
Abstract
Methotrexate (MTX), a folate antimetabolite, is a cytotoxic drug known to cause cytotoxicity associated with free oxygen radicals. This study investigated the effect of vitamins D3 and K2 on MTX-induced liver cell injury using the zebrafish liver cell line (CRL-2643). Observed effects, levels of antioxidant enzymes, lipid peroxidation marker, and total antioxidant/oxidant status were evaluated by spectrophotometric methods. The mRNA expressions of p53, Bax, and Bcl-2 were measured using RT-PCR. In addition, acridine orange/ethidium bromide staining was performed to analyze the apoptosis status of the cells. The IC50 value of MTX at 48 h was calculated as 442 µg/mL with an MTT assay. The doses of D3&K2 are determined based on the recommended dose to be taken daily. As a result, this study suggests that MTX treatment induced oxidative damage on the liver cell, as assessed by increased reactive oxygen levels, lipid peroxidation, and decreased glutathione levels. In addition, while MTX increased the expression of the p53 and proapoptotic marker Bax, it decreased the expression of the anti-apoptotic factor Bcl-2. In conclusion, D3&K2 treatment protects against MTX-induced liver cell toxicity. It is thought that they can be used as a potential agent in clinical applications with MTX in treatment.
INTRODUCTION
Methotrexate (MTX), a structural analog of folic acid, has been widely used in the treatment of solid tumors and leukemia since the 1940s. 1,2 This pharmaceutical agent is designed to kill rapidly growing cells by disrupting folate metabolism and inhibiting DNA synthesis. 3 Today, it is one of the most common cytotoxic drugs used to treat rheumatoid arthritis, psoriasis, sarcoidosis, inflammatory bowel disease, severe asthma, organ transplantation (as an immunosuppressive agent), and various other autoimmune diseases. 4 However, the expanding clinical use of MTX is associated with significant adverse effects, the most notable being hepatotoxicity. 5,6 Liver damage, particularly following high doses or chronic administration of MTX, has been frequently reported. 7 Drug-induced liver injury accounts for approximately 5% of all jaundice cases, with a spectrum of conditions ranging from acute and chronic hepatitis to bile duct abnormalities and neoplasms. 8 Animal experiments and epidemiological studies have also demonstrated that MTX is carcinogenic, mutagenic, and/or teratogenic. 9
Recent studies by Bhardwaj et al. (2023) have highlighted the reproductive toxicity induced by chemotherapeutic agents, including MTX, and explored innovative therapeutic approaches to mitigate such effects. 10 Additionally, research on apoptosis-inducing compounds, such as the triazolopyrimidine-pyrazole hybrids studied by Kumar et al. (2017), has provided insights into molecular mechanisms that could be leveraged to counteract MTX-induced cellular damage. 11 Furthermore, advancements in metabolomics, as reviewed by Bhardwaj et al. (2024), have shed light on altered cellular pathways in response to chemotherapeutic agents, offering a deeper understanding of MTX’s metabolic impact and potential targets for intervention. 12 By integrating these findings, this introduction underscores the importance of developing targeted strategies to alleviate MTX-induced toxicity while maintaining its therapeutic efficacy.
Cholecalciferol, or vitamin D3, is synthesized in the human body and serves as a hormone precursor. It is converted into 1,25-dihydroxycholecalciferol (1,25-(OH)2 D3), the most biologically active form, through hydroxylation in the liver and kidneys. 13 Vitamin K, a fat-soluble vitamin, acts as a cofactor for γ-glutamyl carboxylase and plays a critical role in blood coagulation and bone formation. Vitamin K supplementation is widely considered essential for managing liver disease. 14 Recently, the combination of vitamin D3 and K2 has gained scientific and public attention due to their complementary effects. Promising research has emerged on the synergistic benefits of combining these two vitamins in dietary supplements. 15 Oxidative stress occurs when there is an imbalance between the production of free radicals and the cell’s antioxidant defenses, leading to lipid peroxidation and oxidative damage. 16 The increase in reactive oxygen species (ROS) and the subsequent decline in antioxidant defenses have been linked to biochemical and physiological alterations, making oxidative stress a key focus of toxicological studies in recent years. 17 –19
The Bcl-2 family of proteins includes both proapoptotic and anti-apoptotic members. These proteins are localized on the mitochondrial membrane and regulate mitochondrial pore permeability. While anti-apoptotic proteins such as Bcl-2 suppress apoptosis, proapoptotic proteins such as Bax play a crucial role in initiating apoptosis. 20 The p53 protein, a well-known tumor suppressor, promotes the synthesis of enzymes involved in DNA repair and regulates the cell cycle. When DNA damage occurs, p53 halts the cell cycle at the G1 phase, preventing progression to the S phase. If the damage is irreparable, p53 triggers the synthesis of Bax, leading to apoptosis. 21
To our knowledge, there are no reports on the protective effects of vitamins D3 and K2 against MTX-induced acute liver toxicity. Therefore, the current study was designed to explore the effects of D3 and K2 on MTX-induced acute oxidative and apoptotic damage in zebrafish liver (ZFL) epithelial cells. Zebrafish are widely used as vertebrate models due to their genetic similarity to mammals and their suitability for large-scale studies.
MATERIALS AND METHODS
Test chemicals
MTX (M8407) was purchased from Sigma Aldrich (Missouri, USA). Cholecalciferol (D3; CS-1179) and Menaquinone (K2; CS-0046153) were purchased as analytical standards from ChemScene LLC (New Jersey, USA).
Cell culture
The ZFL cell line (CRL-2643) was maintained at 28°C in a complete growth medium specified on the ATCC (Virginia, USA) web page. This medium is composed of Leibowitz-15 (50%) (Gibco™, Massachusetts, USA), DMEM High Glucose (35%) (Gibco), and Ham’s F12 (15%) (Gibco), supplemented with 15 mM Hepes (Gibco) and 0.15 g/L NaHCO3 (Sigma Aldrich) and penicillin-streptomycin-amphotericin (1%) (Gibco). For the cell culture, 5% fetal bovine serum (Gibco), 0.01 mg/mL bovine insulin (Gibco), and 50 ng/mL mouse epidermal growth factor (Gibco) were added. 22 Before exposure, the adherent cells were removed from the culture surface with 0.25% trypsin-EDTA (Sigma Aldrich), suspended in a complete medium, and spun at 300 g for 5 min. The resuspended pelleted cells were counted using the Trypan Blue (Sigma Aldrich) exclusion method.
Cell viability and IC50 value were measured using MTT (3-[4,5-dimethylthiazol-2yl]-2,5-diphenyltetrazolium bromide) (Invitrogen™, Massachusetts, USA), method with Thermo Scientific™ Multiskan™ GO Microplate Spectrophotometer (Massachusetts, USA). The principle of this method is to determine the presence of tetrazolium salts in the mitochondria of living cells that form purple formazan crystals as catalyzed by a reductase system. 23
Assessment of oxidative stress and biochemical parameters
The samples were taken into Eppendorf tubes with a 10% (w/v) cooled homogenate buffer and disintegrated in the homogenizer (TissueLyser LT, QIAGEN, Hilden, Germany) with the help of glass beads. Samples were preserved in ice at all stages of the studies. The homogenate was centrifuged at 11.000 g for 20 min at 4°C. The supernatant was carefully decanted and the pelleted part was discarded.
Total antioxidant status (TAS) and total oxidant status (TOS) were determined according to the protocol with commercial kits (Rel Assay Diagnostic, Gaziantep, Türkiye). 24,25 The TOS/TAS ratio is accepted as the oxidative stress index (OSI), which is accepted as a marker of oxidative stress.
NADP-dependent isocitrate dehydrogenase (NADP-IDH) enzyme activity, which is descriptive in liver diseases was performed according to the protocol determined with commercial kits (Rel Assay Diagnostic, Gaziantep, Türkiye).
Malondialdehyde (MDA) level was quantified in the tissue and cell samples using the thiobarbituric acid reaction assay according to the method described by Ledwozyw. 26 Absorbance was measured at 535 nm and the concentration was expressed as nmol MDA/g protein.
The catalase (CAT) enzyme activity was measured following the decrease in absorbance at 240 nm due to hydrogen peroxide (H2O2) consumption. 27 The enzyme activity was expressed as U/mg protein.
The total glutathione (GSH) level was evaluated using the dithionitrobenzoic acid recycling method. 28 GSH concentration was expressed as nmol GSH/g protein.
Acid phosphatase (ACP) and alkaline phosphatase (ALP) enzymes hydrolyze p-nitrophenyl phosphate to p-nitrophenol depending on the pH of the medium (pH = 4.8; pH = 9.8, respectively). The absorbance given by the resulting product at 405 nm is determined by spectrophotometry. 29
Total protein was determined according to the method of Bradford. 30 The intensity of the blue color development was measured at 595 nm against the blank. The total protein concentration was expressed as µg/µL.
mRNA expression analyses
RNA isolation of cell culture samples for genetic analysis was performed with a GeneAll Ribospin RNA isolation kit (Seoul, Korea). The amount and purity of the RNAs obtained were measured with a nanodrop (Thermo-Fisher, Massachusetts, USA) device. The concentration of each isolated RNA sample was standardized at 2 µg/µL.
cDNA synthesis, A.B.T.™ cDNA Synthesis Kit (Atlas Biotechnology, Ankara, Türkiye) was performed in a thermal cycler (Thermo-Fisher) device according to the protocol recommended by the manufacturer. All steps were performed in one cycle and the resulting cDNA was stored at −80°C.
Primer designs were made by identifying specific mRNA sequences and using the NCBI website. The genes and specific primer sequences used in the expression step are given in Table 1. Expression levels of genes were measured on the ABI StepOnePlus (Applied Biosystems, Massachusetts, USA) device using the A.B.T. 2xqPCR CYBR-Green MasterMix (Atlas Biotechnology, Ankara, Türkiye) kit according to the manufacturer’s protocol. In the study, the β-Actin gene was used as a housekeeping gene and Ct values of this gene were used for normalization. Relative quantification of gene expression was performed using the ΔΔCt comparison method.
Primer Sequences Used in RT-PCR Analysis
Acridine orange/ethidium bromide staining
In total, 100 µg/mL acridine orange (AO; Sigma Aldrich), and 100 µg/mL ethidium bromide (EB; Sigma Aldrich) are mixed in PBS (Gibco). A total of 50,000 cells are seeded into each well of a 6-well plate. After 24 h, the substances are administered. At the end of the exposure period, the medium in the wells is discarded and 200 µL of AO/EB staining is applied to it. It is then analyzed under a fluorescent microscope (Zeiss Axio Vert A.1, Oberkochen, Germany) at 480–505–535 nm. Tests were done in triplicate, counting a minimum of 100 total cells each. 31
Statistical analysis
Statistical analysis was done using the SPSS software program (IBM Corp., Armonk, NY, USA). Data were expressed as mean ± standard error of the mean. Comparisons between the two groups were done by parametric Student’s t-test and nonparametric Mann–Whitney U test while assuming unequal variances. A one-way analysis of variance (ANOVA) was made between more than two groups. Tukey’s post hoc test was performed to compare the significant differences between groups following ANOVA. P < .05 was considered significant.
RESULTS
ZFL epithelial cells were cultured in a standard medium for 24 h prior to MTX treatment. Following this, the cells were exposed to MTX at concentrations of 10, 25, 50, 100, 250, 500, and 1000 µg/mL and incubated for 48 h. Control cells were treated with a culture medium only. The cytotoxic effects of MTX were assessed using the MTT assay. Cell viability, compared with the control, was determined to be 96%, 90%, 75%, 69%, 65%, 48%, and 45% at MTX concentrations of 10, 25, 50, 100, 250, 500, and 1000 µg/mL, respectively (Fig. 1). These results demonstrate that MTX exerts a dose-dependent cytotoxic effect on ZFL epithelial cells. Statistical analysis revealed that the IC50 value of MTX at 48 h was 442 µg/mL. The doses of vitamins D3 and K2 were selected based on daily recommended intake levels, 15 with final concentrations of 50 ng/mL for D3 and 40 ng/mL for K2.
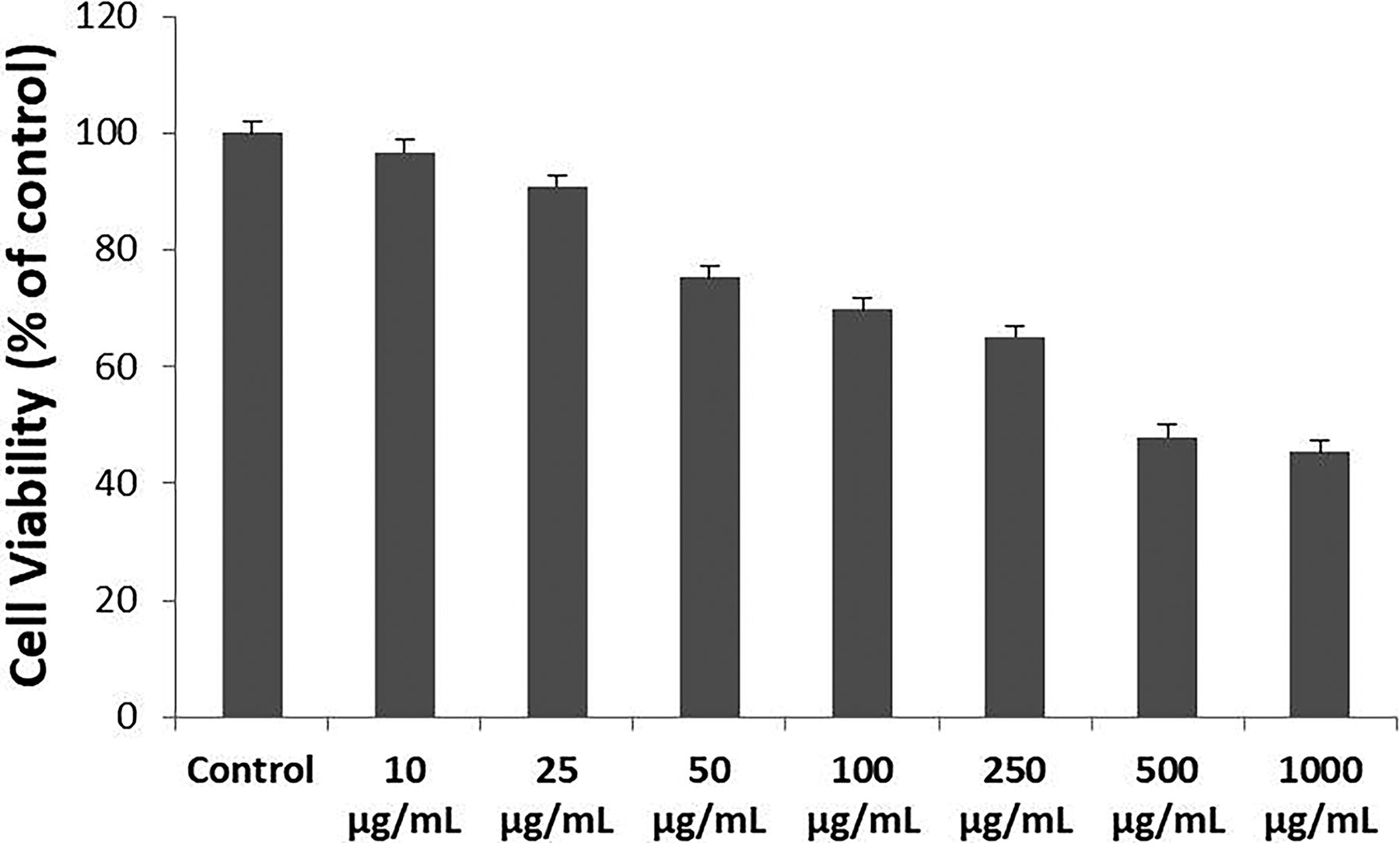
Effects of MTX on the viability of ZFL epithelial cells. Cells were treated with different concentrations of MTX for 48 h, and cell viability was assessed using the MTT assay. All experiments were independently performed three times. Data are expressed as mean ± standard error (SE). MTX, methotrexate; ZFL, zebrafish liver.
After evaluating the IC50 value of MTX, ZFL epithelial cells were exposed to MTX, D3&K2, and combined MTX, D3&K2 for 48 h and analyzed oxidative stress status. TOS of ZFL cells was increased in the MTX group (P = .03), was decreased in the D3&K2 group (P = .01), and statistically did not change in the MTX+D3&K2 group (P = .37) when compared with the control (Fig. 2A). The TAS of cells in the MTX group was significantly reduced (P = .004), while in the D3&K2 group was enhanced (P = .02). TAS in the MTX+D3&K2 group was not statistically significant (P = .3) when compared with the control (Fig. 2B). When the OSI is calculated according to these results, it is suggested that vitamins D3&K2 can balance MTX-induced oxidative stress (Fig. 2C).
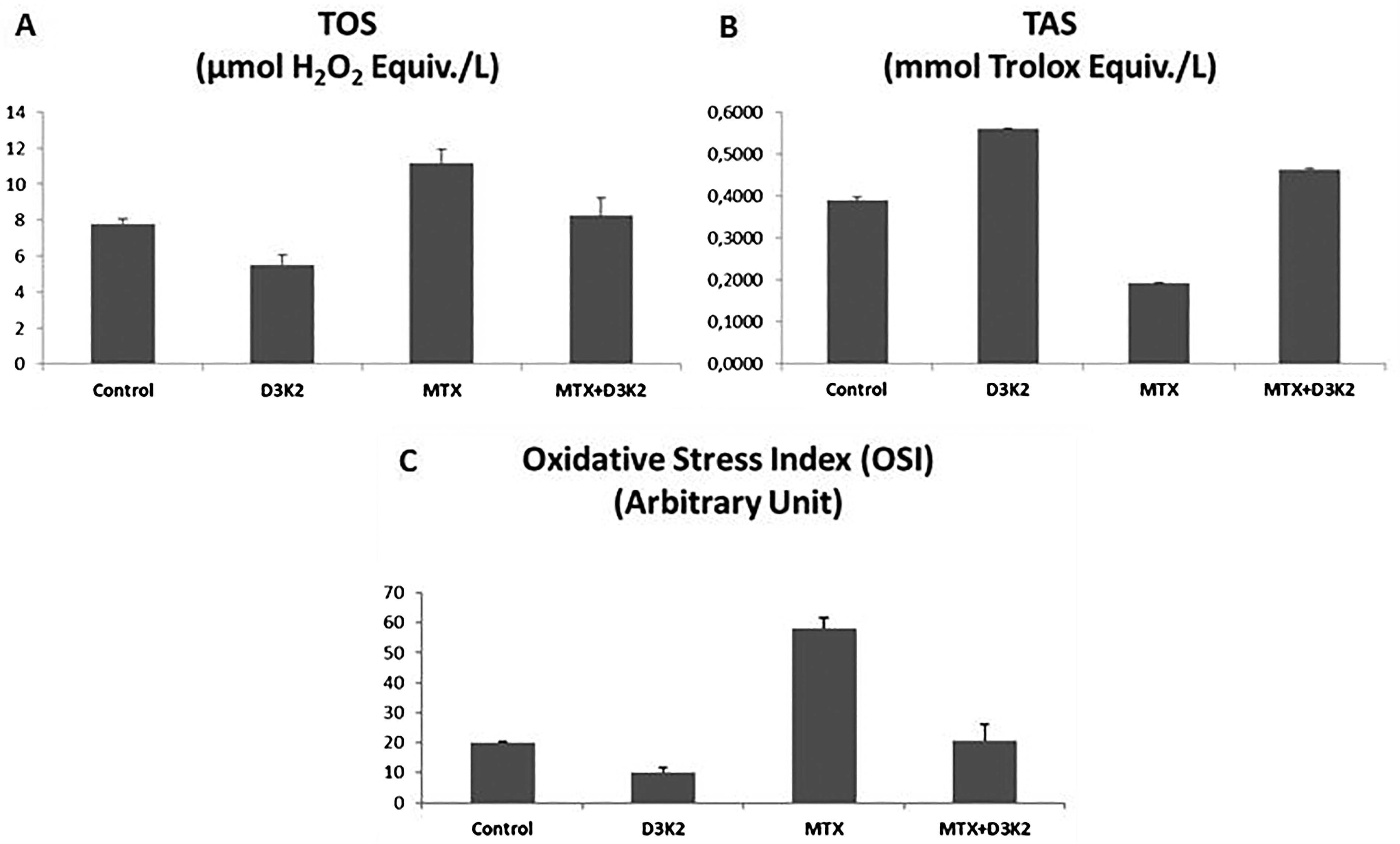
Effects of vitamins D3&K2 on MTX-induced oxidative stress in ZFL epithelial cells.
After exposure, NADP isocitrate dehydrogenase (NADP-IDH) enzyme activity decreased significantly in the MTX group (P = .01) but remained unchanged in the D3&K2 and MTX+D3&K2 groups compared with the control (Fig. 3A). CAT enzyme activity was significantly reduced in the MTX group (P = .02) but increased in the D3&K2 (P = .04) and MTX+D3&K2 (P = .009) groups (Fig. 3B). It was observed that the ACP enzyme activity was significantly reduced in the MTX group, which was administered 48 h IC50 dose (442 µg/mL), compared with the control (P = .002). But it was not statistically significant when compared with the control in D3&K2 (P = .06) and MTX+D3&K2 (P = .05) groups (Fig. 3C). ALP enzyme activity enhanced in MTX (P = .04), D3&K2 (P = .01), and MTX+D3&K2 (P = .03) groups was compared with the control (Fig. 3D).
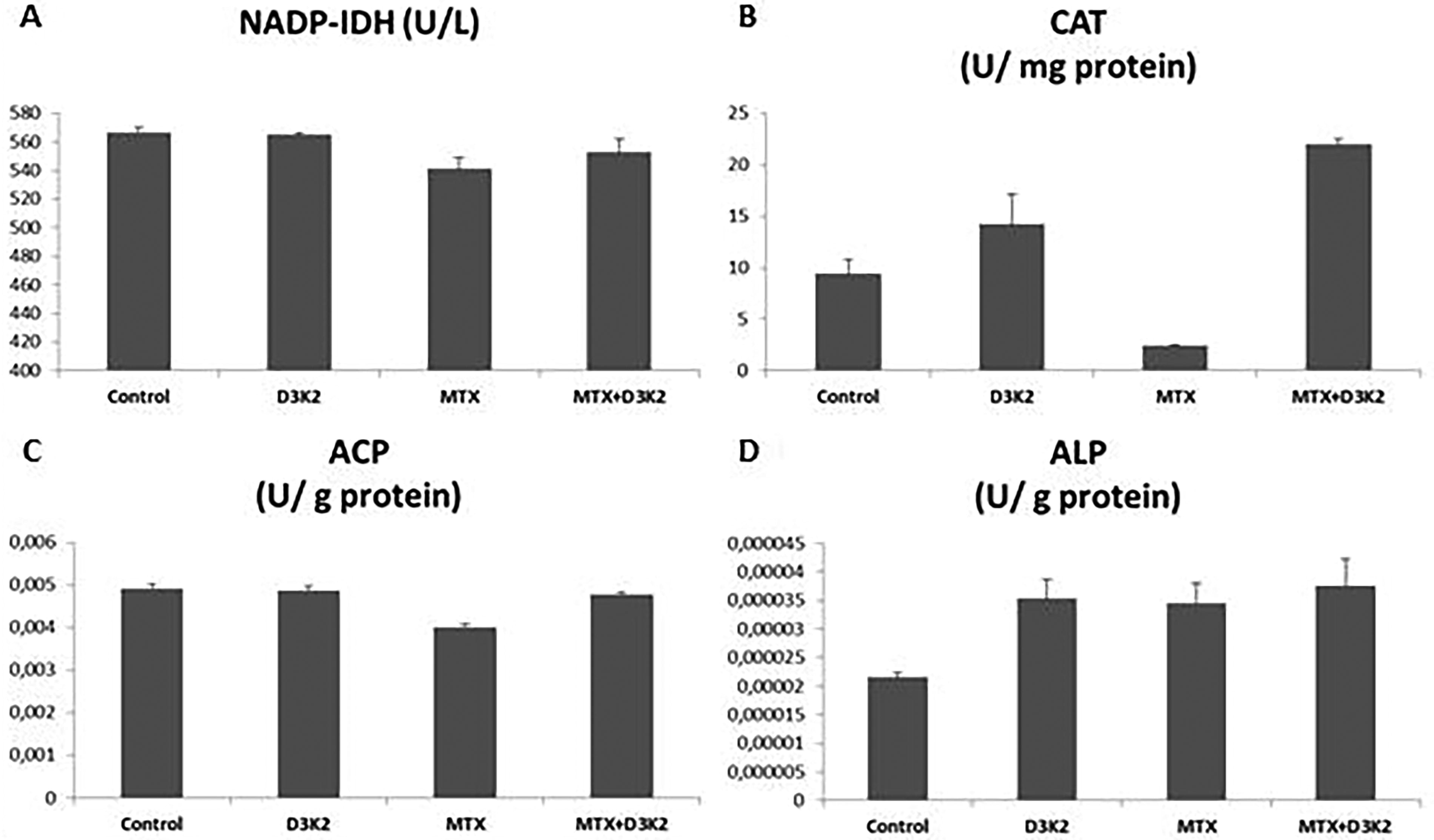
Effects of MTX, D3&K2 on enzyme activities in ZFL epithelial cells.
Effects of MTX on ZFL epithelial cells caused lipid peroxidation. MDA is one of the important metabolites of lipid peroxidation. The amount of MDA increased in the MTX group, and was administered 48 h IC50 dose (442 µg/mL), compared with the control (P = .03) was observed. But showed no significant changes in the D3&K2 (P = .2) or MTX+D3&K2 (P = .3) groups compared with the control (Fig. 4A). The total content of GSH was considerably reduced in the MTX group (P = .009), while it was insignificantly changed in D3&K2 (P = .09) and MTX+D3&K2 (P = .06) groups compared with the control (Fig. 4B).
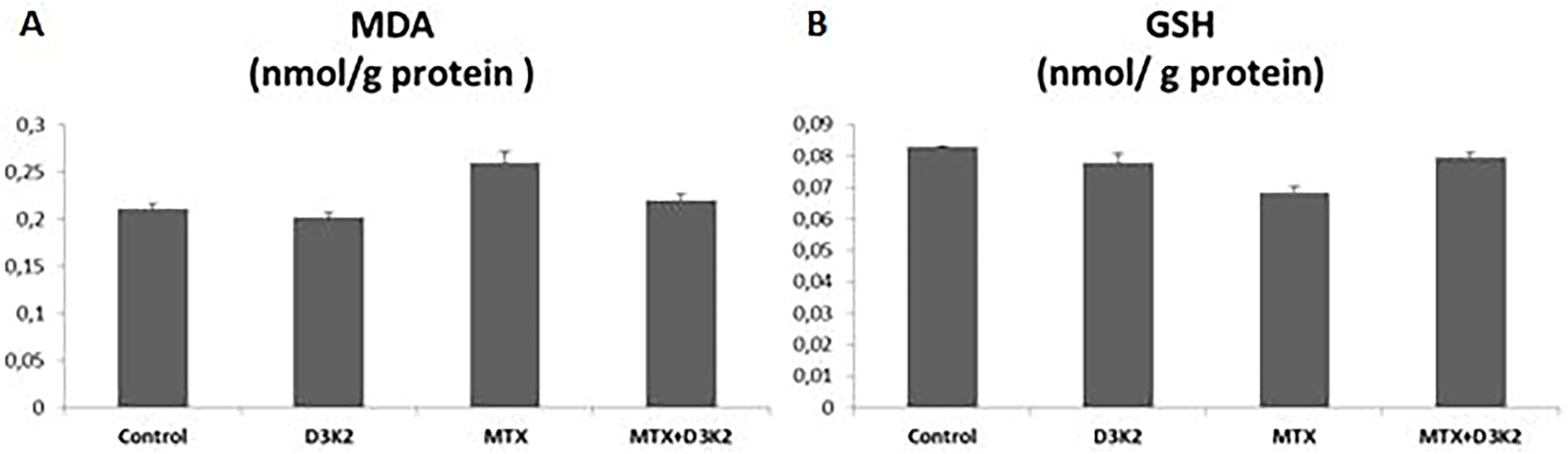
The oxidative effects of MTX on ZFL epithelial cells.
The expression levels of apoptosis-related genes (Bax, Bcl-2, and p53) were analyzed in ZFL cells treated with MTX and D3&K2. Compared with the control, treatment with the IC50 dose of MTX (442 µg/mL) for 48 h resulted in a 3.63-fold increase in Bax expression, a 1.69-fold increase in p53 expression, and a 5.2-fold decrease in Bcl-2 expression. In the D3 (50 ng/mL) and K2 (40 ng/mL) group, Bax and p53 expression levels decreased by 1.38-fold and 1.23-fold, respectively, while Bcl-2 expression increased by 1.19-fold compared with the control. When equated the MTX and MTX+D3&K2 groups, Bax and p53 gene expression decreased, while Bcl-2 gene expression increased, indicating a protective effect of the vitamins (Fig. 5).
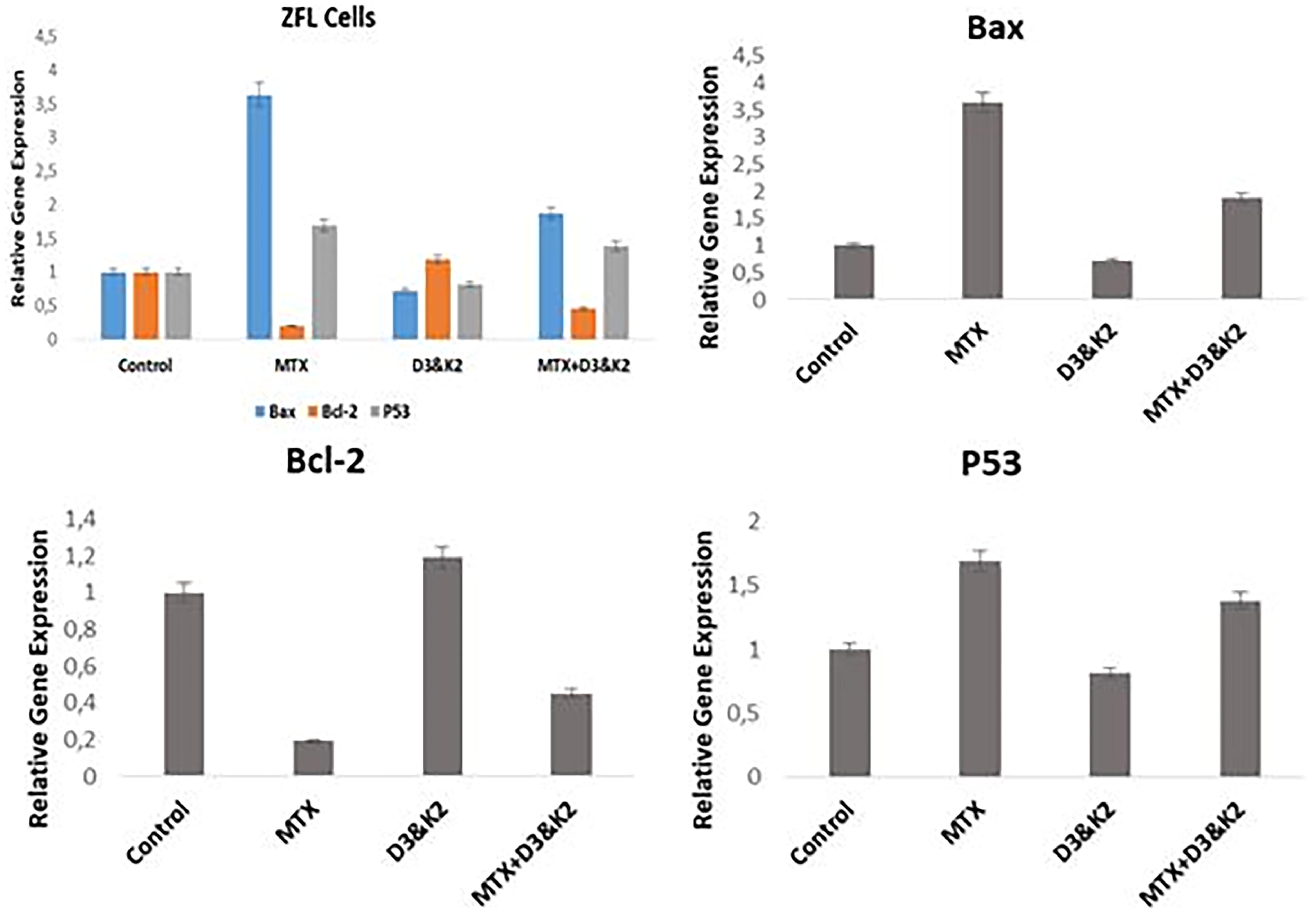
Results of the Bax, Bcl-2, and p53 gene expressions in ZFL epithelial cells after exposure to 48 h IC50 doses of MTX (442 µg/mL), D3 (50 ng/mL) and K2 (40 ng/mL), and MTX+D3&K2. All experiments were independently performed three times. Data are expressed as mean ± standard error (SE).
ZFL cells were stained with AO/EB 48 h after treatment with the IC50 dose of MTX (442 µg/mL), D3&K2, or MTX+D3&K2. Fluorescent microscopy revealed apoptotic cells, characterized by granular yellow-green AO nuclear staining, and necrotic cells, marked by uneven orange-red fluorescence at their periphery (Fig. 6). Fragmented cells were also observed. Quantitative analysis showed an increase in apoptotic and necrotic cells in the MTX group, while the addition of D3&K2 reduced the number of apoptotic cells in the MTX+D3&K2 group (Table 2).
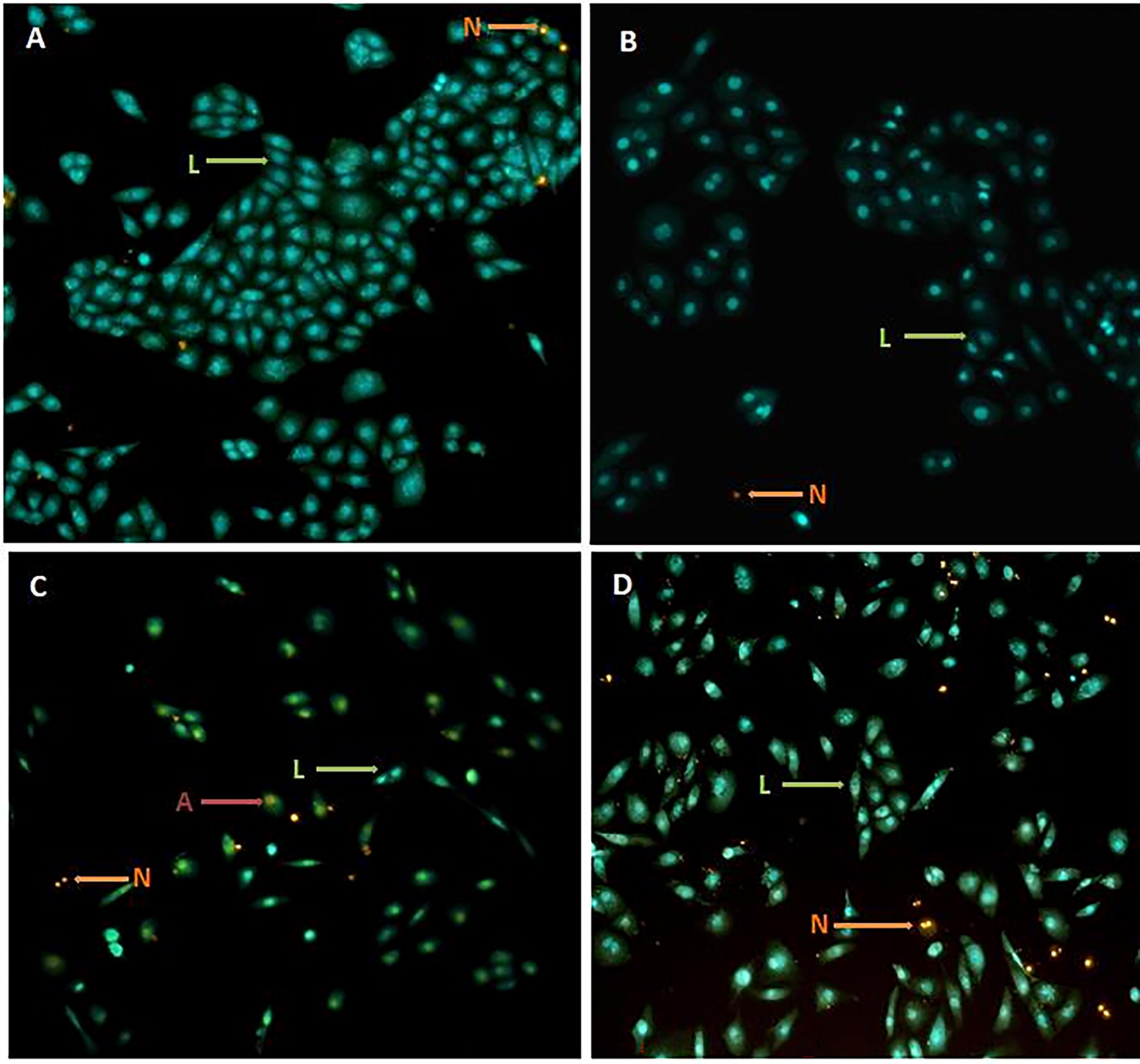
Results of acridine orange/ethidium bromide (AO/EB) staining in ZFL epithelial cells after exposure to MTX and D3&K2.
The Percentage of Live, Apoptotic, and Necrotic Cells After Exposure to 48 h IC50 Doses of MTX (442 µg/mL), D3 (50 ng/mL) and K2 (40 ng/mL), and MTX+D3&K2
Data are expressed as mean ± standard error (SE).
MTX, methotrexate.
DISCUSSION
MTX is a drug applied clinically in a wide variety of diseases, including systemic lupus erythematosus, rheumatoid arthritis, psoriasis, and neoplastic diseases. 32,33 Despite its known therapeutic effects, MTX has some serious side effects and causes damage to certain organs such as the liver, kidney, and bone marrow. 34 Hepatotoxicity is the most important complication during MTX therapy. 6 Although the mechanism of the hepatotoxic effect of MTX is not fully known yet, it is thought that intracellular MTX polyglutamate accumulation and folate reduction may be responsible. 35 Folic acid is often added to the treatment regimen to avoid the side effects of MTX. However, side effects are also seen in excessive folic acid use. 36 In this study, we investigated the effect of vitamins D3 and K2 due to their complementary effects on MTX-induced hepatotoxicity in zebrafish liver cells.
The use of the ZFL cell line (CRL-2643) as a model for liver cell injury offers a reliable platform for studying MTX-induced toxicity and evaluating potential protective agents. However, future studies should consider extending these findings to in vivo models and human clinical trials to validate the therapeutic potential of vitamins D3 and K2 in MTX-treated patients. Additionally, exploring the synergistic effects of these vitamins with other antioxidants or hepatoprotective agents, such as N-acetyl-l-cysteine (NAC), could further enhance their efficacy.
MTX is indirectly involved in liver mitochondria damage via depletion of mitochondria enzymatic and non-enzymatic antioxidants. 37 MTX administration may also cause the accumulation of ROS, targeting macromolecules and many pathological processes. 38 The mechanisms underlying MTX-induced hepatotoxicity are not fully understood, but it has been suggested that both free radicals and inflammatory cytokines cause hepatotoxicity. 39,40 Previous studies have reported that excessive production of free radicals, increased lipid peroxidation, and depletion of enzymatic or non-enzymatic antioxidant defense systems play a role in MTX-induced liver damage. 40,41 It was understood that the development of MTX-induced toxicity was related to free oxygen radicals and hydrogen peroxide, and antioxidant agents such as melatonin, nicotine amide, methionine, vitamin E, and NAC were tried to protect the tissues from the damage caused by MTX. 42,43 Likewise, many experimental studies have been conducted showing the protective effects of antioxidant substances such as L-carnitine, nicotinamide, methionine, melatonin, folic acid, 42,44,45 and ursodeoxycholic acid. 7 Şen et al. (2014) examined the oxidative lung damage induced by MTX in rats, it was reported that the levels of TOS and OSI increased, both in tissue and serum, in subjects given a single dose of 20 mg/kg MTX. In the same study, it was observed that carvacrol and pomegranate extract, given as antioxidants, increased TAS levels and decreased OSI. 46 It is a proportional index obtained by dividing total oxidants by total antioxidants. A high OSI indicates an increase in oxidative stress. In this study, it was observed that MTX application increased the levels of TOS and OSI in ZFL cells. However, these results changed with the D3 and K2 applications and reached the levels of control.
The IC50 value of 442 µg/mL for MTX at 48 h provides a quantitative basis for understanding the drug’s cytotoxic effects on liver cells. This value, combined with the observed biochemical and molecular changes, reinforces the need for protective strategies to minimize MTX-associated hepatotoxicity. The doses of vitamins D3 and K2 used in this study were based on daily recommended intake levels, making them clinically feasible for further investigation in human trials. In addition, the study by Bhardwaj et al. (2024) on cadmium-induced toxicity in the male reproductive system highlights the broader applicability of antioxidant interventions in mitigating chemical-induced cellular damage. 47 Their findings on the role of oxidative stress and apoptosis in toxicity mechanisms further validate the importance of exploring protective agents like vitamins D3 and K2 in MTX-induced hepatotoxicity.
The first line of defense against the cell-damaging effects of oxidative stress on the liver is the antioxidant defense system that neutralizes free radicals’ deleterious effects. 48 Therefore, the level of ROS in liver tissue may be estimated by the analysis of the activity of the antioxidant enzymes. Using an in vitro model, the current study investigated the effect of vitamins D3 and K2 in MTX-induced hepatotoxicity. The observed effects were assessed by measuring the levels of antioxidant enzymes, lipid peroxidation markers, as well as biochemical evaluations.
Free radicals can affect many molecules such as proteins, carbohydrates, enzymes, and DNA in the organism. However, considering all biological molecules, lipids are perhaps the most sensitive and most affected by free radicals. Lipid peroxidation and post-peroxidation products damage the membrane structure and various cell components; membrane permeability and microviscosity are significantly affected. 49 Malondialdehyde is a metabolite of lipid peroxidation and is used as an indicator of oxidative stress in cells. 40,50 Aslaner et al. (2015) found that medical ozone reversed the hepatotoxicity induced by MTX in liver tissues with its antioxidant effect; showed that it prevents lipid peroxidation and neutrophil infiltration and increases the amount of glutathione. 51 Glutathione is an important cytosolic radical scavenger. 50 Studies report that MTX causes a decrease in liver GSH levels. 40,52,53 Similar to previous studies, in this study, it was determined that MTX decreased the total GSH level and increased the MDA level. It has been observed that D3 and K2 has curative effects on biochemical and oxidative stress markers. Akbulut et al. (2014) showed that amifostine, ascorbic acid, and NAC can prevent MTX-induced hepatotoxicity through oxidative stress. 54 Safaei et al. (2018) showed that gallic acid is effective against MTX-induced hepatotoxicity and that catalase, superoxide dismutase, and glutathione peroxidase activities, which increased with MTX, decreased with gallic acid administration. 55 In this study, NADP-IDH, CAT, and ACP enzyme activities decreased with MTX in ZFL cells.
The protective effects of vitamins D3 and K2 against MTX-induced liver cell injury represent a significant finding of this study. The restoration of antioxidant enzyme levels, reduction in lipid peroxidation, and normalization of apoptotic marker expression suggest that these vitamins mitigate oxidative stress and prevent apoptosis. This is in line with emerging evidence on the antioxidant and anti-inflammatory properties of vitamins D3 and K2. 56,57 Furthermore, the role of antioxidants in mitigating toxicity is supported by studies such as Bhardwaj and Saraf (2020), who demonstrated that NAC effectively regulated DNA fragmentation and apoptosis in granulosa cells exposed to methoxychlor toxicity. 58 This parallels the protective mechanisms observed in our study, where vitamins D3 and K2 similarly counteract oxidative damage and apoptotic events. Apoptosis is a form of cell suicide that plays an important role in the development and maintenance of tissue homeostasis in multicellular organisms. This study investigated the role of several apoptosis regulators (p53, Bax, and Bcl-2) in MTX-induced hepatotoxicity. The tumor suppressor gene p53, involved in cell cycle arrest after DNA damage, was significantly up-regulated after exposure to MTX but decreased in the MTX+D3&K2 group. Bcl-2 and Bax produce mitochondrial-associated proteins with antagonistic effects, exhibiting anti-apoptotic and pro-apoptotic activity, respectively. Bcl-2 was slightly down-regulated compared with p53 and the expression level of Bax was up-regulated (Fig. 5). These results indicated that p53-induced apoptosis was enhanced by MTX, mediated by the p53-regulated gene (eg Bax). Studies have reported that p53 mRNA levels can be greatly induced under different stimuli. 59 MTX is known and shown to increase p53 expression to induce cell apoptosis in mammalian cells. 60 –62 In the present study, it was observed an increased Bax expression and a decreased Bcl-2 expression after MTX exposure. These results could indicate a possible increase in the apoptosis pathway.
Oxidative stress contributes to the development of proinflammatory in liver cells, and ultimately to apoptosis of cells. 63,64 Vitamins D and K are particularly involved in calcium metabolism, but both have apoptosis-inhibiting properties. Studies are showing that vitamin D reduces proapoptotic proteins, but increases anti-apoptotic proteins. 65,66 Huang et al. (2021) indicated that vitamin K2 also could activate Bad via PI3K/Akt-signaling and inhibit the caspase-3 mediated apoptosis. 67 The findings of this study demonstrate that MTX induces significant oxidative stress and apoptosis in liver cells, as evidenced by increased ROS levels, lipid peroxidation, and altered expression of apoptotic markers. These results align with previous studies highlighting MTX’s role in generating free radicals and disrupting cellular redox balance, leading to hepatotoxicity. 68,69 The observed increase in p53 and Bax expression, coupled with a decrease in Bcl-2 levels, underscores the activation of the intrinsic apoptotic pathway in response to MTX-induced oxidative damage. This mechanistic insight is consistent with the well-documented role of p53 in mediating cell cycle arrest and apoptosis under conditions of cellular stress. 70
CONCLUSIONS
This study demonstrates that MTX treatment induces oxidative tissue damage in liver cells, as evidenced by elevated ROS levels, increased lipid peroxidation, and reduced glutathione (GSH) levels. Importantly, the findings reveal that combined treatment with vitamins D3 and K2 provides significant protection against MTX-induced liver cell toxicity. These results highlight the potential of vitamins D3 and K2 to exert both protective and curative effects on oxidative hepatic damage caused by MTX.
The implications of this research are substantial, as vitamins D3 and K2 may serve as promising adjuvants to mitigate the adverse effects of chemotherapy or other MTX-based treatments. Their ability to counteract oxidative stress and cellular damage suggests that they could be integrated into clinical applications to enhance the safety and efficacy of MTX therapy. This is particularly relevant for patients undergoing long-term or high-dose MTX treatment, where hepatotoxicity remains a major concern. Furthermore, this study underscores the suitability of the zebrafish cell model as a reliable and effective tool for studying liver toxicity. The model’s genetic and physiological similarities to mammals, combined with its experimental advantages, make it a valuable alternative or complement to traditional in vivo models.
To advance these findings, future studies are recommended to elucidate the precise molecular mechanisms underlying the hepatoprotective effects of vitamins D3 and K2. Additionally, further research should explore the optimal dosing, timing, and long-term safety of these vitamins in combination with MTX, both in preclinical models and clinical trials. Such investigations will be critical for translating these promising results into practical therapeutic strategies, ultimately improving patient outcomes in chemotherapy and MTX-based treatments.
Footnotes
AUTHOR’S CONTRIBUTIONS
The author designed the study, performed the experiments, analyzed the results, and wrote the article.
ETHICS APPROVAL
This article does not contain any studies with human participants or animals performed by any of the authors.
DATA AVAILABILITY STATEMENT
No data associated with the article.
AUTHOR DISCLOSURE STATEMENT
The author has no relevant financial or non-financial interests to disclose. The authors confirm that there are no conflicts of interest.
FUNDING INFORMATION
The author declares that no funds, grants, or other support were received during the preparation of this article.