
Abstract
In this study, we evaluated the effects of Lactobacillus reuteri NCIMB (LRC™) supplementation on hypercholesterolemia by researching its effects on cellular cholesterol metabolism in hypercholesterolemic rats (KHGASP-22-170) and HepG2 cell line. Rats were separated into six groups after adaptation and were then fed a normal control (NC), a high-cholesterol diet (HC), or a HC supplemented with simvastatin 15 mg/kg body weight (positive control [PC]), LRC 1 × 109 colony-forming units (CFU)/rat/day, LRC 4 × 109 CFU/rat/day, or LRC 1 × 1010 CFU/rat/day (1 × 109, 4 × 109, or 1 × 1010). The rats were dissected to study the effects of LRC on cholesterol metabolism and intestinal excretion at the end of experimental period. We discovered that LRC mainly participated in the restraint of 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase, the uptake of low-density lipoprotein (LDL) cholesterol into tissues, partially in the transport of cholesteryl esters into high density lipoprotein for maturation, and intestinal excretion of cholesterol. These results are supported by the expression of transcription factors and enzymes such as HMG-CoA reductase, SREBP2, CYP7A1, CETP, and LCAT in both messenger RNA (mRNA) and protein levels in serum and hepatic tissue. Furthermore, the LRC treatment in HepG2 significantly reduced the mRNA expression of HMG-CoA reductase, SREBP2, and CEPT and significantly increased the mRNA expression of LDL-receptor, LCAT, and CYP7A1 at all doses. Hence, we suggest that LRC supplementation could alleviate the serum cholesterol level by inhibiting the intracellular cholesterol synthesis, and augmenting excretion of intestinal cholesterol.
INTRODUCTION
Cholesterol is necessary for maintaining the fluidity of cell membranes. It is also required in various biochemical systems, including the synthesis of steroid hormones, bile acids, and vitamin D. 1 –3 However, elevated levels of low-density lipoprotein (LDL), defined as hypercholesterolemia, are positively correlated with atherosclerotic cardiovascular diseases (CVDs). 4 It has been suggested that 40.5% of the U.S. population could have a certain type of CVD by 2030, which would become the leading cause of death by then. It has also been proposed that a 1% reduction in LDL cholesterol in the human serum decreases the risk of CVDs up to 3% over the lifetime of a person. 5,6 Therefore, normalization of cholesterol levels and maintaining appropriate cholesterol homeostasis are important for preventing CVDs and related vascular diseases.
Cholesterol homeostasis in cells is controlled by both exogenous (diet) and endogenous (biosynthesis) cholesterol. 7,8 LDL receptors on the membrane bind to LDL cholesterol and are taken up from the blood into the cells, providing cholesterol to the cells. Accumulation of cholesterol in the cells activates acyl-CoA cholesterol acyltransferase (ACAT) that converts cholesterol esters for storage. When cholesterol is stored excessively in the cell, the expression of LDL receptor is suppressed to reduce the uptake, leading to the buildup of cholesterol in the blood. 7 –9 Conversely, when the cell is deficient in cholesterol, the LDL receptor expression is promoted and biosynthesis of cholesterol gets activated by 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase, especially in the liver. Cholesterol is the precursor of bile acids in the liver. Thus, bile acid synthesis is a major pathway for the catabolism of hepatic cholesterol. Therefore, the liver is the major organ of cholesterol homeostasis, which is tightly regulated by biosynthesis and secretion. 7 –10
Many recent studies have attempted to treat hypercholesterolemia using natural cholesterol-lowering agents. The pharmacological effects of these agents are similar to that of drugs used to treat hypercholesterolemia, but they have fewer side effects. 11 It has recently been shown that lactic acid bacteria, also known as probiotics, have potential health-promoting advantages as they decrease both the total serum cholesterol and LDL cholesterol levels. Although it is not yet completely understood, the mechanism through which lactic acid bacteria lowers the serum cholesterol level has been proposed.
This mechanism includes the following steps: enzymatic deconjugation of bile salts, assimilation of cholesterol, microbial transformation of cholesterol to coprostanol, and cholesterol binding to the bacterial cell wall. 12 In addition, various randomized, double-blind clinical trials have demonstrated that the administration of lactic acid bacteria is helpful in lowing both the total and LDL cholesterol of serum/plasma. Interestingly, these results imply that the reduction of cholesterol depends on the origin and classes of lactic acid bacteria. 13 The present research investigates the antihypercholesterolemic effects of Lactobacillus reuteri NCIMB 30242 on cholesterol synthesis and excretion in rats fed high-cholesterol diet (HC).
MATERIALS AND METHODS
Preparation of materials
The strain used in this study, L. reuteri NCIMB 30242 (LRC™), was originally produced by Chr. Hansen (Hoersholm, Denmark) and obtained from Vixxol Corp. (Gunpo, South Korea) in the dried powder form at a cell density of ∼1 × 1011 colony-forming units (CFU)/g, which was adjusted to 1 × 1010 CFU/mL for laboratory work. Briefly, the strain is gram-positive and is an anaerobic bacteria identified by 16S ribosomal RNA (rRNA) sequencing and its metabolic and growth characteristics (original data were suggested by Chr. Hansen). The strain was kept in an airtight container at −20°C until the experiment.
Cell culture and treatment
The HepG2 cell line (ATCC HB-8065) was acquired from the American Type Culture Collection (Rockville, MD, USA) and cultured under appropriate conditions (5% CO2 and 37°C). The cells were grown in high-glucose Eagle's minimum essential medium (EMEM) supplemented with 10% fetal bovine serum, 1% penicillin/streptomycin, 1%
Animals and treatment
The Institutional Animal Care and Use Committee of Kyung Hee University approved the protocol (KHGASP-22-170) for animal studies. The animals were cared for according to the “Guidelines for Animal Experiments” established by the university. Sprague–Dawley rats (6-week-old, male) were purchased from SaeRon Bio, Inc. (Uiwang, Korea) and were housed in cages at appropriate temperature, light, humidity (22°C ± 2°C, about 50%, semidiurnal light on–off. Rats were separated into six groups after adaptation and were then fed a normal control (NC), a HC, or a HC supplemented with simvastatin 15 mg/kg body weight (positive control [PC]), LRC 1 × 1010 CFU/rat/day, LRC 4 × 109 CFU/rat/day, and LRC 1 × 1010 CFU/rat/day (1 × 109, 4 × 109, and 1 × 1010).
The rats in the NC control group were fed an AIN-93G rodent purified diet (Orient Bio) and were provided with water ad libitum. The rats in the diet-induced hypercholesterolemia group were fed an Atherogenic Rodent Diet D12336 (HC; Research Diet, Inc.) for 12 weeks. The diet was controlled by oral administration to the rats using a feeding needle. The body weight and the amount of consumed food by each group were measured twice per week for 12 weeks. After 12 weeks, the rats were anatomized to gather serum and tissues.
Histological observation
The aortas of the rats were fixed, embedded, sliced, and stained according to the hematoxylin and eosin analysis protocol. 14
Protein extraction and Western blot analysis
Protein extraction of liver tissues and cells were proceeded in accordance with previous study's methods. 14 HMG-CoA reductase, LDL receptor, SREBP2, CYP7A1, β-actin, and anti-rabbit IgG HRP-linked antibody antibodies were purchased from Cell Signaling Technology (Beverly, MA, USA).
Isolation of total RNA and real-time polymerase chain reaction
Total RNA was isolated from the livers and analyzed for the expression of genes, including HMG-CoA reductase, LDL receptor, SREBP2, CYP7A1, LCAT, CETP, and GAPDH, by real-time polymerase chain reaction according to the methods described previously (Table 1).
Primer Sets Used for Real-Time Polymerase Chain Reaction
H, human; HMG-CoA, 3-hydroxy-3-methylglutaryl-coenzyme A; LDL, low-density lipoprotein; R, rat.
Measurement of lipid profiles, CETP activity, and LCAT contents in serum
Rat sera were collected for detecting total cholesterol (TC), total triglycerides, high-density lipoprotein (HDL) cholesterol, LDL cholesterol, alanine aminotransferase (ALT), and aspartate aminotransferase (AST). The serum CETP activity was detected by fluorescent measurement (excitation/emission = 480/511 nm) using a CETP activity assay kit (Abcam), and serum LCAT contents were measured using traditional enzyme-linked immunosorbent assay (ELISA) methods (Abbex, Sugar Land, TX, USA).
Statistical analysis
All data are presented as mean ± standard deviation. The data were statistically evaluated using Duncan's multiple range tests after one-way analysis of variance using SPSS (SPSS PASW Statistic v.23.0; SPSS, Inc., Chicago, IL, USA). Statistically significant differences were accepted at P < .05.
RESULTS AND DISCUSSION
Protection of LRC against liver damage and improvement of lipid profiles
The HC-fed mice had increased serum levels of ALT and AST as well as that of the liver weight compared with those in the NC-fed mice, implicating liver damage. On the contrary, the HC with LRC 4 × 109 and 1 × 1010 CFU/mL presented a significant decrease in the serum levels of ALT and AST compared with that in the HC control group (P < .05) (Table 2). Compared with the NC control group, the HC control group showed elevated levels of total triglycerides, TC, and LDL cholesterol, while displaying a reduction in HDL cholesterol levels. However, the HC with LRC 4 × 109 and 1 × 1010 CFU/mL, and those with decreased total triglycerides, TC, and LDL cholesterol levels were compared with those of the HC control group (P < .05) (Table 2). Previous studies showed that lactic acid bacteria supplementation may have beneficial effects on serum lipid levels in high cholesterol–induced mouse models. 14 –16 These results suggest that certain doses of LRC supplementation protect against liver damage and improve the lipid profile.
Changes in Serum Lipid Profiles of Hypercholesterolemic Rats Fed with Diets Supplemented with Lactobacillus reuteri NCIMB 30242
Values are presented as mean ± SD. Different lowercase letters show a significant difference at P < .05, as determined by Duncan's multiple range test. NC: AIN93G diet; HC (control): HC; PC: HC+simvastatin 15 mg/kg BW; 1 × 109: HC+L. reuteri NCIMB 30242 1 × 109 CFU/rat/day; 4 × 109: HC+L. reuteri NCIMB 30242 4 × 109 CFU/rat/day; 1 × 1010: HC+L. reuteri NCIMB 30242 1 × 1010 CFU/rat/day.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; BW, body weight; CFU, colony-forming units; HC, high-cholesterol diet; HDL, high density lipoprotein; NC, normal control (AIN93G diet); PC, positive control; SD, standard deviation; TC, total cholesterol; TG, triglyceride.
LRC inhibited cholesterol synthesis via attenuation of HMG-CoA reductase and SREBP2
We investigated the advantageous effects of LRC ingestion on hypercholesterolemia to obtain a scientific basis for efficient natural cholesterol-lowering agents. HMG-CoA reductase is the rate-limiting enzyme of the metabolic pathway that produces cholesterol by catalyzing the conversion of HMG-CoA to mevalonic acid and is a target of many drugs, including statins to lower the serum cholesterol levels in hyperlipidemic or hypercholesterolemic patients. 17,18 We determined the HMG-CoA reductase protein expression (Fig. 1A, B) and messenger RNA (mRNA) expression (Fig. 1C) in the liver. As expected, the HC control group exhibited increased mRNA expression (2.37 ± 0.33) and protein expression (1.23 ± 0.13) compared with the NC control group (1.00 ± 0.09 and 0.36 ± 0.07). LRC supplementation significantly reduced mRNA expression (1.99 ± 0.13, 1.42 ± 0.18, 1.31 ± 0.15) and protein expression (0.97 ± 0.11, 0.75 ± 0.08, 0.63 ± 0.05) compared with that in the HC control group.
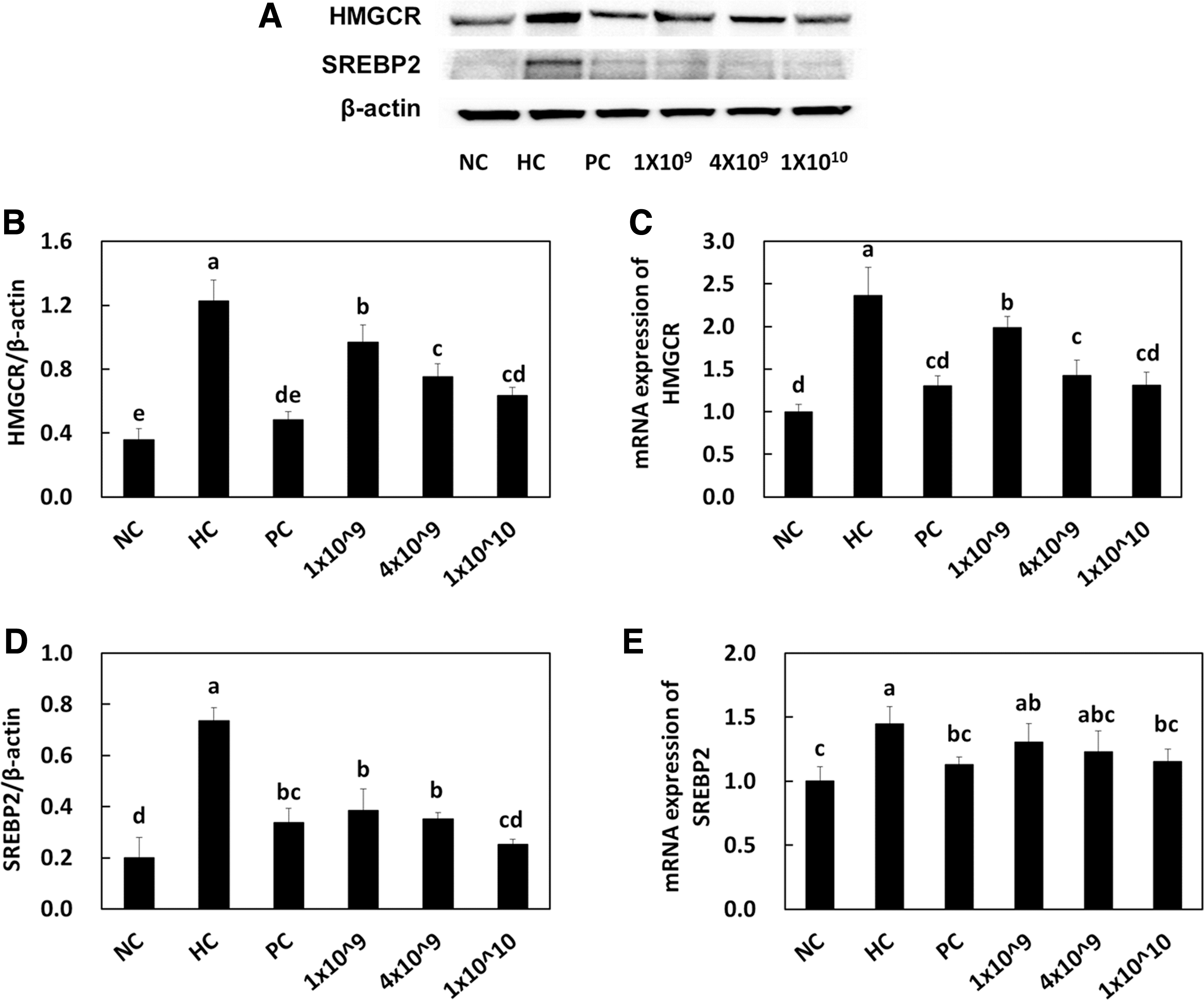
HMG-CoA reductase protein expression
Chen et al. suggested that certain lactic acid bacteria play a role in HMG-CoA reductase gene transcription by regulating the NF-κB signaling pathway. 19 Other previous findings have also indicated that the cellular components of lactic acid bacteria could participate in gene regulation through cell signaling pathways. 20 Based on these results, it can be suggested that certain strains of lactic acid bacteria as well as their secreted metabolites such as organic acids could be used to regulate cellular metabolism.
SREBP2 is a transcription factor that upregulates cholesterol synthesis by activating the transcription of HMG-CoA reductase genes and mevalonate genes. 25 A few recent studies have suggested the involvement of lactic acid bacteria in activating SREBP2 to lower cholesterol levels. 25,26 For instance, Le and Yang announced that Leuconostoc mesenteroides inhibits the expression of SREBP mRNA and related transcription factors in Caco-2 cell lines. 26 Our results suggested that the SREBP2 mRNA expression (Fig. 1E) in hepatic tissues was significantly reduced by the higher number of LRC (1.30 ± 0.15, 1.23 ± 0.16, 1.15 ± 0.10) compared with that of the HC control group (1.45 ± 0.14). Moreover, all doses of LRC (0.39 ± 0.08, 0.35 ± 0.03, 0.25 ± 0.02) significantly inhibited the SREBP2 transcription factor expression in protein levels, in accordance with other studies (Fig. 1D).
LRC increased expressions of LDL receptor and CYP7A1
The LDL receptor is a critical component in the cellular uptake of lipoproteins containing apo-B. ACAT2 plays a critical role in the esterification of intracellular cholesterol storage. 27,28 Yang et al. recently suggested that two lactic acid bacteria, Enterococcus faecium strain and Lactobacillus paracasei, regulated the expressions of LDL receptor, SREBP-1, and CYP7A1 genes to promote cholesterol metabolism and inhibit triglyceride accumulation in hypercholesterolemia rats. 29 Hu et al. identified that lactobacillus strains regulate the mRNA expression of ACAT and CYP7A1 in liver tissues. 27 As expected, we found that the protein and mRNA expressions of LDL receptor and CYP7A1 (Fig. 2) in hepatic tissues are significantly increased by the higher number of LRC compared with that in the HC control group.
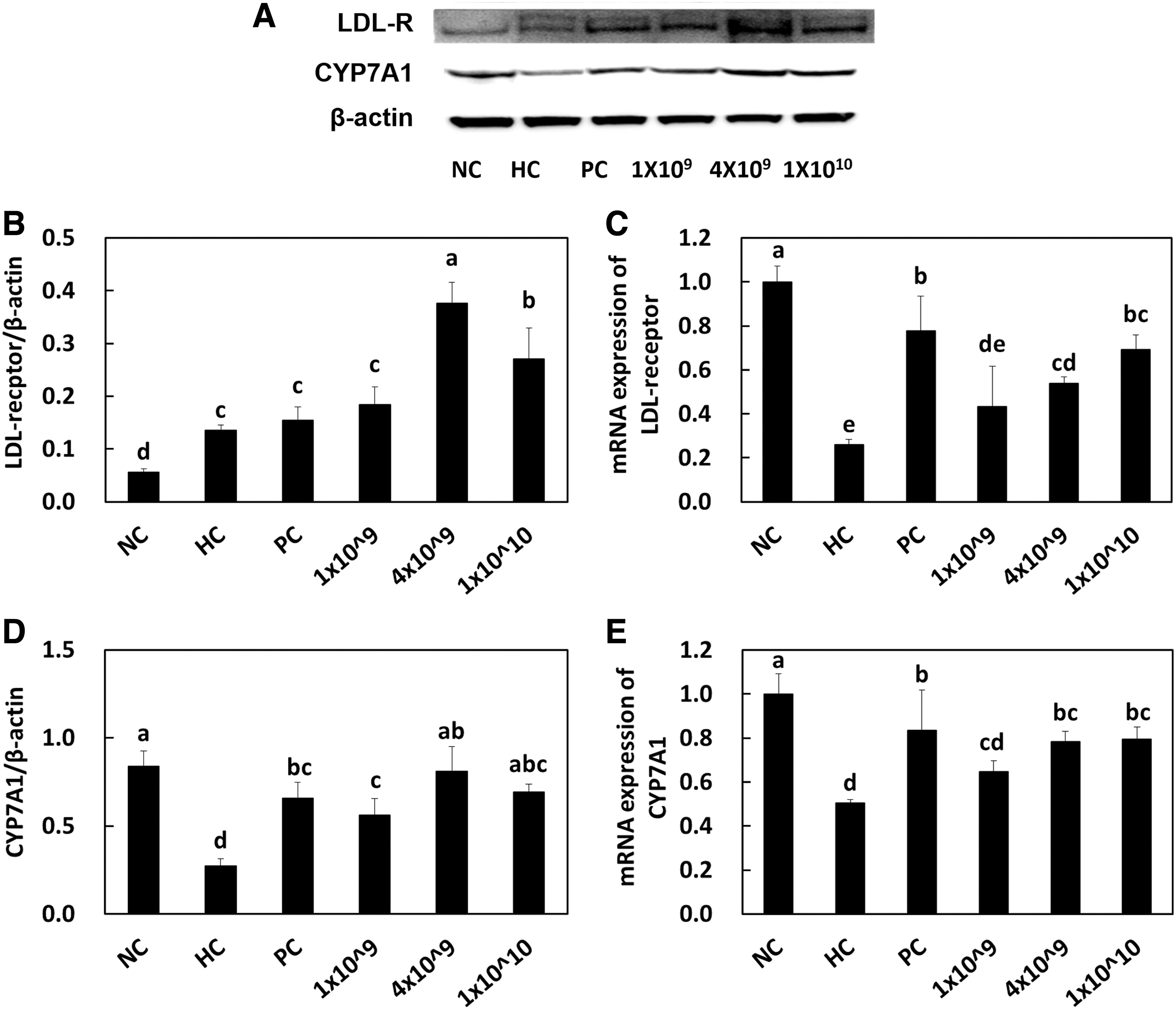
LDL receptor protein expression
Cholesterol is a precursor component of bile acid synthesis that accelerates the metabolism of cholesterol, resulting in the reduction of serum cholesterol levels. CYP7A1 is a rate-limiting enzyme that converts cholesterol to 7-a-hydroxycholesterol, which in turn increases bile acid synthesis and eventually reduces serum cholesterol levels. 30 Sufficient amount of evidence is available that indicates that lactic acid bacteria in involved in the upregulation of CYP7A1 in animal studies, 24,27,31,32 which agrees with our study that states that the LRC treatment increased the CYP7A1 expression at both gene and protein levels in a dose-dependent manner.
However, it is still under debate whether lactic acid bacteria are involved in regulating the CYP7A1 expression. A study has shown that this expression is downregulated in another study, which implicates that the function of CYP7A1 may be activated in a strain-dependent manner. 33 Indeed, abundant evidence is available that states that some strains of lactic acid bacteria can capture intestinal cholesterol as their own extracellular lipopolysaccharides, resulting in a lack of cholesterol absorption and a decrease of CYP7A1 expression. Based on the results discussed so far, it can be suggested that CYP7A1 could be both upregulated and downregulated based on strains and the cellular properties of lactic acid bacteria.
LRC decreased the activity of CETP and increased the content of LCAT in serum
CETP is a serum protein that facilitates the transport of cholesteryl esters generated by LCAT and triglycerides between LDL and HDL. Inhibition of CETP allows improvement of HDL levels in patients with the metabolic syndrome. 34,35 LCAT is also an enzyme that converts serum free cholesterol to cholesteryl ester, which is then sequestered into the core of HDL particles where it further participates in HDL maturation. 36,37 Although not many studies have identified the effects of lactic acid bacteria on the activity or gene expression of CETP and LCAT, we hypothesized that increased serum levels of HDL may be caused by CETP and/or LCAT.
Unfortunately, because of the limited access to detection kits from manufacturers, we could only measure the serum CETP activity and serum LCAT contents. Our data reveal that LRC treatment significantly decreased the serum CETP activity in a dose-dependent manner. However, LRC slightly increased the serum content of LCAT with statistical significance (Fig. 3) (P < .05). These data can indicate that LRC involved in the increased levels of HDL by downregulating the CETP activity rather than upregulating the LCAT expression.
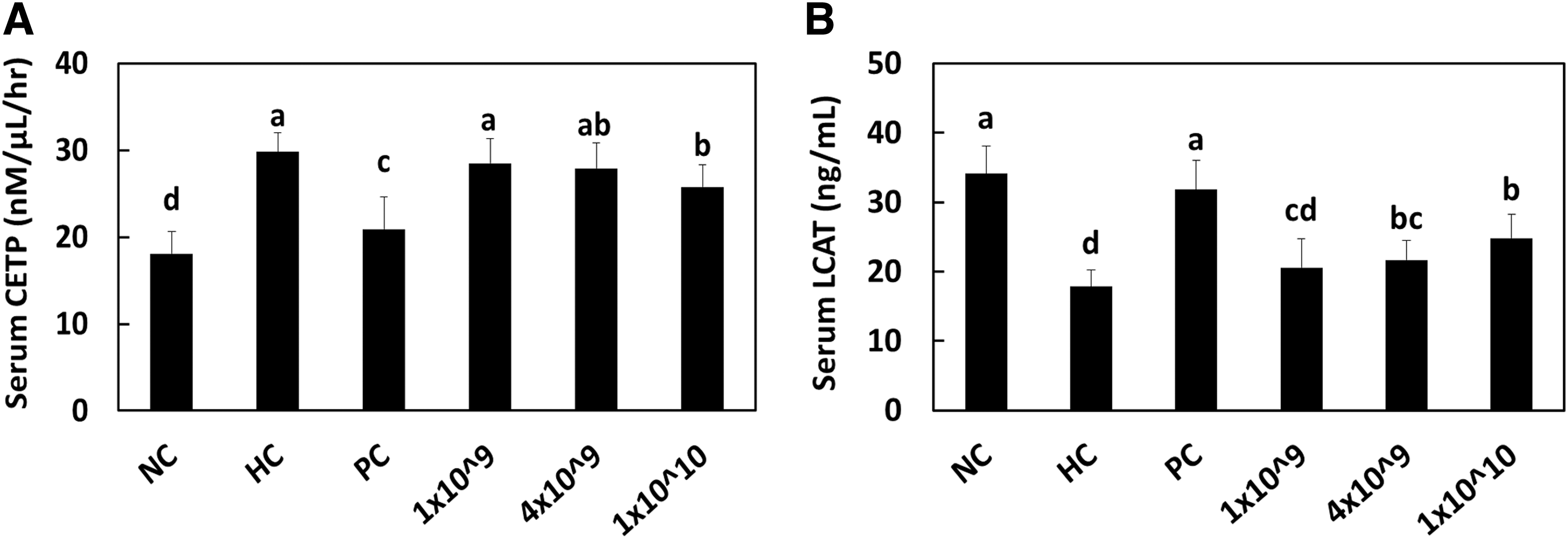
Serum CETP activity
LRC decreased atherosclerotic wall thickness
The HC used herein induced a significant increase in the thickness of the aortic wall than that achieved by the NC. LRC supplementation at higher doses significantly decreased the thickness of the aortic wall compared with that observed in the HC group (P < .05) (Fig. 4). These data indicate that LRC ingestion can restrain the development of atherosclerosis by controlling the aortic wall thickness or elasticity in hypercholesterolemic rats.
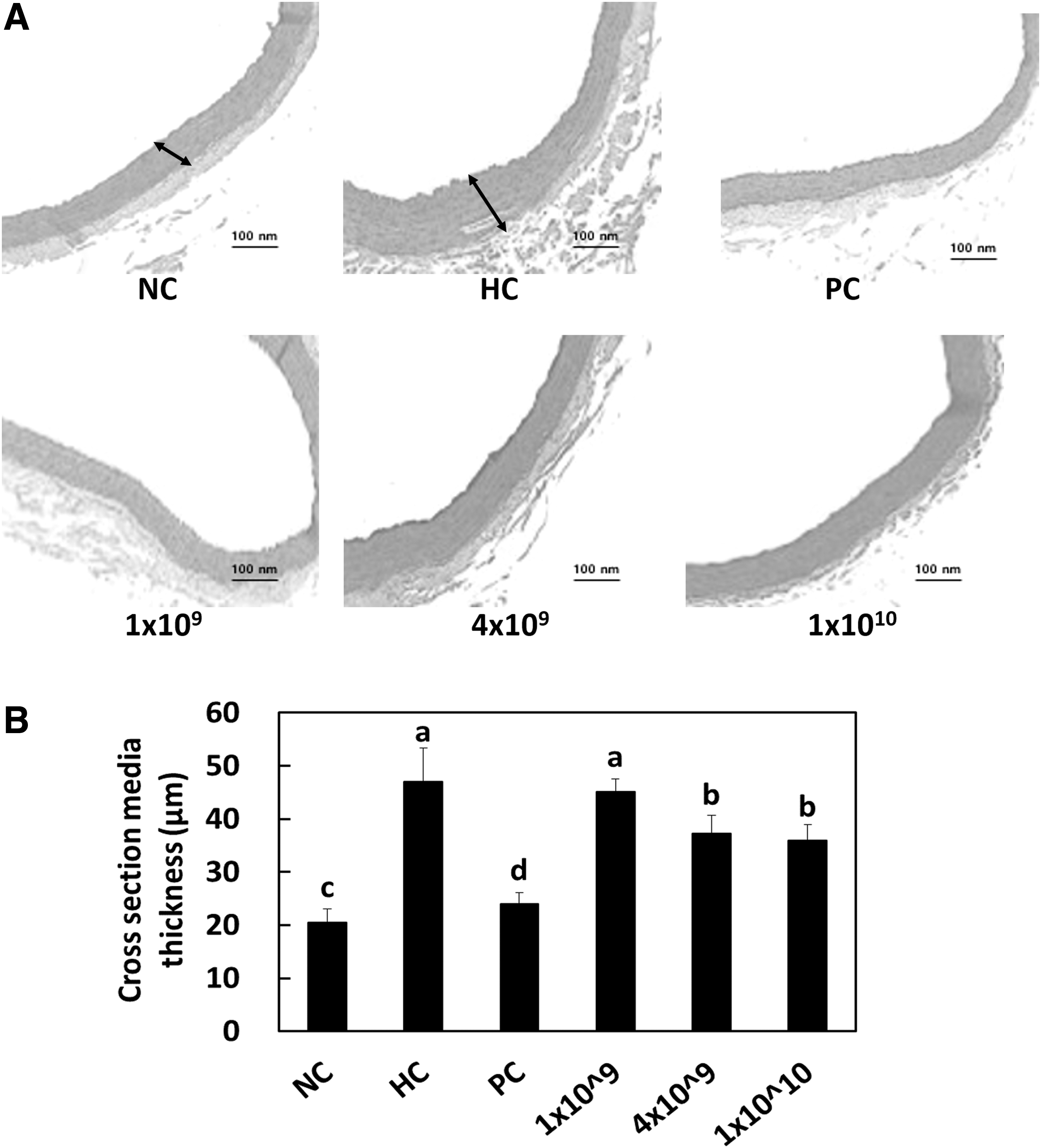
Hematoxylin and eosin staining
LRC stimulated the excretion of cholesterol and bile acid
We measured the TC (Fig. 5A), total triglyceride (Fig. 5B), and bile acid (Fig. 5C) from the feces of rats to establish whether LRC ingestion causes cholesterol excretion. Compared with the control group fed a NC, the group fed a HC showed a significant increase in fecal levels of TC and bile acid. LRC supplementation further increased the TC and bile acid levels significantly in the feces of hypercholesterolemic rats with a statistical significance (P < .05), compared with those in the HC control group (Fig. 5). Increased TC and TG contents in feces suggest that LRC supplementation may be able to reduce the absorption of lipids in the gut, leading to lower circulating levels in the blood.
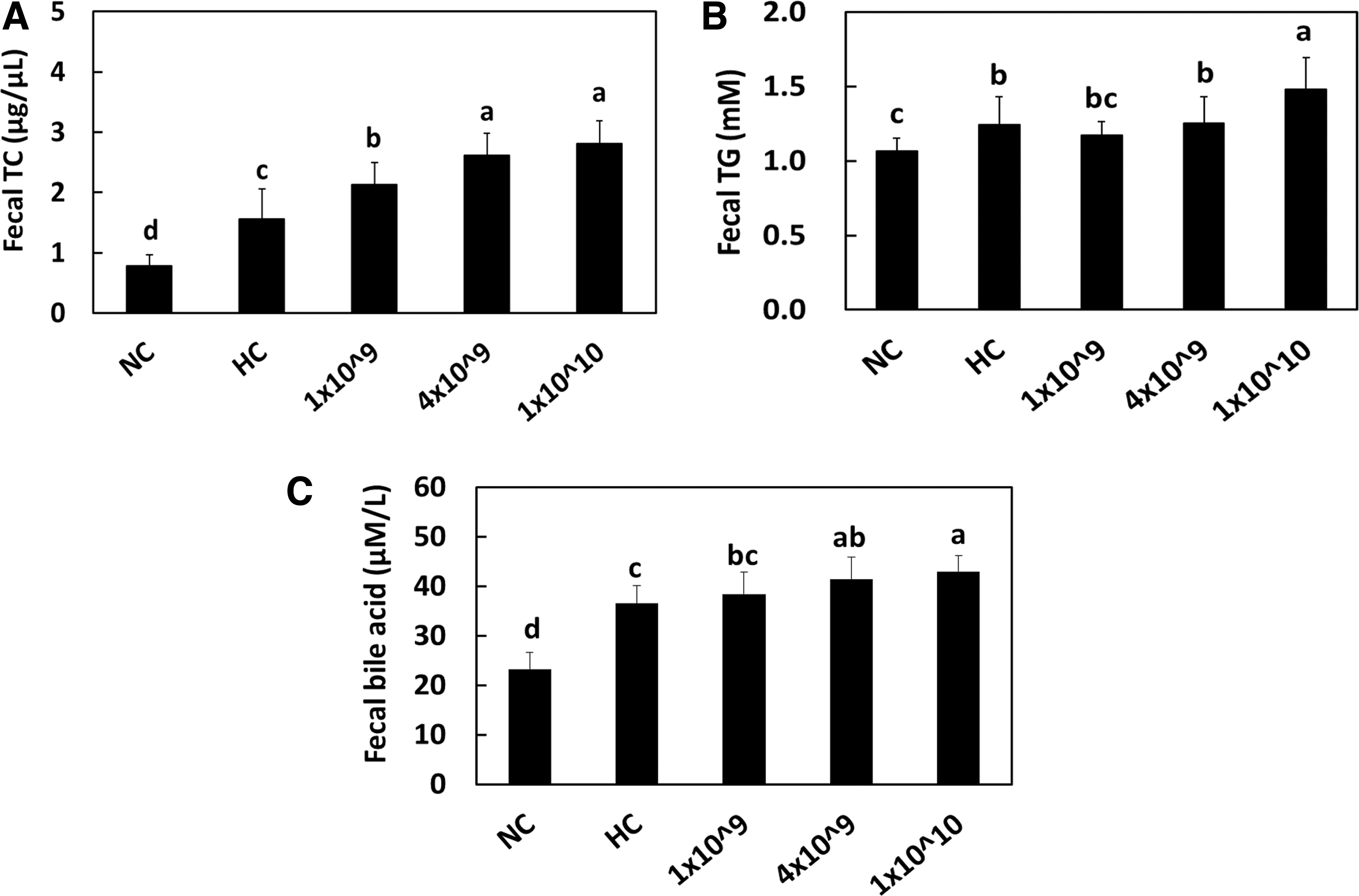
Total cholesterol
These results show that the decreased serum lipid profile levels (Table 2). Also, increased bile acid excretion in feces, suggesting that LRC supplementation may be able to improve bile acid metabolism, which has been associated with various metabolic disorders, such as diabetes, obesity, and nonalcoholic fatty liver disease. These results show that the increased CYP7A1 expression is perhaps involved in bile acid synthesis (Fig. 2D, E). Thus, we suggest that LRC supplementation at higher doses stimulated the excretion of cholesterol and bile acid in hypercholesterolemic rats.
Cholesterol metabolism–related gene and protein expressions in HepG2 cells
To determine the effects of LRC on the mRNA expression of genes related to cholesterol metabolism, including HMG-CoA reductase, LDL receptor, SREBP2, CETP, LCAT, and CYP7A1, we employed four different densities of LRC (5 × 104, 1 × 105, 5 × 105, and 1 × 106 CFU/well) in HepG2 cells. The mRNA expression of HMG-CoA reductase was significantly increased in the control group (2.86 ± 0.24) when compared with the NC group (1.00 ± 0.60). The LRC treatment of HepG2 cells significantly reduced the mRNA expression of HMG-CoA reductase at all doses (1.66 ± 0.21, 1.61 ± 0.12, 1.47 ± 0.21) (P < .05) (Fig. 6A). The mRNA expression of LDL receptor was significantly lower in the control group (0.28 ± 0.33) when compared with the NC group (1.00 ± 0.07).
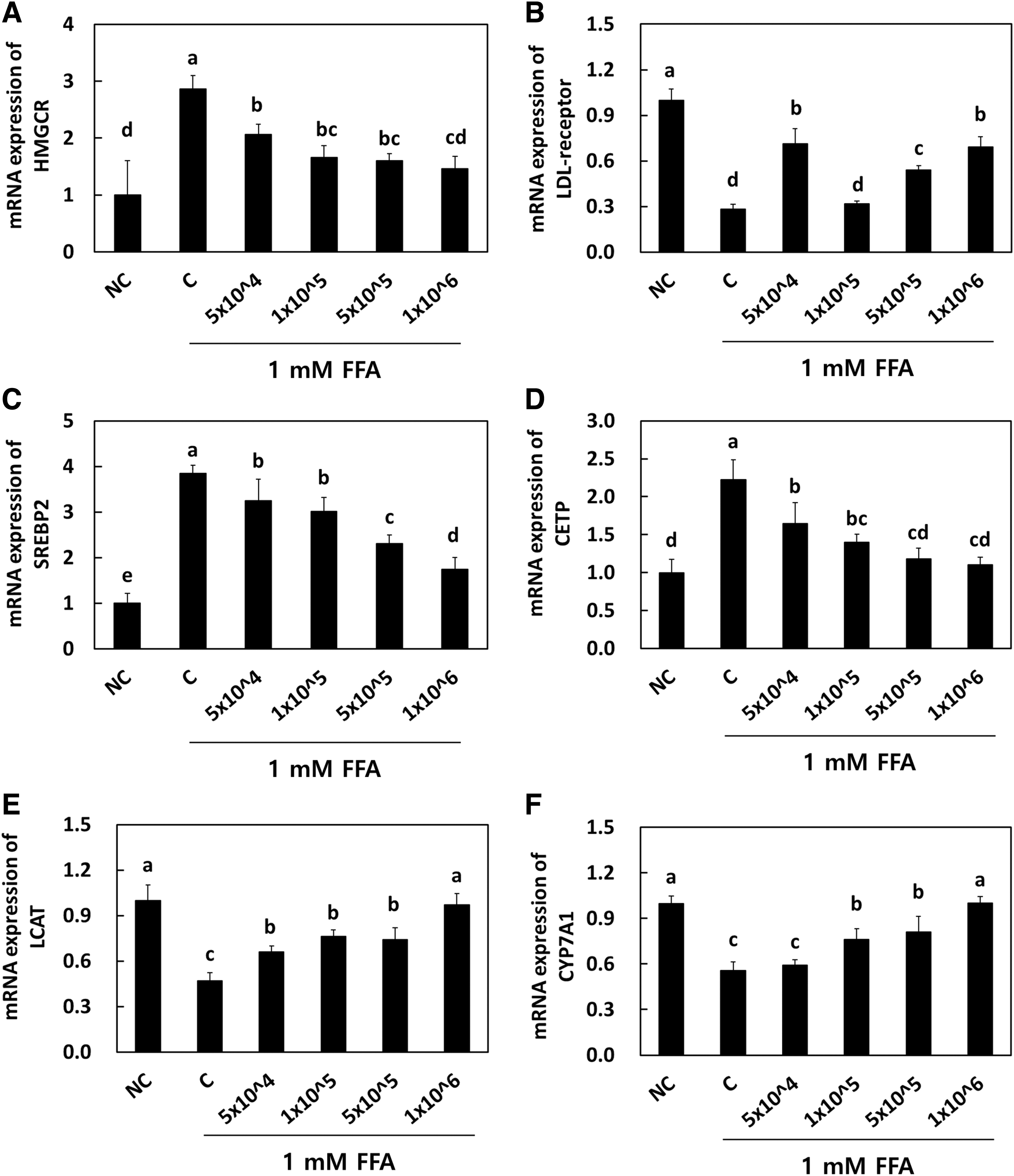
Effects of Lactobacillus reuteri NCIMB 30242 on mRNA expression of HMGCR
The LRC treatment in HepG2 cells significantly increased the mRNA expression of LDL receptor at all doses (0.32 ± 0.02, 0.54 ± 0.03, 0.69 ± 0.07) (P < .05) (Fig. 6B). The mRNA expression of SREBP2 was significantly increased in the control group (3.85 ± 0.17) when compared with the NC group (1.00 ± 0.22). The LRC treatment in HepG2 cells significantly reduced the mRNA expression of SREBP2 at all doses (3.01 ± 0.31, 2.31 ± 0.19, 1.74 ± 0.26) (P < .05) (Fig. 6C). The mRNA expression of CETP was significantly increased in the control group (2.22 ± 0.27) when compared with the NC group (1.00 ± 0.18). The LRC treatment in HepG2 cells significantly reduced the mRNA expression of CETP at all doses (1.40 ± 0.11, 1.18 ± 0.14, 1.11 ± 0.09) (P < .05) (Fig. 6D).
The mRNA expression of LCAT was significantly decreased in the control group (0.47 ± 0.05) when compared with the NC group (1.00 ± 0.10). The LRC treatment in HepG2 cells significantly increased the mRNA expression of LCAT at all doses (0.76 ± 0.04, 0.74 ± 0.08, 0.97 ± 0.07) (P < .05) (Fig. 6E). The mRNA expression of CYP7A1 was significantly decreased in the control group (0.56 ± 0.05) when compared with the NC group (1.00 ± 0.05). The LRC treatment in HepG2 cells significantly increased the mRNA expression of CYP7A1 at all doses (0.76 ± 0.07, 0.81 ± 0.10, 1.00 ± 0.04) (P < .05) (Fig. 6F). We further analyzed the cholesterol metabolism–related protein contents (Fig. 7) in HepG2 cells. Similar to genetic expression, the LRC treatment at higher doses significantly reduced the protein levels of HMG-CoA reductase, SREBP2, and CETP, while that of LDL receptor, LCAT, and CYP7A1 significantly increased it. Hence, our vitro study achieved the same results as those of other animal studies.
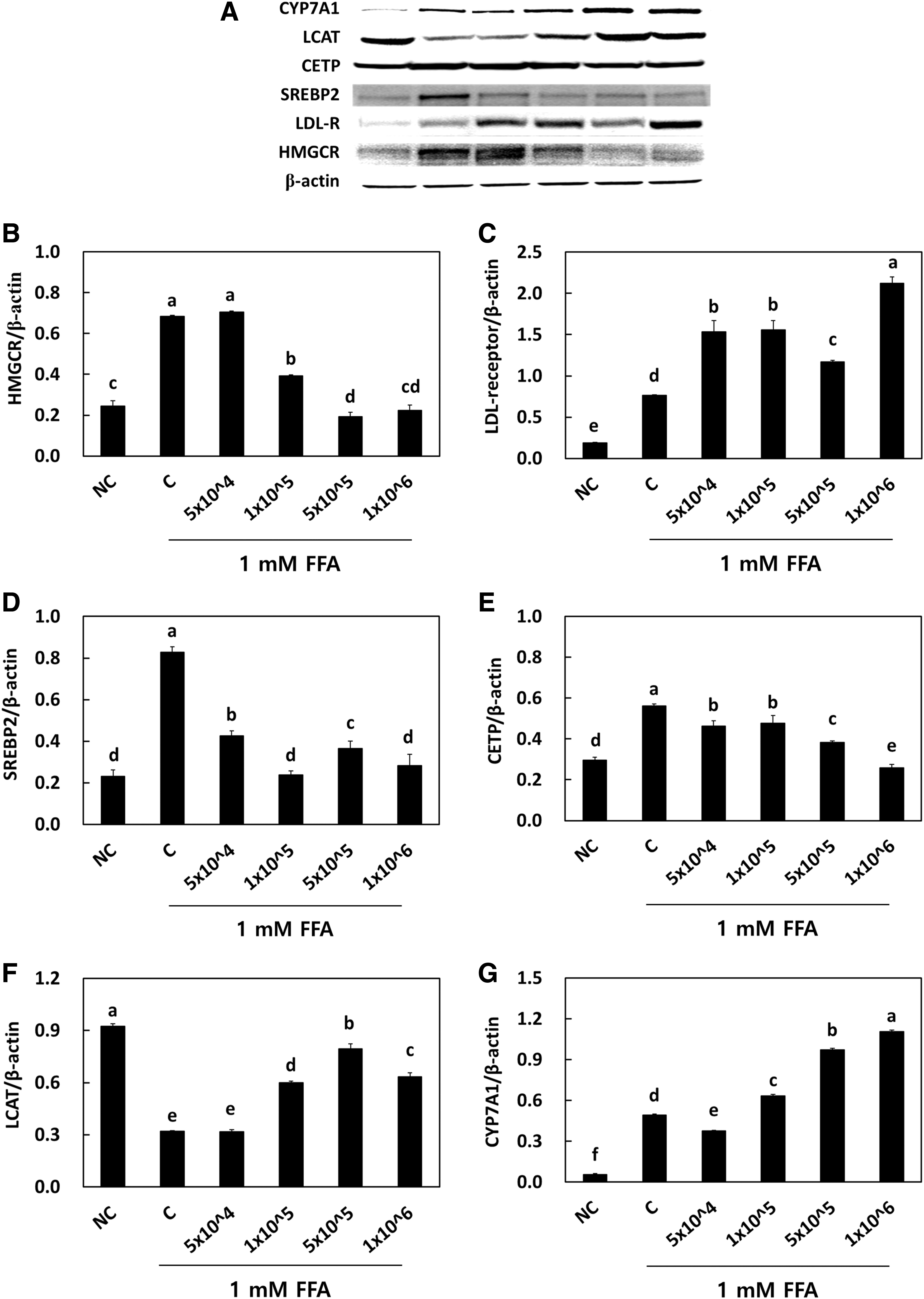
Effects of Lactobacillus reuteri NCIMB 30242 on the protein expression
SUMMARY
The current study evaluated the effects of LRC supplementation on hypercholesterolemia by identifying its effect on cellular cholesterol metabolism and intestinal excretion in hypercholesterolemic rats. We found that LRC, at rather higher doses, suppressed the expression of HMG-CoA reductase, the uptake of LDL cholesterol into tissues, partially inhibited the transport of cholesteryl ester into HDL for maturation, and decreased the intestinal excretion of cholesterol. These results are supported by both mRNA and protein levels in the hepatic tissue and HepG2 cell lines. This study supplied scientific evidence for and explains the mechanisms underlying the antihypercholesterolemic effects of LRC.
Footnotes
AUTHORs' CONTRIBUTIONS
M.L.: Conceptualization, methodology, data curation, visualization, writing—original draft, and writing—review and editing. J.P.: Investigation, formal analysis, and visualization. O.-K.K.: Data curation, writing—original draft, and writing—review and editing. D.K.: Formal analysis, data curation, and visualization. S.K.: Project administration. M.H.: Project administration. J.L.: Conceptualization, writing—original draft, writing—review and editing, project administration, and supervision.
AUTHOR DISCLOSURE STATEMENT
The authors have no conflicts of interest to declare.
FUNDING INFORMATION
No funding was received for this article.