
Abstract
The transport of cations in the cardiomyocytes, crucial for the functioning of the heart, can be affected by walnut diet due to the high content of polyunsaturated fatty acids. Healthy and metabolically compromised rats (drinking 10% fructose solution) were subjected to a diet supplemented with 2.4 g of walnuts for 6 weeks to investigate the effect on proteins involved in cation transport in the heart cells. Fructose increased the level of the α1 subunit of Na+/K+-ATPase and the phosphorylation of extracellular signal-regulated kinase 1/2 in the heart of control and walnut-eating rats, while elevated L-type calcium channel α (LTCCα), sodium–calcium exchanger 1 (NCX1), and Maxi Kα level were observed only in rats that did not consume walnuts. However, walnuts significantly increased the cardiac content of LTCC, NCX1, and Maxi Kα, as well as Kir6.1 and SUR2B subunits of KATP channel, but only in fructose-naive rats. In animals that drank fructose, a significant increasing effect of walnuts was observed only in Akt kinase phosphorylation, which may be a part of the antiarrhythmic mechanism of decreasing cation currents in cardiomyocytes. The walnut diet-induced increase in LTCC and NCX1 expression in healthy rats may indicate intense cardiac calcium turnover, whereas the effect on Kir6.1 and SUR2B subunits suggests stimulation of KATP channel transport in the cardiac vasculature. The effects of walnuts on the cation-handling proteins in the heart, mostly limited to healthy animals, suggest the possible use of a walnut-supplemented diet in the prevention rather than the treatment of cardiological channelopathies.
INTRODUCTION
Diets rich in essential fatty acids (FAs), or purified essential FA administration, have anti-inflammatory, vasodilatory, antiarrhythmic, antithrombotic, antioxidant, and antiatherogenic effects. 1 In the areas where the seafood is not abundant in a diet, walnuts are an important source of essential polyunsaturated fatty acids (PUFAs) such as omega-6 linoleic acid and omega-3 α-linolenic acid (ALA). In addition, walnuts contain other biologically active components important for cardiovascular health, such as polyphenols, minerals, and vitamins. 2
The 2013 guidelines of the American College of Cardiology Foundation/American Heart Association for the management of heart failure recommend the consumption of omega-3 PUFAs. 3 We recently detected that a walnut-rich diet increases FA metabolism in the heart and, in rats with metabolic disturbances, redirects cardiac metabolism from saturated and monounsaturated FA to omega-3 PUFA. 4 Opposite of the many well-documented harmful effects of saturated palmitic acid (PA) on the heart, 5 –7 the products of the ALA exert favorable metabolic effects in the heart. 8
Particular emphasis is on the anti-inflammatory effects of eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), which may be directly related to their metabolism, and antiarrhythmic effects targeting proteins involved in membrane ion transport. 9 After entering the cells, omega-3 PUFAs incorporate into the membranes, performing important functions, including regulating ion channels. Omega-3 PUFAs have an antiarrhythmic effect by inhibiting voltage-dependent sodium and calcium currents in cardiomyocytes. 10
Some channels/exchangers/pumps for sodium, potassium, and calcium, such as the L-type calcium channel (LTCC) and Kv4.2 potassium channel, are directly involved in generating the action potential in cardiomyocytes. Others, such as the Na+/K+ pump and the Na+/Ca2+ exchanger (NCX1), are responsible for maintaining ion homeostasis in cardiomyocytes. 11 In addition, some ion channels, such as KATP, connect metabolism and excitation/contraction processes. This type of channel responds to changes in ATP concentration and modulates action potential-related repolarization. 12 Maxi K channel is a large-conductance Ca2+- and voltage-activated K+ channel, whose opening in the mitochondria of cardiac cells favorably affects respiratory function. 13
In addition, some of the protein kinases involved in the regulation of metabolism, proliferation, and cell death in the heart, such as Akt and extracellular signal-regulated kinase 1/2 (ERK1/2), are also involved in the regulation of the action potential through the modification of ion channels or in a channel-independent way. 14
Increased cardiac sodium and calcium currents are often associated with arrhythmias, so agents that reduce these currents, such as EPA and DHA, have potential antiarrhythmic effects on the heart. 9 As we demonstrated before, walnut consumption in healthy and metabolically compromised rats led to increased levels of ALA, EPA, and docosapentaenoic acid while decreased PA level and its conversion to palmitoleic acid in the heart. 4
Therefore, this study was designed to assess the relationship between FA intake and subsequent metabolic impact in the hearts of walnut-fed healthy and metabolically compromised rats, with the expression of proteins involved in the handling of calcium, sodium, and potassium ions responsible for the cardiac electrical and contractile activities. Protein kinases that may mediate the effects of omega-3 PUFA on ion channels were also included in the study.
MATERIALS AND METHODS
Chemicals
Fructose was purchased from Omnia Nisasta Sanayi ve Ticaret A.S. (Adana, Turkey). Antibodies raised against LTCCα, Kv4.2, Kir6.1, Kir6.2, SUR2B, Maxi Kα, α1, and α2 subunits of Na+/K+-ATPase, NCX1, p-Akt Ser473, Akt, and β-actin, as well as secondary horseradish peroxidase (HRP)-conjugated antirabbit antibodies, were obtained from Santa Cruz Biotechnology, Inc., (Heidelberg, Germany). Antibodies recognizing p-ERK1/2 Thr202/Tyr204 and ERK1/2 were products of Cell Signaling Technology (Danvers, MA, USA).
Antimouse secondary antibody was a product of R&D Systems, Inc., (Minneapolis, MN, USA). Reagents for the bicinchoninic acid (BCA) assay were purchased from SERVA Electrophoresis GmbH (Heidelberg, Germany). Electrophoretic reagents were products of Sigma-Aldrich Corporation (St. Louis, MO, USA).
Animals and treatments
Walnuts (Juglans regia) used in this study were obtained from a local market, and the contents of the walnuts are described before. 2 Owing to the PUFA susceptibility to oxidation when provided as part of the standard animal consumption, we distributed the whole walnut kernels to the experimental animals. We strictly monitored them to ensure every animal ate its daily walnut portion at the moment of the distribution.
As we reported before, 15 36 male Wistar rats, 21 days old, were randomly divided into two experimental groups: the control (C) and fructose-rich diet (F) group. All rats were fed ad libitum a standard laboratory food, but instead of tap water, F group animals drank tap water supplemented with 10% fructose. As we reported earlier, after 9 weeks of intake of 10% fructose solution, male rats developed symptoms of metabolic syndrome. 16 –19 Thus, after 9 weeks, both groups were additionally divided into two subgroups. Diet for half of C and half of F rats was supplemented with 2.4 g of walnut kernels per day each during the next 6 weeks (W and FW group, respectively). Rats in the F group continued to drink the 10% fructose solution during the walnut treatment. The experimental design is shown in Figure 1.
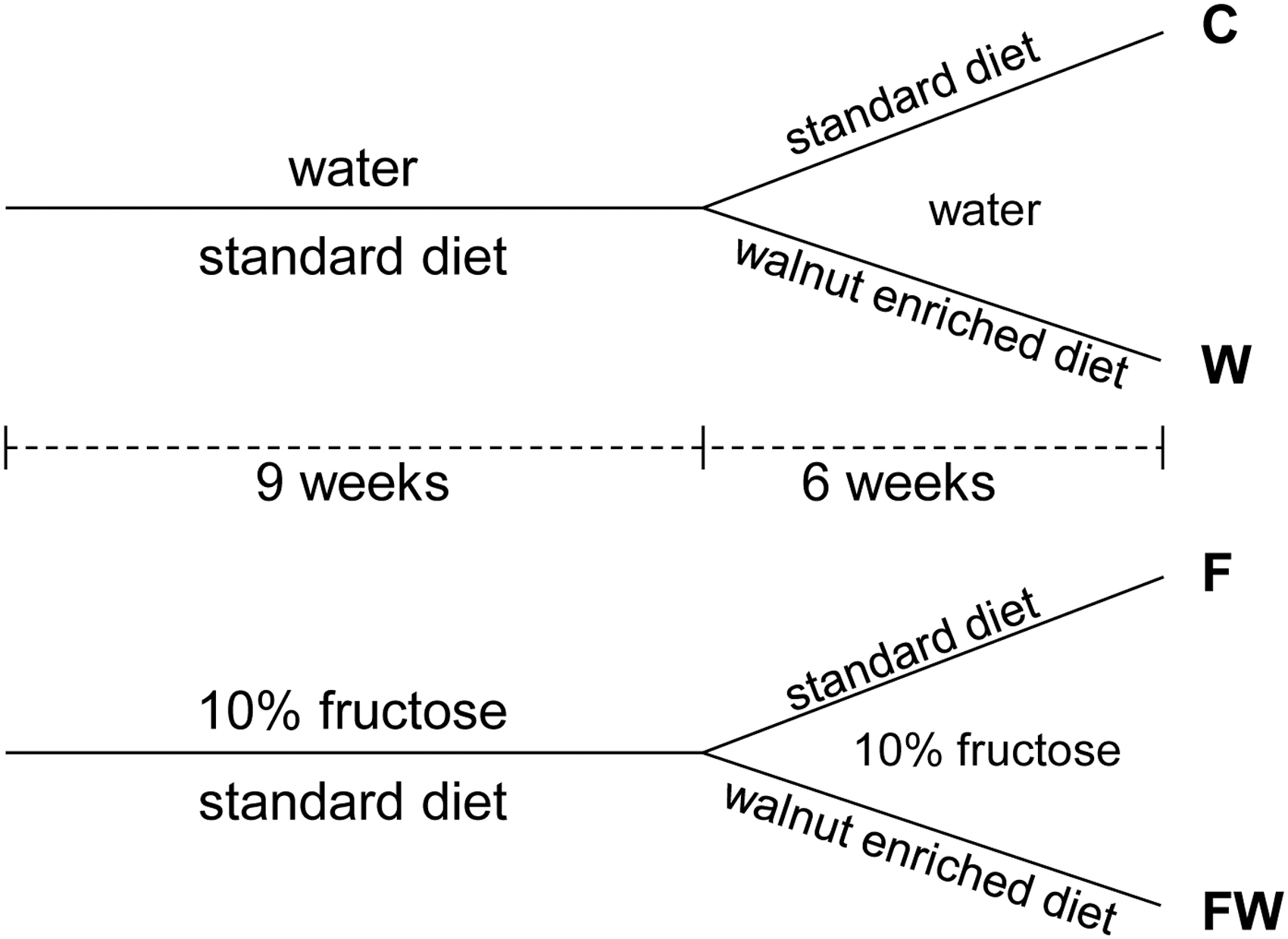
Experimental design. C: control; W: walnuts; F: fructose; FW: fructose+walnuts.
The selected amount of walnuts resembles a 56 g/day dose in humans, previously shown to exert relevant cardiometabolic effects. 2,20,21 The human equivalent dose was calculated according to the method by Reagan-Shaw and coworkers, 22 taking into account between-species metabolic differences. After the animals were sacrificed, the hearts were isolated and stored at −80°C.
All animal procedures and protocols were conducted in accordance with the EU Directive (2010/63/EU) on the protection of animals used for experimental and other scientific purposes and were approved by the official Vinča Institute's Ethical Committee for Experimental Animals.
Cell lysate preparation
Heart samples were homogenized for 3 × 30 s at 4°C with an Ultra-Turrax in modified RIPA buffer (50 mmol/L Tris-HCl, pH 7.4, 150 mmol/L NaCl, 1% Triton X-100, 0.2% Na-deoxycholate, 0.2% sodium dodecyl sulfate (SDS), 1 mmol/L ethylenediaminetetraacetic acid, protease inhibitors, phosphatase inhibitors). The homogenate was centrifuged for 30 min at 15,000 g, and the supernatant was referred to as cell lysate. After protein concentration assessment by the BCA method, samples were prepared for electrophoresis.
SDS polyacrylamide electrophoresis and Western blot
Proteins were separated on 7.5 or 10% SDS polyacrylamide gels and transferred to polyvinylidene difluoride membranes. Membranes were blocked with 5% BSA and blotted with an antibody against LTCCα, Kv4.2, Kir6.1, Kir6.2, SUR2B, Maxi Kα, α1, and α2 subunits of Na+/K+-ATPase, NCX1, p-Akt Ser473, or p-ERK1/2 Thr202/Tyr204. After extensive washing, membranes were incubated with the secondary HRP-conjugated antibody and used for detection with enhanced chemiluminescent substrates. Membranes blotted with p-Akt and p-ERK1/2 were stripped and reprobed with the Akt and ERK1/2 antibodies, respectively. Blots were stripped and probed with the β-actin antibody to ensure equal protein loading in the lysate. Films were scanned and analyzed using ImageJ software (NIH, USA).
Statistics
All data are presented as means ± standard deviation (n = 9 per group). Statistical comparisons were performed by two-way analysis of variance with a Tukey's test for post hoc comparison using STATISTICA software (StatSoft, Inc., Tulsa, OK, USA). A value of P < .05 was considered statistically significant.
RESULTS
Voltage-gated channels for potassium and calcium
No significant changes in the level of voltage-gated potassium Kv4.2 channel in the heart of rats were observed either under the action of walnuts or under the influence of fructose (Fig. 2A). However, analyzing the results of Maxi Kα expression (Fig. 2B), we noticed that there were interactions between walnuts and fructose effects (P < .001). The results of the post hoc test showed that compared with the control group, the level of Maxi Kα was significantly increased in animals that ate walnuts, as well as those that drank fructose, but also in rats exposed to a combined diet (P < .01 W vs. C, P < .001 F vs. C and P < .01 FW vs. C).
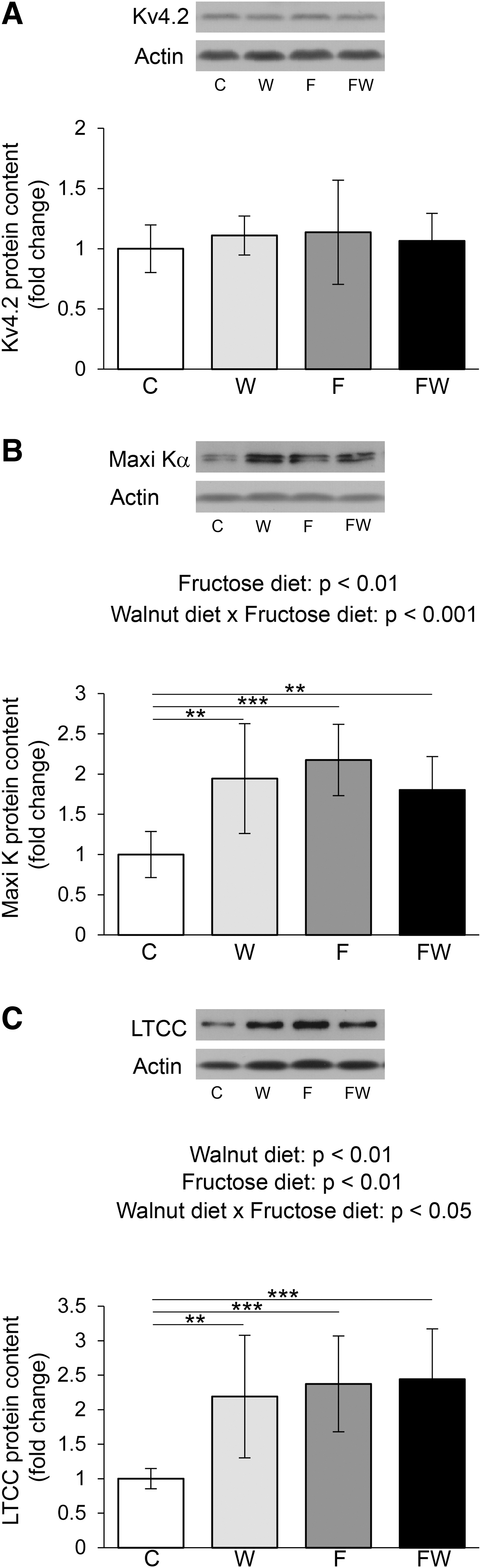
Effects of walnut supplementation on cardiac voltage-gated potassium and calcium channels in fructose-naive and fructose-fed rats. Levels of Kv4.2
The α subunit expression of the voltage-gated calcium channel LTCC was affected by interactions of two factors (P < .05) (Fig. 2C). According to the post hoc test, upon both a walnut-supplemented diet and 10% fructose, rats presented with higher levels of LTCCα in the heart than control rats (P < .01 W vs. C, P < .001 F vs. C). Also, the combined diet (fructose and walnuts) caused an increase in the level of this protein compared with the control (P < .001 FW vs. C).
ATP-regulated channels for potassium
Walnuts and fructose interacted in the regulation of Kir6.1 KATP channel subunit expression (P < .01) (Fig. 3A). Tukey's test indicated that the level of Kir6.1 protein was significantly increased in the heart cells of rats whose diet was supplemented with walnuts, compared with rats on a standard diet (P < .01 W vs. C). The reduction in Kir6.1 expression in rats that drank fructose and ate walnuts compared with animals that ate walnuts alone was close to the significance limit (P = .08 FW vs. W).
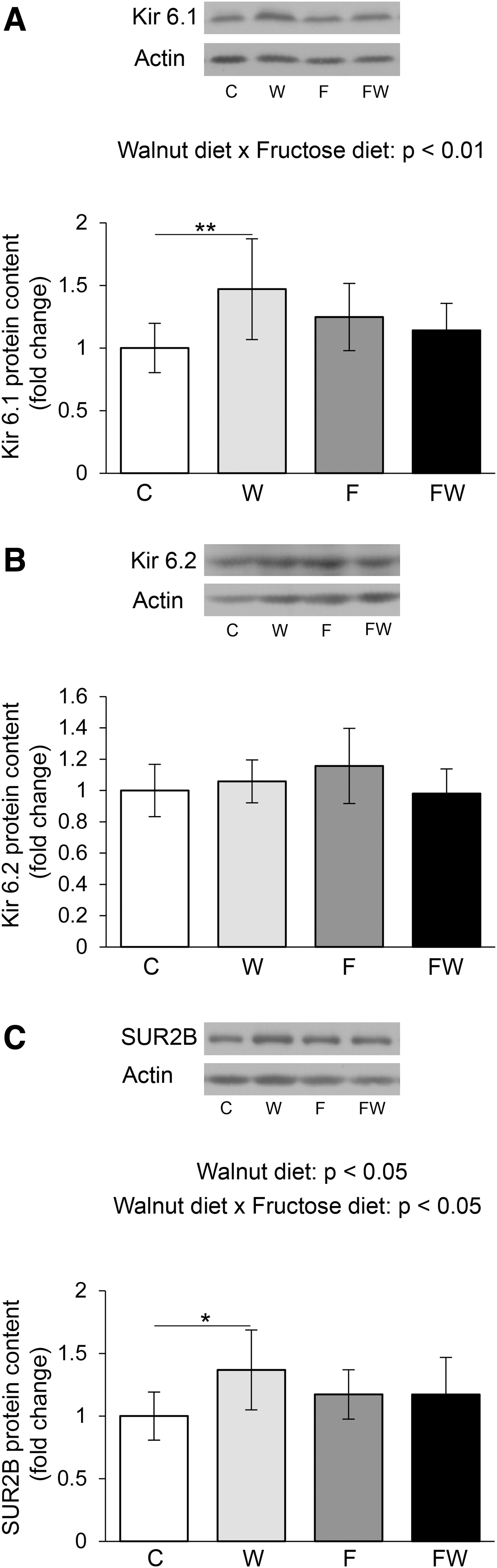
ATP-sensitive potassium channels in the heart of healthy and metabolically compromised rats on the walnut-rich diet. Levels of Kir6.1
Unlike the Kir6.1 subunit, neither walnuts nor fructose had a significant effect on the Kir6.2 subunit of KATP in rat heart cells, although the tendency of interactions of walnuts and fructose in the regulation of Kir6.2 was observed (P = .06) (Fig. 3B). However, regarding SUR2B expression, interactions between walnuts and fructose were observed (P < .05) (Fig. 3C). Tukey's test showed that the level of SUR2B was significantly higher in animals that consumed walnuts than controls (P < .05 W vs. C).
Sodium–potassium pump and NCX1
Regarding the sodium–potassium pump that maintains the ionic balance in the heart cells between the two contractions, the main effect of fructose was observed. Namely, the α1 subunit was significantly increased under the action of fructose (P < .01) (Fig. 4A). The trend of interactions of walnuts and fructose in the regulation of α1 was also observed (P = .06). Fructose and walnuts did not affect the level of the α2 subunit in the heart, although walnuts tended to decrease the α2 level (P = .08) (Fig. 4B).
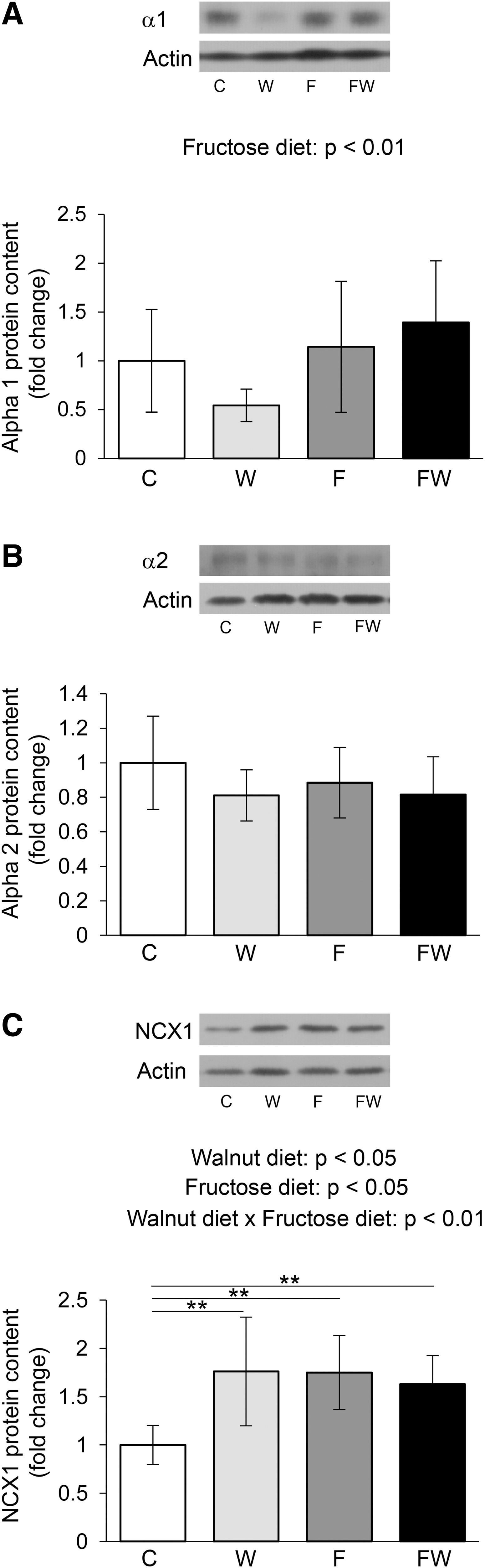
Effects of walnut consumption on Na+/K+-ATPase and Na+/Ca2+ exchanger in the heart of fructose-naive and fructose-fed rats. Levels of α1
Regarding the level of NCX1 transporter in cardiac cells, interactions between walnuts and fructose were observed (P < .01) (Fig. 4C). The post hoc test showed that, compared with the control group, NCX1 expression was significantly increased in animals that ate walnuts, as well as in those that drank fructose, but also in those that combined these two types of diet (P < .01 for all; W vs. C, F vs. C and FW vs. C).
Akt and ERK1/2 protein kinases
The expression of Akt kinase in the heart was not altered under the influence of applied dietary treatments, although walnuts tended to decrease the level of Akt (P = .06) (Fig. 5A). However, interactions of walnut and fructose in the regulation of Akt phosphorylation have been observed (P < .01) (Fig. 5B). In animals that drank fructose, walnuts significantly increased Ser473 phosphorylation level compared with those that drank fructose only (P < .01 FW vs. F).
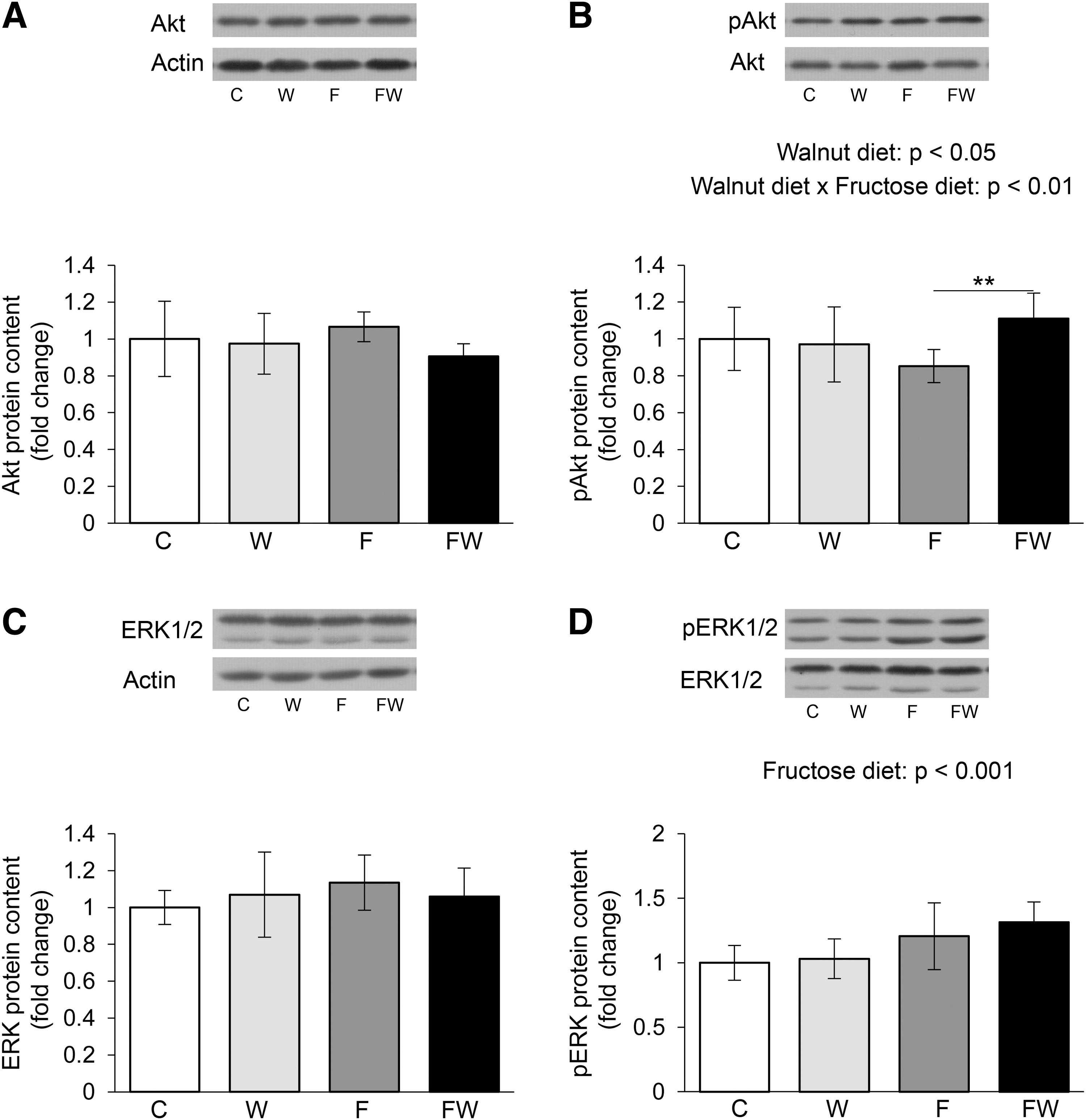
Phosphorylation of Akt and ERK1/2 protein kinases in the heart of healthy and metabolically compromised rats subjected to walnut-rich diet. Levels of p-Akt
Dietary supplementation with walnuts did not affect the expression of ERK1/2 kinase in the heart, nor did it affect the increased intake of fructose (Fig. 5C). However, fructose expressed the effect on ERK phosphorylation. Namely, fructose significantly increased the phosphorylation of this kinase at Thr202/Tyr204 (P < .001) (Fig. 5D).
DISCUSSION
Although research on the antiarrhythmic potential of omega-3 PUFAs has been going on for years, supporting the idea that they can be used in the treatment of heart rhythm disorders, some studies deny this effect, and others even testify to the proarrhythmic effects of EPA and DHA. 23,24 Considering the FA composition of walnuts, 2 as well as the cardiac FA profile in walnut-fed rats, 4 and knowing the cardiac effects of omega-3 PUFA on ion transport, 1 we assumed that a walnut-rich diet affects ion channels in this tissue beneficially. The assumption was additionally supported by the results of our recently published study, showing that walnut consumption affects the expression of ion channels/exchangers/transporters involved in the regulation of vascular tone in aortic tissue. 25
As we recently reported, rats from this study exposed to the fructose-rich diet consumed more liquid and less food, and total energy intake was increased. A diet supplemented with walnuts did not affect food and fluid intake but resulted in an increase in energy intake. 25
Increase in body weight, visceral fat mass, insulinemia, and deterioration of the lipid profile clearly indicate that 15-week treatment with 10% fructose led to the development of a cluster of symptoms defined as metabolic syndrome. 15,25 Neither diet changed the blood glucose level of the rats. It should be noted that the diet supplemented with walnuts led to an increase in the body weight of the rats and visceral adipose tissue, and caused a significant increase in insulinemia. 25 Also, surprisingly, a diet rich in walnuts did not change the parameters of the lipid profile. 15 In contrast, the intake of walnuts was associated with favorable changes in the FA profile in the plasma and heart. 4,15
In this study, we found that a diet rich in fructose, which resulted in the development of metabolic syndrome phenotype, 15,25 increased the level of the α1 subunit of Na+/K+-ATPase and the phosphorylation of ERK1/2 kinase in the heart of rats, regardless of whether they ate walnuts. We recently reported that consumption of 10% fructose for 9 weeks also increased the levels of α1 and α2 subunits of Na+/K+-ATPase in the heart of rats. 26 Increased fructose intake also elevated the levels of LTCCα, Maxi Kα, and NCX1 in the heart of walnut-naive rats, but a diet high in walnuts abolished all these effects.
As suggested by Lee et al., 27 the activation of the Na+/K+ pump, indicated in fructose-fed rats in our study due to a rise in α1 protein level, can activate LTCC through the activation of ERK1/2. Furthermore, the increased flow of Ca2+ through the LTCC channel can stimulate the NCX1 exchanger and the calcium-dependent Maxi K channel.
Walnuts increased the content of LTCCα, NCX1, Maxi Kα, Kir 6.1, and SUR2B, but only in the heart of fructose-naive rats. In animals that drank fructose, a significant positive effect of walnuts was observed only in the phosphorylation of Akt kinase. According to Catalucci et al., 28 Akt regulates L-type Ca2+ channel activity by modulating Cav α1 protein stability. However, that mechanism probably cannot be attributed to a walnut-enriched diet because the effect of walnuts on Akt is observed only in metabolically compromised rats, whereas the effect on LTCCα exists only in the heart of healthy walnut-fed rats.
Increased expression of LTCCα and NCX1 in fructose-naive animals eating walnuts may indicate that a walnut-rich diet intensifies calcium turnover during the cardiac contraction/relaxation cycle. Namely, if expression correlates with the function of the calcium-handling proteins, walnuts probably stimulate calcium entry into heart cells in the contraction phase (via increased LTCCα) and calcium excretion from the heart cells in the relaxation phase (due to increased NCX1). 29 However, the question is whether the increased expression of ion channels such as LTCC, observed in fructose-naive animals that consumed walnuts, indicates a proarrhythmic effect.
The controversy concerning the expected reduction in ionic currents related to the increased level of omega-3 PUFA in the heart of walnut-consuming rats 4 versus the observed increase in calcium channel and transporter expression may be hypothetically explained by the model proposed by Ballou et al., 30 which includes different outcomes of phosphoinositide 3-kinase/Akt signaling pathway activation. In the case of Nav1.5 channels, they indicated that Akt activation, in our experiment associated with the action of walnuts in fructose-fed rats, leads to phosphorylation of Nav1.5 and suppression of sodium current.
In contrast, activation of Akt resulted in phosphorylation and deactivation of forkhead box protein O1 and increased gene expression of the α subunit Nav1.5. In short, increased ion channel expression and decreased ion current can go together.
Given that PA induced Ca2+ and Na+ influxes in cardiomyocytes, 31 the reduction of PA level and its conversion in the heart of walnut-eating animals 4 could be another mechanism of beneficial effects of walnuts on cation-handling proteins, particularly those involved in Ca2+ and Na+ transport. Increased PA level impaired cellular Ca2+ handling and contraction in normal cardiomyocytes, whereas, in contrast, high PA level was shown to be vital to sustaining cardiac Ca2+ handling and contraction in insulin-resistant conditions. 32
PA treatment also shortened action potential duration, whereas voltage-gated ion channels in left atrial myocytes showed a reduction of the L-type calcium current and the transient outward potassium current. 33 Nevertheless, in our study, we observed that the high content of PA in the heart of rats with increased intake of fructose 4 coincides with an increased level of LTCC, supporting Fauconnier's 32 observation regarding the insulin resistance. In addition, palmitate induced contractile dysfunction via a defect in calcium-induced calcium release through endoplasmic reticulum ryanodine receptor 2 34 and decreased the rate of cardiomyocyte relaxation via sarcoplasmic/endoplasmic reticulum Ca2+-ATPase 2a activity modification. 35
Although increased production of long-chain acylcarnitines is associated with a reduced level of Ca2+ in the cytoplasm, 36 the intense cardiac β-oxidation that we observed accompanied by increased intake of fructose or a diet enriched with walnuts 4 was associated with an increased level of LTCC and NCX1 in both treatments. Thus, increased acylcarnitine production coupled with intense FA oxidation may explain the increase in the level of NCX1 removing calcium from the cytoplasm, but not the elevated level of LTCC acting the opposite.
Considering that fructose transporters and enzymes of fructose metabolism have been detected in heart cells and as fructose metabolism is a big consumer of ATP, 37 it can be expected that increased fructose intake affects cardiac KATP. However, no effect of fructose was observed on any of the KATP subunits. As reviewed by Ye et al., 38 KATP channels are a class of potassium channels inhibited by intracellular ATP increase. They are composed of Kir6.x subunits and sulfonylurea receptors (SURs). Kir6.2/SUR2A is present in ventricular muscle cells, whereas Kir6.1/SUR2B is present in vascular smooth muscle cells. 38
Under pathophysiological conditions, in particular, KATP channels play an essential protective role by regulating cardiac repolarization. In this light, the increase in SUR2B and Kir 6.1 levels observed in the hearts of fructose-naive rats eating walnuts can be considered beneficial for cardiac vasculature.
Maxi K channels are characterized by a large conductance to K+ and their sensitivity to voltage and Ca2+. Maxi K channels are expressed at the plasma membrane, with adult cardiomyocytes being an interesting exception. Specifically, in the mitochondria of the cardiac cells, the opening of the Maxi Kα channel improves the respiratory function of the mitochondria and protects the heart from ischemic events. 13 Interestingly, we observed an increase in the expression of the Maxi Kα subunit under the individual action of both fructose and walnuts. It can be related to the data reported by Dopico et al. 39 that both PA, which was increased in the heart under the influence of a fructose-rich diet, and DHA, which was increased in the heart of rats that ate walnuts, 4 stimulate Maxi K activity.
As a limitation of this study, measurement of ion currents, as a functional parameter, would be required to conclude whether the expression of ion transporters/exchangers/pumps correlates with their function. Another study limitation is that protein expression was measured in the whole heart extract. Namely, individual ion channels have specific roles in different heart cells, and additional studies on isolated cardiomyocytes from walnut-fed experimental animals would complement the present study.
CONCLUSIONS
Numerous effects of the walnut diet on cation-handling proteins in the heart of healthy animals were observed. Those concerning ion channels for potassium (KATP and Maxi K) can be interpreted as beneficial for the heart. In contrast, the effects of walnuts on calcium channels/exchangers (LTCC and NCX1) suggest intensive calcium traffic through heart cells, but it is difficult to predict the physiological consequences. Contrary to our initial hypothesis, the positive effect of walnuts on the cardiac metabolism of FA that we recently reported 4 was not reflected in the expression of cation-handling proteins in the heart of metabolically compromised animals.
Furthermore, the expression of ion channels in the heart more closely mirrors the pattern of changes in β-oxidation regulators than the cardiac FA profile in rats consuming fructose and/or walnuts. 4 If we want to send a message regarding our eating habits, we would say that eating walnuts for a snack can be beneficial for a complex network of ion-handling proteins in a healthy heart but that moderate walnut consumption has a limited capacity to restore cardiac cation transport in metabolic syndrome.
Footnotes
AUTHORs' CONTRIBUTIONS
S.T. contributed to investigation, formal analysis, and visualization; S.R. was involved in investigation and formal analysis; M.Z. carried out conceptualization, methodology, and writing—review and editing; T.C. and M.S. took care of investigation and writing—review and editing; T.I., M.P., M.K., and J.S. contributed to investigation; and G.K. was in charge of conceptualization, methodology, and writing—original draft.
DATA AVAILABILITY STATEMENTS
The data that support the findings of this study are available on request from the corresponding author.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This article was supported by the Ministry of Education, Science and Technological Development, Republic of Serbia (Grant No.: 451-03-9/2021-14/200017 and No.: 451-03-9/2021-14/200015).