 25 min. The absorbance (A) was measured in a spectrophotometer at 517 nm and the scavenging activity was calculated using the formula: (%inhibition) = [(A control − A sample)/A control] × 100.
Lipid peroxidation assay
Coffee samples (1.25–10 μg/mL) or butylhydroxytoluene (0.2–1 μg/mL) were incubated with brain homogenate at 37°C for 60 min, while duplicates of the samples remained at room temperature. Then, 5% trichloroacetic acid was added, and the microtubes centrifuged at 845 g for 15 min. Thiobarbituric acid (6.7 mg/mL) was added to the supernatant and the mixture boiled for 20 min. After cooling, the absorbance was read in a spectrophotometer at 535 nm. The lipid peroxidation inhibition was determined between the incubated and nonincubated samples, according to the formula: (%inhibition) = [1 − (ΔA sample/ΔA control)] × 100, where ΔA is the difference between the absorbance of incubated and nonincubated sample. 12
AchE inhibition assay
AchE inhibition was evaluated in vitro according to Ellman's method with minor modifications, as previously described 13 Coffee samples (25–800 μg/mL), caffeine (10–100 μg/mL), or rivastigmine (3–100 μg/mL) were pipetted to a microplate containing DTNB (3.3 mM), and AchE 0.05 U/mL and the mixture was incubated at 37°C for 5 min. Finally, the substrate acetylthiocholine iodide (20 mM) was added to the mixture, and the absorbance was monitored at 412 nm for 20 min. The inhibition percentage was calculated according to the formula: (%inhibition) = [1 − (ΔA control − ΔA sample/ΔA control)] × 100, where ΔA is the difference between final absorbance and basal absorbance.
Haloperidol-induced catalepsy
Groups of 11 mice received haloperidol (5 mg/kg, ip), 30 min after the oral administration of water (control group) or coffee extracts (dark roast, medium roast, or decaffeinated coffee at doses of 50, 150, and 500 mg/kg), while the negative control group was pretreated orally with water and then received saline (ip) instead of haloperidol. Cataleptic status was evaluated after 45 min of the haloperidol administration during 10 min. 10 The catalepsy score was obtained dividing catalepsy total time (sec) by the number of times the mice left the bar.
Unilateral 6-OHDA lesion
Surgery
Rats deeply anesthetized with ketamine (90 mg/kg, ip) and xylazine (13 mg/kg, ip) were placed in the stereotaxic apparatus and 12 μg of 6-OHDA (6 mg/mL, solubilized in saline with 0.2% ascorbic acid) were injected at a rate of 0.4 μL/min in the right medial forebrain bundle at the following coordinates: −1.9 mm anteroposterior; −1.9 mm mediolateral; −7.2 mm dorsoventral, calculated from the bregma according to Paxinos and Watson. 14 Saline was injected in negative control animals.
Rotational behavior
Groups of 14–15 rats received oral administration of water (controls), medium roast, and decaffeinated coffee (50, 150, and 500 mg/kg) for 45 consecutive days, initiating 30 days before the 6-OHDA infusion. Rotational behavior was assessed on the 14th day after surgery with the administration of apomorphine (0.1 mg/kg, sc) and on the 15th day, after administration of methamphetamine (2.5 mg/kg, ip). The number of complete (360°) contralateral and ipsilateral rotations was counted by a blind observer for 1 h after the administration of each drug. 10
Immunohistochemistry
Four rats from each group were randomly selected for the immunohistochemistry study. Approximately 24 h after the behavioral test, the animals were anesthetized with urethane (3000 mg/kg, ip) and then euthanized by intra-cardiac perfusion with 1% heparin solution in saline followed by 4% paraformaldehyde in phosphate-buffered saline (PBS). The brains were dissected and stored at −80°C until the moment they were sectioned. Sets of sections equally distributed throughout the anteroposterior brain regions were processed on a cryostat at 40 μm and collected into 24 well-microplates containing anti-freezing solution and stored at −20°C.
Free-floating immunohistochemistry was performed employing mouse monoclonal anti-tyrosine hydroxylase antibody diluted 5000 × in 0.005% bovine serum albumin. The sections were then incubated with the secondary antibody and streptavidin-conjugated peroxidase (Dako K0690) and stained with 3-3′-diaminobenzidine (Dako K3468) according to the manufacturer's protocol. After that, the slices were transferred to gelatin-coated slides and kept at room temperature for 24 h, when they were dehydrated with ethanol gradient and xylol, and finally mounted with Entellan and coverslips.
The slides were screened to identify the sections where the SNpc was clearly separated from the ventral tegmental area by the medial terminal nucleus of the accessory optic tract. The selected sections were photographed, and the images were independently analyzed by two blind investigators using ImageJ software. The number of tyrosine hydroxylase immunoreactive (TH-positive) cells in each side was counted, and the result was expressed as a relative percent in the lesioned side compared to the intact side for each section. 15
Determination of monoamine levels by HPLC
Eight to 10 rats from each group were randomly selected to the dosage of central monoamines by HPLC. Animals were anesthetized as described in the previous session, intracardiacally perfused with PBS, and the fresh brains were frozen at −80°C. Later, the right (lesioned) and left (control) striatum were dissected on a cold surface, weighed, and homogenized and the quantification of monoamines and their metabolites was performed by HPLC according to the method previously described. 10 The peak areas of samples were compared with peak areas of standards of each neurotransmitter or metabolite, which were injected at known concentrations, at intervals between runs (see representative chromatograms in the Supplementary Fig. S1).
Statistical analysis
Data was analyzed using the GraphPad Prism software. The effective concentration (EC50) and inhibitory concentration (IC50) from the in vitro tests were calculated by linear regression using the mean percentage of inhibition for each concentration tested. Data from the behavioral tests, immunohistochemistry, and monoamine quantification were initially submitted to Shapiro–Wilk's normality test and Kolmogorov–Smirnov normality test. Behavioral (catalepsy, rotation) and immunohistochemistry data were found to be nonparametric and were analyzed by Kruskal–Wallis, followed by Mann–Whitney test. The monoamine levels were first analyzed by Kruskal–Wallis (inter group comparison), while the intragroup comparison (left vs. right side) was made by Unpaired t test with Welch's correction for standard deviation, when necessary. Statistical significance was set at P < .05.
Results
Phytochemical profile and caffeine quantification
The chromatographic profiles for each coffee sample are available at the Supplementary Figure S2. We observed a similar qualitative profile for the three caffeinated samples with small differences in the proportion of the constituents detected. The following content of caffeine was detected in the samples of soluble coffee: 0.1% decaffeinated coffee; 9.0% light roast; 5.4% medium roast; 8.4% dark roast. Figure 1 shows the chromatographic profile of medium roast coffee using HPLC-PDA-ESI-MS/MS in positive and negative ionization mode (the chromatographic profile of the light roast, dark roast, and decaffeinated coffee samples are available at the Supplementary Fig. S3). The main compounds identified were caffeic acid, 3-O-caffeoylquinic acid, caffeine, 5-O-caffeoylquinic acid, 3-O-feruloylquinic acid, 4-O-caffeoylquinic acid, and 5-O-feruloylquinic acid.
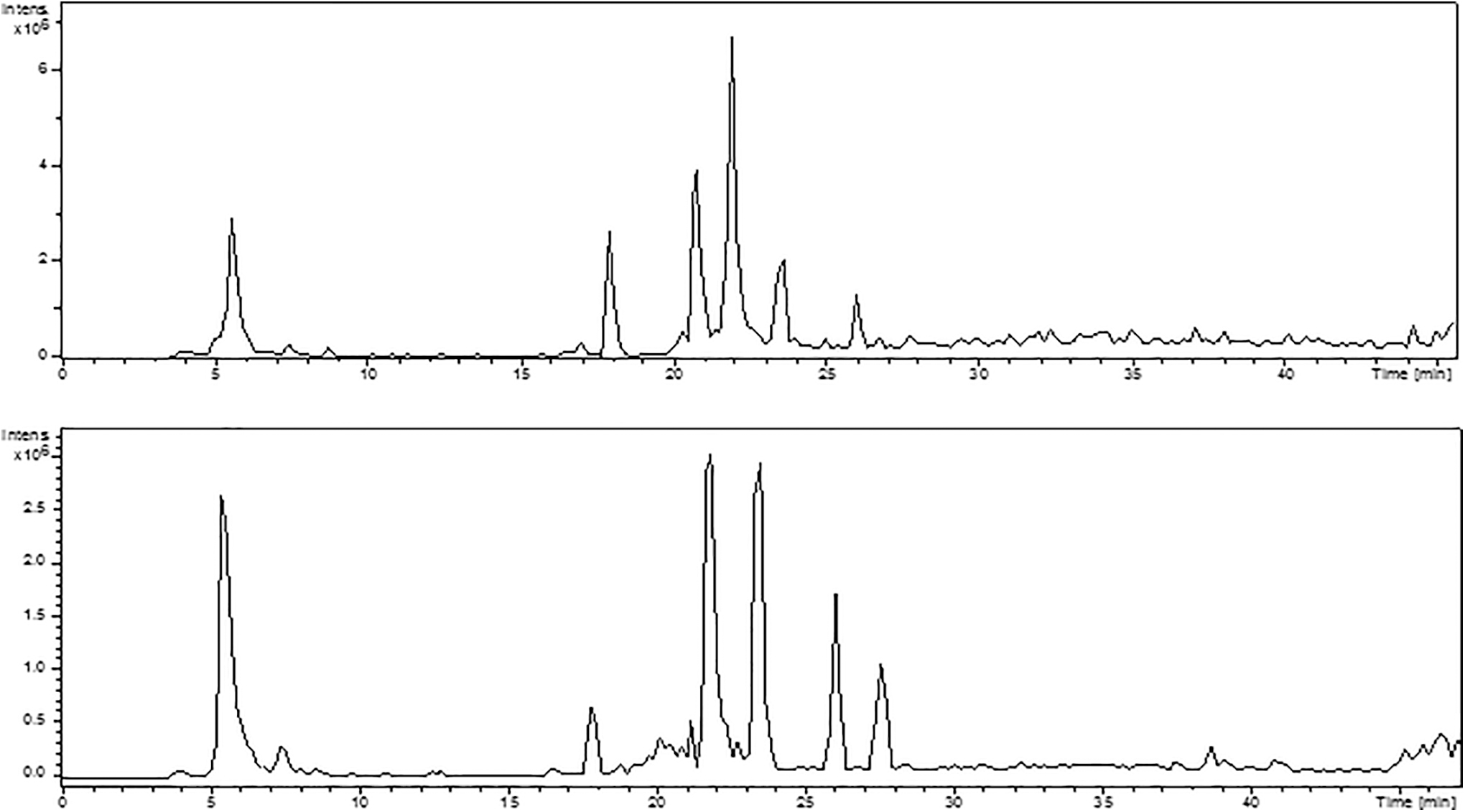
Chromatographic profile obtained by HPLC-ESI-MS/MS in positive ionization mode (upper panel) and negative ionization mode (lower panel) for the medium roast coffee. The main compounds detected were caffeic acid (5.5 min), 3-O-caffeoylquinic acid (17.5 min), caffeine (20.5 min), 5-O-caffeoylquinic acid (chlorogenic acid) (21.8 min), 3-O-feruloylquinic acid (23.5 min), 4-O-caffeoylquinic acid (26.0 min), and 5-O-feruloylquinic acid (27.5 min). Caffeine was not detected in negative ionization mode, and 5-O-feruloylquinic acid was not detected in positive ionization mode. HPLC-ESI-MS/MS, high pressure liquid chromatography electrospray ionization tandem mass spectrometry.
Antioxidant assays
All coffee samples showed reducing power toward the free radical DPPH and inhibited the lipid peroxidation (Table 1) with dark roast coffee presenting the strongest scavenging. On lipoperoxidation assay, the medium roast coffee presented the strongest antioxidant activity, followed by light roast, dark roast, and decaffeinated coffee (Table 1). Rutin and butylhydroxytoluene, used as positive controls, presented typical antioxidant activity in both tests.
Effective Concentration (EC50) or Inhibitory Concentration (IC50) 50% Found for Coffee Extracts and Positive Controls in Two Antioxidant Assays and in Acetylcholinesterase Inhibition Assay, Obtained by Linear Regression Using the Mean Percentage of Scavenging/Inhibition for Each Concentration from Three to Four Independent Assays
BHT, butylhydroxytoluene; DPPH, 2,2-diphenyl-1-picrylhydrazyl; EC50, effective concentration; IC50, inhibitory concentration.
AchE inhibition assay
The light roast coffee had the highest inhibitory effect on AchE, followed by dark roast, medium roast coffee, and decaffeinated coffee (Table 1). Caffeine presented IC50 = 20.54 μg/mL and rivastigmine, used as positive control, IC50 = 22.2 μg/mL.
Haloperidol-induced catalepsy
Kruskal–Wallis test revealed that there was significant difference among the groups [H(10, 119) = 36.59; P < .0001]. Medium roast and dark roast coffee (150 and 500 mg/kg) were able to block the cataleptic status induced by haloperidol (P < .05), while decaffeinated coffee and lower doses of caffeinated coffees did not change the catalepsy score significantly (Fig. 2).
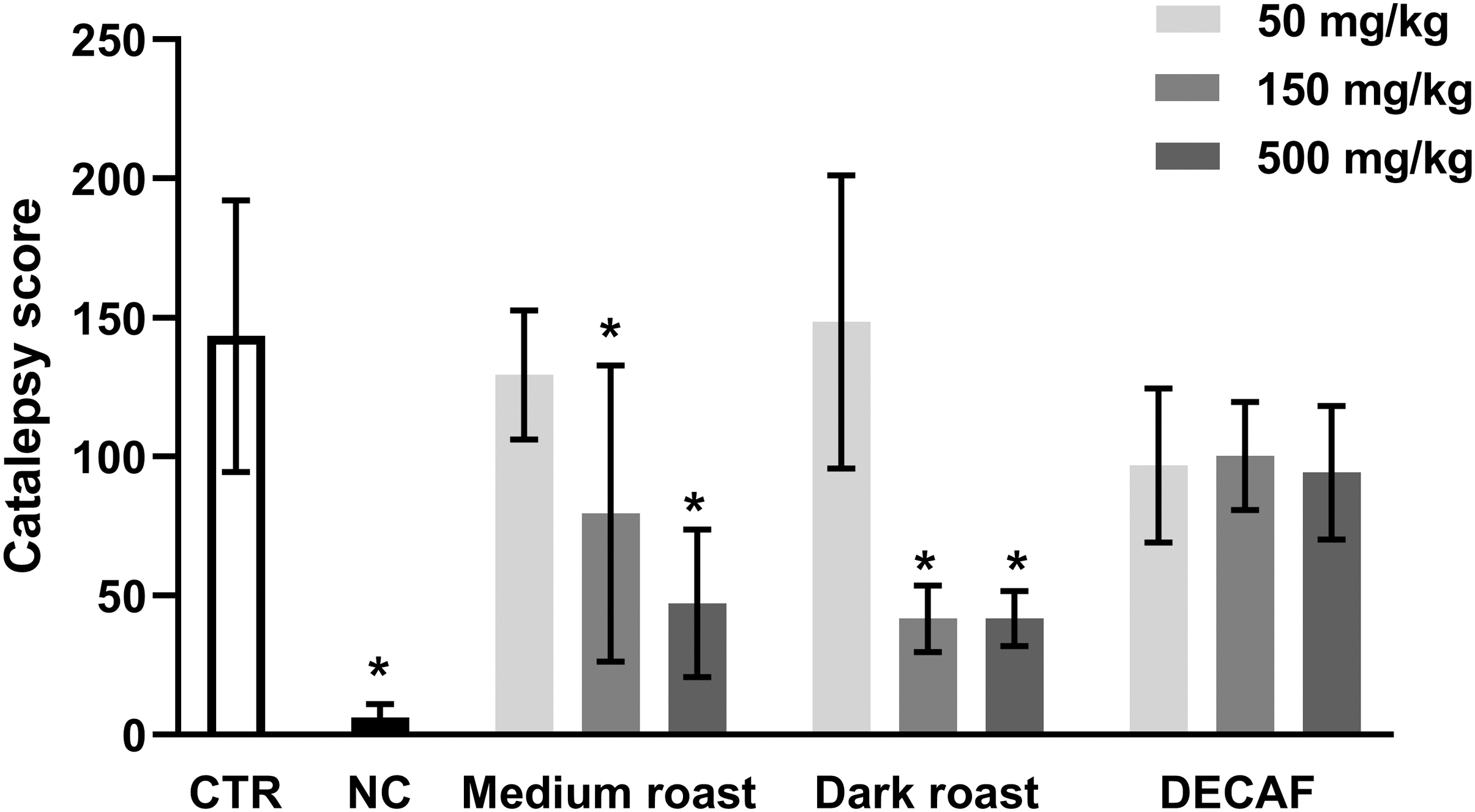
Catalepsy score in mice treated orally with water (CTR, control), 50, 150, and 500 mg/kg of medium roast, dark roast, or DECAF coffee extracts. All animals received haloperidol (5 mg/kg, ip) 30 min after the pretreatment, except the NC, injected with saline. Results presented as mean ± standard error of mean. *P < .05 in comparison to control (Kruskall–Wallis/Mann–Whitney, n = 11). DECAF, decaffeinated; NC, negative control group.
Unilateral 6-OHDA lesion
Rotational behavior
Kruskal–Wallis test revealed significant difference among the groups on contralateral [H(7, 105) = 15.20; P = .0335] and ipsilateral [H(7, 105) = 30.33; P < .0001] rotations. As expected, lesioned control rats presented contralateral rotations induced by apomorphine and ipsilateral rotations induced by methamphetamine, while the negative control group did not show the rotational behavior (Fig. 3). None of the treatments altered the rotational behavior induced by apomorphine when compared with the control group (Fig. 3A). On the other hand, the highest dose of medium roast coffee (500 mg/kg) decreased significantly (P < .05) the number of rotations induced by methamphetamine when compared with the control group (Fig. 3B).
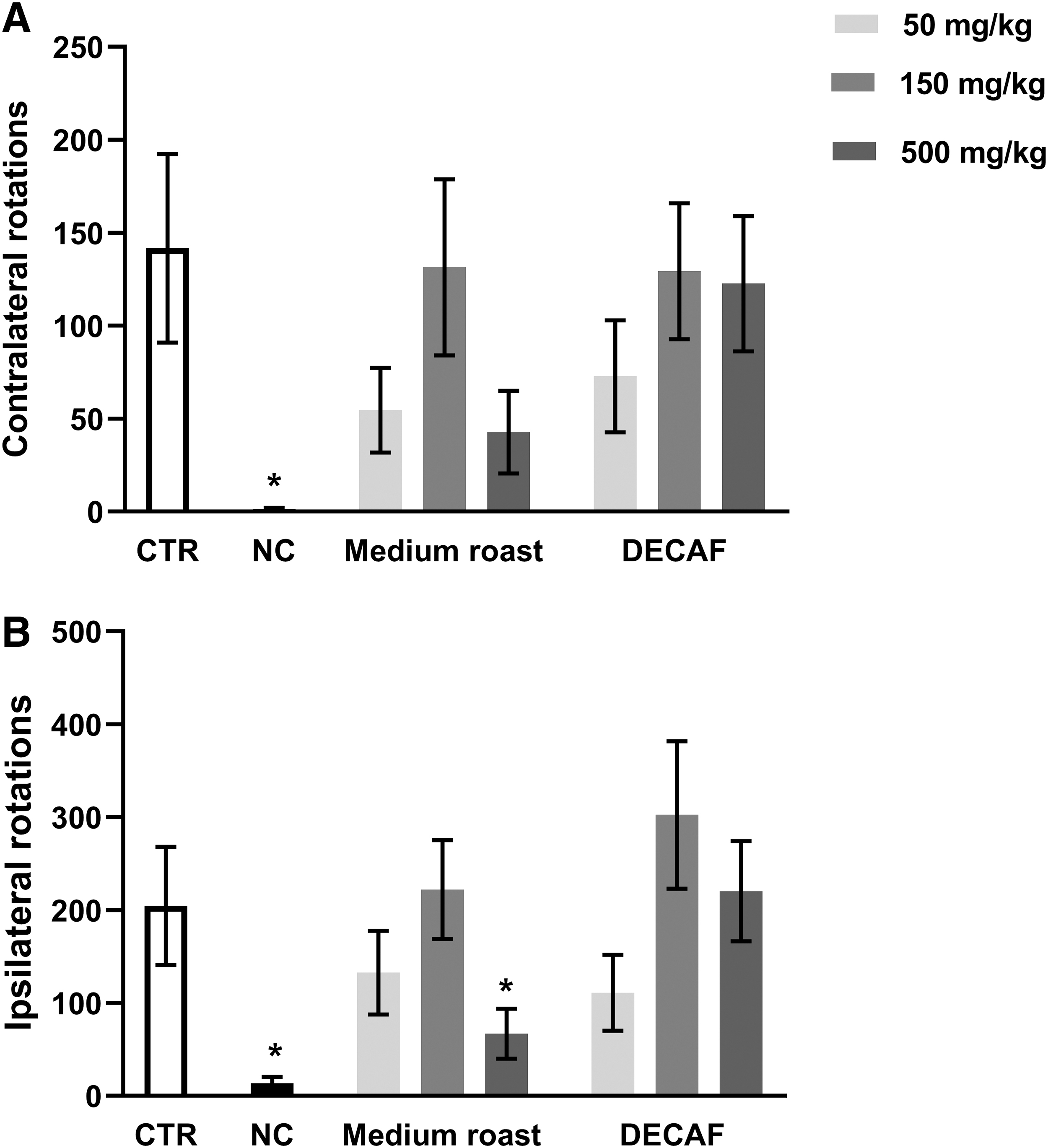
Number of rotations induced by
Immunohistochemistry
The 6-OHDA administration induced a reduction of 50% in the number of TH-positive neurons in the ipsilateral (lesioned side) SNpc on the control group (Fig. 4). The same pattern was observed for the treated groups, which presented a reduction of TH-positive neurons ranging from 40 to 67%. Rats that received saline infusion instead of 6-OHDA (negative control group) presented a small reduction (20%) of TH-positive cells in the ipsilateral side compared to the contralateral side, but no significant differences were observed [H(7, 25) = 9.751; P = .2031].
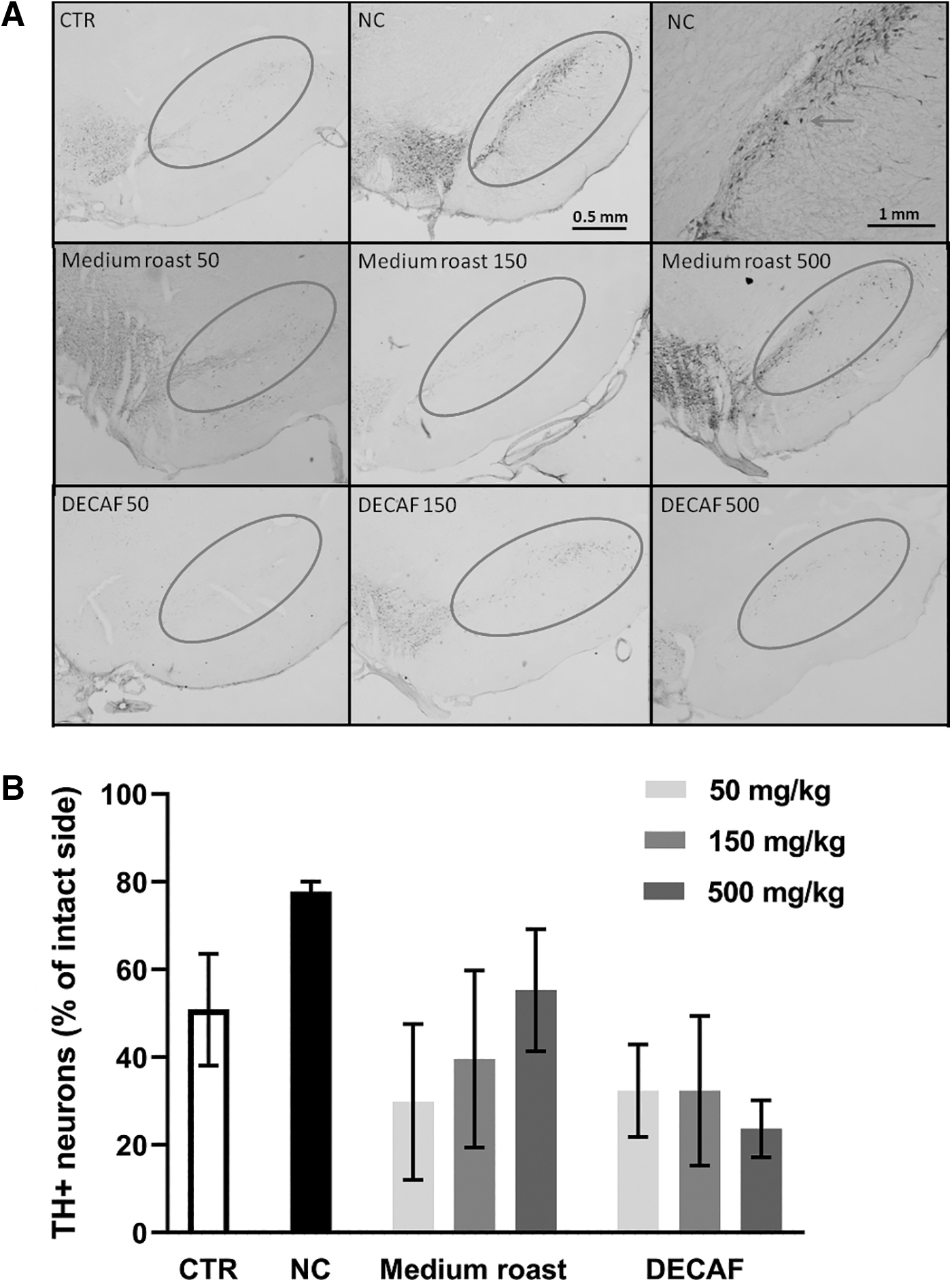
TH positive cells marked in immunohistochemistry.
Determination of monoamine levels by HPLC
There was no difference among the groups as the content of the monoamines and metabolites analyzed in the nonlesioned striatum (Fig. 5). Kruskal–Wallis test revealed significant difference among the groups in the content of DA [H(7, 65) = 14.79; P = .0388] and noradrenaline [H(7, 65) = 14.66; P = .0406] on the right striatum (lesioned side), but no differences were found on posttest comparisons. On the other hand, the infusion of 6-OHDA induced reduction of DA and their metabolites dihydroxyphenylacetic acid (DOPAC) and homovanillic acid on the lesioned side compared to the nonlesioned side (intragroup comparison) in several groups (Fig. 5A, C, D). There was no difference on noradrenaline (Fig. 5B), serotonin (Fig. 5E), and 5-hydroxyindolacetic acid (Fig. 5F) levels between lesioned and nonlesioned striatum in any group.
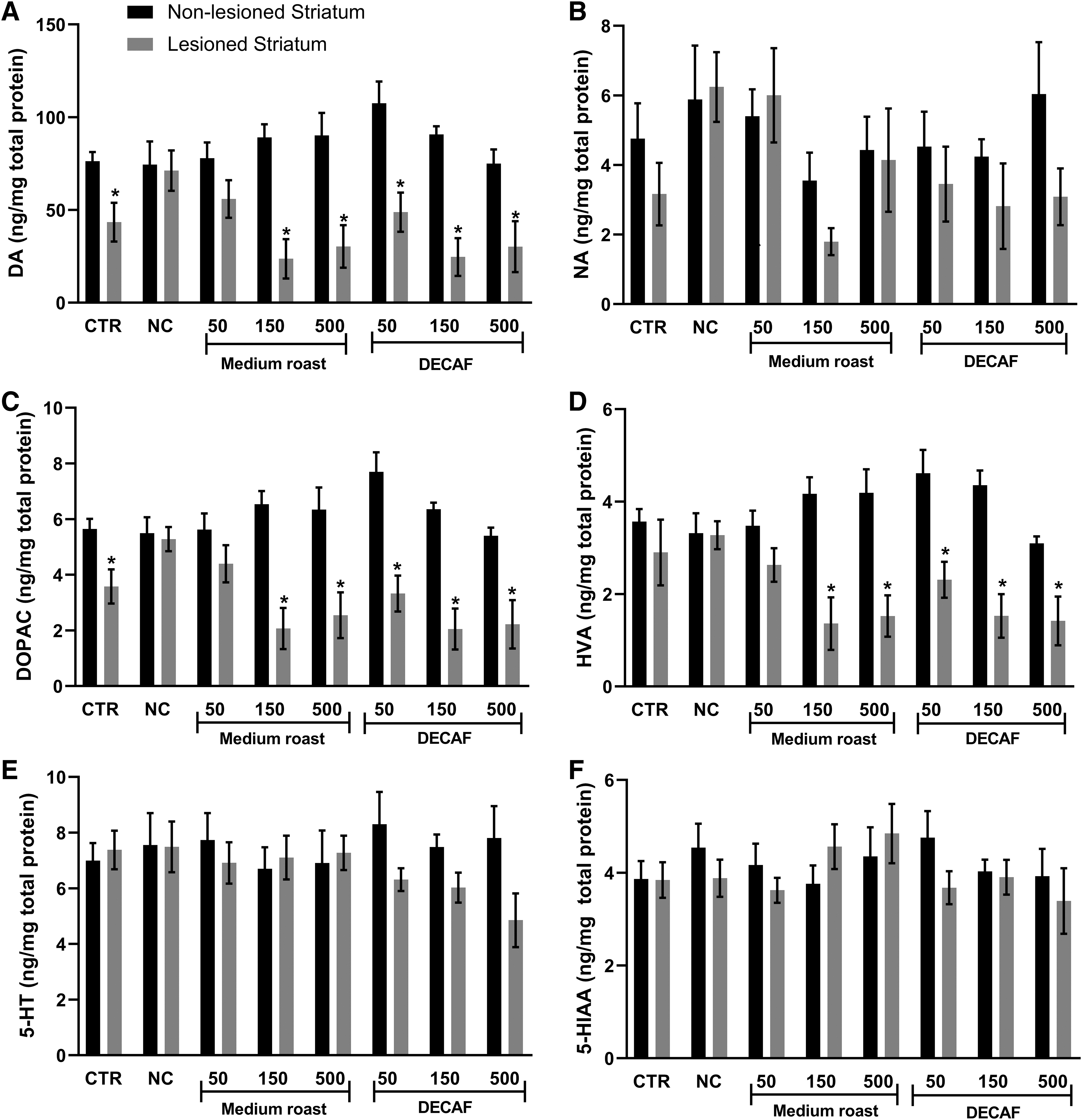
Levels of monoamines and their metabolites (ng/mg of total protein) in the left (nonlesioned) and right (lesioned) striatum of rats with unilateral lesion of the nigrostriatal pathway treated orally with water (CTR, control), 50, 150, and 500 mg/kg of medium roast or DECAF coffee extracts. The neurotoxin 6-OHDA was injected in the right MFB of the animals, except on NC, injected with saline.
Discussion
Observational studies and meta-analysis suggest a positive effect of coffee and other caffeinated beverages in neurological conditions, such as PD and AD. 3 –5,16,17 Experimental studies support the neuroprotective role of caffeine and other coffee constituents in biochemical and animal models. 18 –25 Our study assessed the potential prophylactic/therapeutic effects of instant coffee extracts on PD by different approaches, including antioxidant activity, pro-dopaminergic activity, and neuroprotection, as well as their inhibitory effect on AchE, which may be relevant for cognitive functions both in PD and AD.
First, we considered the influence of the roasting process and caffeine content in two in vitro antioxidant assays. The four coffee samples used in our study presented slight differences in the phytochemical profile, with the light roast and dark roast coffees having higher concentrations of caffeine, while decaffeinated coffee had only trace amounts. Dark roast coffee presented the strongest DPPH scavenging activity, while the medium roast coffee presented the strongest antioxidant effect in the lipid peroxidation assay. The present data corroborates previous studies that show that the antioxidant activity in coffee is not related to the roasting degree, but mainly with the phenolic content. The origin, harvesting, processing, and preparation of the coffee extracts influence the content of caffeine, chlorogenic acids, melanoidins, and many other constituents and as result the total antioxidant capacity. 26 –28
Besides acting as an adenosinergic inhibitor, caffeine has also shown AchE inhibition, 28 –30 which could make this compound relevant to AD therapy considering that cholinesterase inhibitors are the main class of drugs used in the treatment of AD. The three samples of caffeinated coffee inhibited the AchE in vitro, which the order of potency (IC50) related to the caffeine concentration. This result suggests that caffeine is important for AchE inhibition, although other compounds in decaffeinated coffee also exert an inhibitory effect on AchE, because decaffeinated coffee also showed AchE inhibition in higher concentrations.
A recent study observed a moderate correlation between the total phenolic content and AchE inhibition in extracts of roasted coffee, confirming that caffeine is the most active compound, but other components may contribute to the final effect. 30 AchE inhibitors reduce neuroinflammatory responses and neurodegeneration in vitro and in vivo, suggesting these drugs to be a valuable therapeutic agent for neuroinflammation-associated diseases 31,32 ; however, whether the concentration of caffeine after regular use of coffee really contribute for an in vivo protective activity is still matter of debate.
The possible therapeutic effect of caffeine on PD is mainly attributed to its antagonism on the adenosinergic system, which in turn modulates the dopaminergic system. 9 Our results on the catalepsy test suggest that the presence of caffeine is essential to produce behavioral changes. The two caffeinated coffee samples tested were able to inhibit the cataleptic status induced by haloperidol in a dose-dependent way, while the decaffeinated coffee had no effect. Several adenosinergic antagonists, including caffeine, have shown effect at this animal model. 10,33,34
To better investigate the coffee potential as neuroprotective agent in PD, we used the unilateral 6-OHDA lesion model. Coffee treatment began 30 days before the infusion of 6-OHDA and it continued for another 15 days. Thus, we could assess whether the previous coffee administration would exert some neuroprotection (its prophylactic effect), as it is supposed to occur in humans. The highest dose of medium roast coffee was able to decrease significantly the number of rotations induced by methamphetamine, which promotes ipsilateral rotation due to greater release of DA and reuptake blockade in the intact side. This dose of coffee also reduced the apomorphine-induced contralateral rotations in 69%, although it was not statistically significant. None of the other treatments changed the rotational behavior of rats after methamphetamine or apomorphine challenge.
The treatment with medium roast and decaffeinated coffee did not avoid the loss of dopaminergic neurons, but medium roast coffee at 500 mg/kg (the dose which reduced the metamphetamine-induced ipsilateral rotations) showed 22% more TH-positive neurons on the damaged side than the control group. The 6-OHDA lesion also induced diminished levels of DA and its metabolite DOPAC in the lesioned side compared to the intact side in control animals and in the groups medium roast (150 and 500 mg/kg) and decaffeinated coffee (50–500 mg/kg). Interestingly, rats treated with the smallest dose of medium roast coffee (50 mg/kg) presented similar levels of DA and their metabolites in the striatum of both sides. Previous studies have observed a neuroprotective effect of caffeine in the 6-OHDA experimental model. 19,20,25
Other studies employing different neurotoxins and protocols also suggested the neuroprotective effect of caffeine in PD models. 18,22,35,36 In addition, many other coffee components have shown neuroprotective effects in in vitro assays and experimental models of PD. Among them, it is worth mentioning the chlorogenic acid, 21 caffeic acid, 24 and eicosanoyl-5-hydroxytryptamide. 22,23
In brief, our study brings additional evidence of the pro-dopaminergic effect of coffee and its potential to the treatment or prevention of PD. The observed blockade of haloperidol-induced catalepsy and the decrease in the number of rotations in 6-OHDA lesion model suggests that coffee may indeed modulate the dopaminergic system. The antioxidant capacity observed suggests that coffee compounds are important to prevent the accumulation of reactive oxygen species and to inhibit cell damages, while the AchE inhibitory activity can contribute to attenuate cognitive deterioration in PD and AD patients. Taking together, the data obtained in this study support the assumption that coffee exerts moderate pro-cholinergic and pro-dopaminergic effects and caffeine seems to be the main factor responsible for these effects, although the contribution of other constituents cannot be rejected.
Footnotes
Acknowledgments
The authors are thankful to Dr. Joaquim Maurício Duarte-Almeida, Ricardo Borges Machado and Guilherme Cunha Ribeiro for their assistance with some techniques used in this study. This study is dedicated to the memory of Professor E.A.C.
Authors' Contributions
D.O.K., G.R.M., and L.I.G.d.P.-F. contributed to the acquisition, analysis, and interpretation of data, draft and revising the manuscript. G.N. contributed to the caffeine quantification, analysis and interpretation of data, draft and revising the manuscript. E.A.C. and F.R.M. contributed to the conception and design, analysis an interpretation of data, draft, and revising the manuscript.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
D.O.K. received scholarship from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP #2009/13305-9). Associação Fundo de Incentivo à Pesquisa (AFIP) provided some facilities.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.