
Abstract
Previous studies have shown that continuous daily consumption of polyphenolic-rich blueberries (BB) reduced neuroinflammation and improved age-related declines in motor and cognitive function in rodents and humans, but the optimal intake of BBs is currently unknown. To evaluate the beneficial effects of BBs on neuroinflammation and motor and cognitive function relative to the frequency of consumption, the diets of aged F344 rats (18 months; n = 15/group) were supplemented with either a continuous control diet, a continuous 2% wild BB diet, or an intermittent (3 days on, 4 days off) 2% wild BB diet for 2 months prior to behavioral testing. Because neuroinflammation has been identified as a major contributing factor to the behavioral declines seen in aging and neurodegenerative diseases, serum was collected pre-diet and at the end of the study to assess whether diet-induced changes in serum can alter the production of inflammatory stress signals in activated HAPI rat microglial cells in vitro. Serum from rats continuously and intermittently fed BBs had beneficial effects by reducing the expression of inflammatory enzymes cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS) in microglia, but only continuous feeding attenuated nitrite production. Intermittent BB-fed rats performed better on the rotarod and committed fewer errors in the radial arm water maze on Day 2 compared to control-fed animals. Based on these findings, BBs may not have to be consumed daily to elicit beneficial effects, possibly due to the retention of polyphenols and their metabolites in tissue and circulation.
INTRODUCTION
The U.S. population is aging and the number of Americans ages 65 and older is projected to increase 50% in the next 30 years. With advancing age, both animals and humans show declines in motor and cognitive function. 1 As the number of older adults in the population continues to increase, the incidence of age-associated deficits also increases, even in the absence of neurodegenerative disorders. These deficits include changes in fine motor control, gait, and balance, 2 as well as reduced cognitive processing speed, decreased executive function, and impairments in memory and spatial learning. 3 Therefore, improving resilience to behavioral decline has great promise and impact, creating a need to identify modifiable factors that regulate the proper maintenance of brain function and facilitate healthy aging. 4
While the etiology of age-related functional changes is multifactorial, long-term increases in and susceptibility to, the effects of oxidative stress and inflammatory insults are thought to contribute the behavioral deficits. 5 Plant foods contain an assortment of bioactive phytochemicals, which have antioxidant and anti-inflammatory activities. 1 Therefore, diets rich in fruits and vegetables may reduce the risk of age-related motor and cognitive impairment due to the presence of these antioxidant/anti-inflammatory polyphenols. 6,7 Once consumed, polyphenols are absorbed and extensively transformed into bioactive metabolites that circulate throughout the bloodstream and accumulate in various tissues. 8 Studies in aged rats demonstrate that berry polyphenols are bio- and neuro-available, and one study reported a strong relationship between improved performance in a maze task and a total number of anthocyanin compounds found in the cortex of aged rats fed with a 2% blueberry-supplemented diet. 9 Possible mechanisms for the beneficial effects of polyphenols include the ability to scavenge free radicals, reduce inflammation, and modulate neuroplasticity, neurogenesis, autophagy, cellular signaling pathways, and gene expression. 6,7,10
Previous studies have shown that continuous daily consumption of blueberry (BB) improved age-related declines in motor and cognitive function in old animals and animal models of accelerated aging and neuroinflammation. 7,11,12 Studies in our lab showed that BBs were able to reverse several parameters of brain aging, as well as age-related motor and cognitive deficits, when superimposed on an already well-balanced, healthy rodent diet (“chow”) and then fed to rats ad libitum from 19–21 months of age. 13,14 However, the optimal intake of BBs is currently unknown. As most people do not consume BBs every day, it is of interest to determine if there are beneficial effects of intermittent consumption, and if they differ from those of continuous consumption. A study by Elks and colleagues 15 showed that consuming BBs for two days induced oxidative stress in the kidney, brain, and liver of rats; however, continuous feeding ultimately led to reduced oxidation. 15 In this and other studies, hormesis has been proposed as a mechanism by which phytochemicals could improve health indirectly by stimulating cellular repair mechanisms. 13,15 –17 Therefore, it is possible that intermittent consumption of BBs could: (1) have increased beneficial effects relative to continuous consumption, due to the repeated stimulation of cellular repair mechanisms, i.e., hormesis; (2) have a reduced beneficial effect, as the total dose is lower; or (3) have the same effect as continuous consumption, if metabolites are retained in tissue.
The purpose of the present study was to evaluate the beneficial effects of wild blueberries on motor and cognitive function relative to the frequency of blueberry consumption. The diets of aged F344 rats (18 months; n = 15/group) were supplemented with either a continuous control diet (modified NIH-31 diet), a continuous 2% wild blueberry diet, or an intermittent 2% wild blueberry diet for 2 months prior to behavioral testing. Rats in the intermittent 2% blueberry diet group consumed the 2% blueberry diet for 3 days, followed by a 4-day control diet (washout period) each week for the duration of the study. Serum was collected pre-diet and at the end of the study to assess whether diet-induced changes in serum can reduce the production of inflammatory stress signals in stressed HAPI rat microglial cells in vitro.
MATERIALS AND METHODS
Diets
The pelleted diets were prepared at Envigo (now Inotiv, Inc., Madison, WI) by adding freeze-dried wild blueberry powder (provided by the Wild Blueberry Association of North America, WBANA, Old Town, ME) to the control diet (20 g/kg diet, 2% w/w). The control diet was a modification of the NIH-31 diet, with the amount of corn adjusted to compensate for the added volume of the blueberry powder. The control NIH-31 diet was the same as that used in earlier studies where fruits, including blueberries, were found to be beneficial in mitigating brain aging when fed for 2 months at 2% of the diet. 14,18 –20
Animals
45 18-month-old male Fischer 344 rats, obtained from the NIA colony (Charles River, Frederick, MD), were individually housed in stainless steel mesh suspended cages, provided food and water ad libitum, and maintained on a 12-hour light/dark cycle. Following a 3-week acclimatization period to the facility, which included handling and tail bleed (see below), the rats were weight-matched and assigned to three groups fed with either a continuous control diet (CON), a continuous 2% wild blueberry diet (CBB), or an intermittent 2% wild blueberry diet (INT) (n = 15/group) for a total of 8 weeks until euthanized. Rats in the intermittent diet group consumed the 2% wild blueberry diet for 3 days (Wednesday morning through Saturday morning), followed by a 4-day control diet (washout period, Saturday morning through Wednesday morning) each week for the duration of the study. Behavioral testing was conducted in weeks 7 and 8. After 8 weeks on their respective diets, the now 21-month-old animals were euthanized by decapitation, and trunk blood and brain tissue were collected for future research.
Body weights were recorded once a week throughout the study. There were no differences in average weight between the groups during the study (P > .05, CON 392.5 ± 10.0; CBB 400.2 ± 11.0; INT 391.5 ± 8.8). Food intakes were assessed 2x/week (on the days when the food was switched, Wednesday and Saturday) for the first 5 weeks of experimental feeding. There were no differences in average food intake between the groups during the study (P > .05, CON 17.70 ± 0.24; CBB 18.05 ± 0.35; INT 18.11 ± 0.30). All rats were observed daily for clinical signs of disease. During the course of the study, 6 rats died or were euthanized due to excessive weight loss and were excluded from final analyses: 1 in the CON, 1 in the CBB, and 4 in the INT group, leaving 14 rats in the CON and CBB groups, and 11 rats in the INT group for final analyses. These numbers are in line with mortality rates for this strain and age of rats. 21 Animals were utilized in compliance with all applicable laws and regulations as well as the principles expressed in the National Institutes of Health, USPHS, and the Guide for the Care and Use of Laboratory Animals. This study was approved by the Tufts Medical Center Animal Care and Use Committee.
Behavioral tests
Motor function assessment
During week 7, rats performed a battery of age- and diet-sensitive tests of psychomotor behavior, 22 –24 to measure strength, endurance, and balance. Each test was performed once, separated by a break between tasks. Rats were tested in random order blocked by the treatment group. Briefly, the tests included the following: (1) rod walking, which measures psychomotor coordination and the integrity of the vestibular system by placing the rats on a stationary, horizontal rod and recording latency to fall; (2) wire suspension, which measures muscle strength and the prehensile reflex by allowing the rats to grasp a horizontal wire suspended above a thick foam-core pad with its forepaws and recording latency to fall; (3) plank walking, which measures balance and coordination by exposing the rats to horizontal planks of three different widths (13 mm, 25 mm, and 38 mm; counterbalanced) and recording latency to fall; (4) inclined screen, which measures muscle tone, strength, stamina and balance by placing the animal on a wire mesh screen that is tilted 60° to the horizontal plane of the floor and recording latency to fall; and (5) accelerating rotarod, which measures fine motor coordination, balance and resistance to fatigue by having rats walk on a rotarod (Ugo Basile, Italy), consisting of a slowly accelerating (+2 rpm/30 s) rotating dowel (7 cm diameter), and recording latency to fall. For a more detailed description of the tests, see Shukitt-Hale et al. 24
Cognitive assessment
During week 8, all rats were tested in the 8-arm radial water maze (RAWM), which is an age- and diet-sensitive test of spatial learning and memory. 20,25 The maze is a black, circular pool of water (maintained at 23°C) fitted with internal walls that form 8 channels (‘arms’) radiating from a central zone. The maze is surrounded by numerous extra maze cues in a dimly lit room, and the rat uses the cues to effectively navigate the maze. Once placed in the pool, rats must swim to a submerged black platform (2 cm below the water’s surface), hidden in one of the maze arms, to escape.
The RAWM test was conducted for 5 trials per day, for 5 consecutive days, with a new platform location each day. Trials 1–4 were consecutive acquisition trials and Trial 5 was a 30-minute delayed retention trial. Rats were tested in quasi-random order, with the restriction that one rat from each group be tested in succession. For each trial, rats were given 90 sec to find the platform and then were gently guided to it by the researcher if they failed to find it on their own. Rats were allowed to remain on the platform for 15 sec once found, after which time rats were immediately placed in a novel start location (trials 2–4) to again locate the platform. The sequence of start locations changed daily without repetition. After the 4th trial, rats were towel-dried and then returned to their home cage for 30 min, after which they were run on the 5th trial, to test for retention. Trial 5 was analyzed because it is a good assessment of short-term memory and memory consolidation. Latency and speed were recorded using image tracking software (HVS Image, Hampton, England), while arm entries were recorded and scored manually. Errors were assessed by measuring the number of times a rat entered any maze arm that did not contain the platform. For a more detailed description of the maze, see Shukitt-Hale et al. 20,25
Serum collection
Blood samples were collected from either tail bleed (pre-diet) or trunk blood (post-diet) at euthanasia. Tail bleeds were performed as previously described, 20 with ∼1 mL of blood being withdrawn from the lateral tail vein. Trunk blood was collected in a 50 mL Falcon tube immediately after the rats were decapitated, which took place on a Monday, ∼48 h following the switch to control diet in the intermittent group. To obtain serum, blood was allowed to coagulate for one hour at room temperature, and the serum was separated from the clot, centrifuged at 10,000 g for 2 min, and stored at −80°C until analysis.
Cell culture
Microglia are the primary immune cells of the central nervous system (CNS) and regulate the inflammatory response in the CNS. 26 Highly aggressively proliferating immortalized (HAPI) rat microglial cells (generously provided by Dr. Grace Sun, University of Missouri, Columbia, MO) were maintained in Dulbecco’s modified Eagle’s medium (DMEM, Invitrogen, Grand Island, NY) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 ug/mL streptomycin at 37°C in a humidified incubator under 5% CO2. Cells were grown in 100 mm plates and then split into 12-well plates prior to treatment. All treatments were assayed in duplicate. For experiments, cells were incubated in serum-free DMEM, without phenol red, supplemented with serum collected from the rats pre- and post-diet at a concentration of 10% for 8 h. The media was then removed followed by stimulation with lipopolysaccharide (LPS, Sigma-Aldrich, St. Louis, MO), a potent endotoxin and activator of microglia 26 at 100 ng/mL overnight.
Nitrite quantification
To assess the production of the reactive nitrogen species nitric oxide (NO) from LPS-stressed microglia, extracellular release of nitrite (NO2 −) was measured in cell-conditioned supernatant by Greiss reagent (Invitrogen, Carlsbad, CA) according to manufacturer’s instructions and as described in Cahoon and colleagues. 27 Absorbance was read at 548 nm and the concentration of nitrite was calculated with the linear equation derived from the standard curve generated by known concentrations of nitrite.
Western blots
Following the serum treatment and LPS stimulation, cells were washed in ice-cold PBS, re-suspended, and lysed by agitation in CelLytic-M Cell Lysis Reagent (Sigma) containing phenylmethylsulfonylfluoride (PMSF, 10 ug/mL), and centrifuged at 18,000 g for 10 min at 4°C to yield the resultant supernatant lysate. Western blots were performed as described previously. 27,28 Primary antibodies for inflammatory enzymes inducible nitric oxide synthase (iNOS, Millipore, Billerica, MA) and cyclooxygenase-2 (COX-2, Santa Cruz, Dallas, TX) were used at 1:1000 dilutions for incubation overnight at 4°C. Following ECL (enhanced chemiluminescence) development using a detection kit (Bio-Rad, Hercules, CA), the optical density of antibody-specific bands was analyzed by the VisionWorks LS image acquisition and analysis software (UVP, Upland, CA).
Statistical analyses
For each serum measure, the data were analyzed by a two-way (diet group and time of collection) analysis of variance (ANOVA) to test for statistical significance at a level of P < .05 using Systat (SPSS, Inc., Chicago, IL). Post-hoc comparisons were performed using Fisher’s Least Significant Difference (LSD) test to determine differences among groups. For the motor tests and the RAWM, data were analyzed by Dunnett’s test using the animals on the control diet as the reference group.
RESULTS
Behavioral tests
For the rotarod test, even though the diet group effect showed only a trend toward significance [F(2,36) = 2.26, P = .12], post-hoc testing showed that rats in the INT group walked on the rotarod longer than CON-fed rats (P < .05), demonstrating improved motor performance, while performance for the CBB-fed group was not different from the CON group (Fig. 1). There were no differences between the diet groups on any of the other motor tests.

Latency to fall (mean ± SEM) in the rotarod task in rats fed a control (CON), continuous (CBB), or intermittent (INT) blueberry diet. Asterisk (*) indicates a significant difference from the control group (P < .05) as measured by Dunnett’s test.
In the RAWM, there were no differences between the diet groups on the retention trials (Trial 5) in latency to find the hidden platform overall or by day (Fig. 2A). However, the diet group effect for total number of errors showed a trend toward significance on Day 2 [F(2,33) = 2.68, P = .08], with post-hoc testing showing that rats in the INT group made fewer working memory errors (arm reentries) than the CON group (P < .05, Fig. 2B).
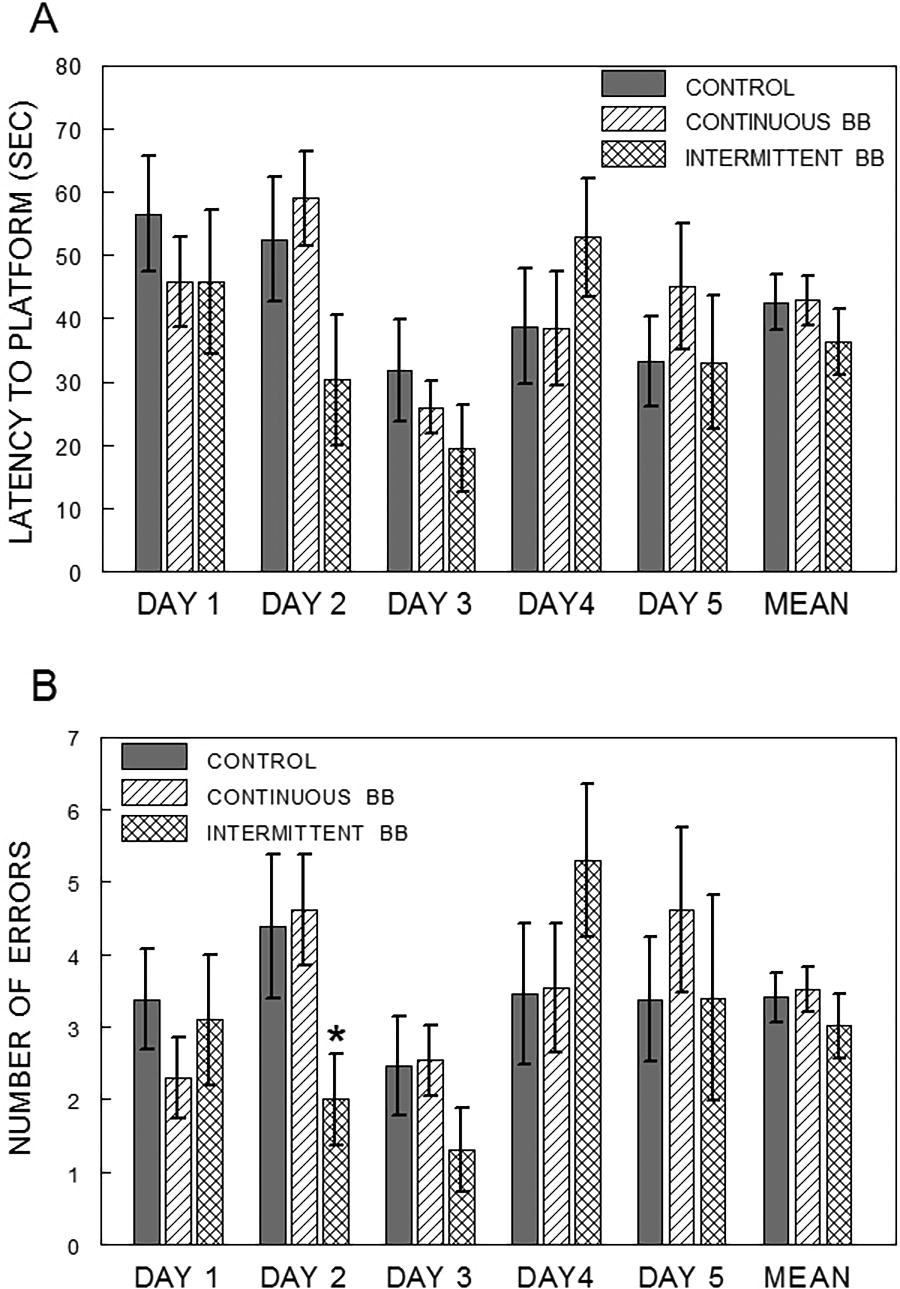
Trial 5 latency (A; mean ± SEM; seconds) and number of total errors (B; mean ± SEM), for days 1–5 and an average of all 5 days (mean), to find the hidden platform in the radial arm water maze in rats fed a control (CON), continuous (CBB), or intermittent (INT) blueberry diet. Asterisk (*) indicates a significant difference from the control group (P < .05) as measured by Dunnett’s test.
Serum inflammation analyses
Results from the in vitro study using the serum as a pretreatment on rat microglial cells showed that blueberry supplementation enhanced anti-inflammation capability, and frequency of intake determined the level of protection against LPS-induced inflammation. Specifically, statistical analyses showed that nitrite concentration had a significant main effect for diet [F(2,474) = 11.27, P < .001] and time [F(1,474) = 32.03, P < .001], and a significant interaction between diet group and time [F(2,474) = 20.10, P < .001] (Fig. 3). Serum from rats fed a continuous BB (CBB) diet significantly attenuated LPS-induced NO production in HAPI microglial cells, but this improvement was not seen in rats fed a control (CON) or intermittent BB (INT) diet. Specifically, post-hoc testing showed that NO was lower after 8 weeks (POST) on the CBB diet, compared to pre-diet levels (PRE) (P < .05). NO was also lower at the POST timepoint in the CBB group, compared to the INT group (P < .05) and the CON group (P < .05), which were not different from each other.
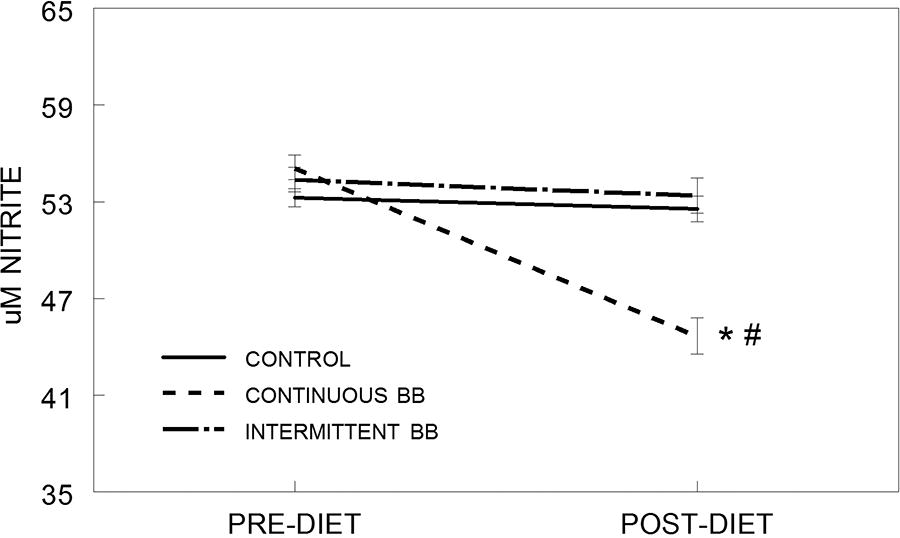
Production of extracellular NO, as measured by μM nitrite, in HAPI rat microglial cells pre-treated with serum from rats both pre-diet (PRE) and after 8 weeks on a control (CON), intermittent (INT), or continuous blueberry (CBB) diet (POST) and stimulated overnight with LPS (100 ng/mL). Data are represented as mean ± SEM (μM nitrite) and quantified using Greiss reagent. Asterisk (*) indicates a significant difference from baseline (PRE, *P < .05); pound (#) indicates a significant difference between diet groups at the same time point (#P < .05) as measured by Fisher’s LSD post-hoc test.
Although COX-2 expression did not show a significant main effect for diet [F(2,363) = 1.28, P > .05], there were significant effects of time [F(1,363) = 196.48, P < .001] and a significant interaction between diet group and time [F(2,363) = 4.10, P < .05] (Fig. 4). Interestingly, serum from rats fed a CON, CBB, or INT diet all significantly attenuated LPS-induced expression of COX-2 in HAPI microglial cells after 8 weeks (POST) compared to pre-diet levels (PRE) (P < .05). However, levels of COX-2 were significantly higher in activated microglia incubated with post-diet serum from CON-fed rats, compared to serum from rats fed with either CBB or INT diets (P < .05), which were not different from each other.
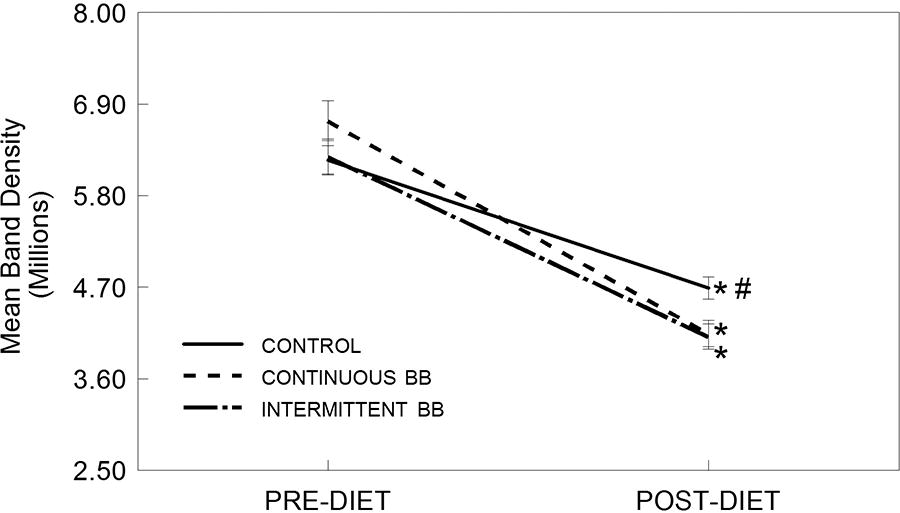
Expression of cyclooxygenase-2 (COX-2, mean band density) in HAPI rat microglial cells pre-treated with serum from rats both pre-diet (PRE) and after 8 weeks on a control (CON), intermittent (INT), or continuous blueberry (CBB) diet (POST) and stimulated overnight with LPS (100 ng/mL). Data are expressed as mean ± SEM of the immunoreactive band density per 20 µg total protein as measured by Western blot. Asterisk (*) indicates a significant difference from baseline (PRE, *P < .05); pound (#) indicates a significant difference between diet groups at the same time point (#P < .05) as measured by Fisher’s LSD post hoc test.
Similarly, iNOS expression did not show a significant main effect for diet [F(2,370) = 0.223, P > .05], but there were significant effects of time [F(1,370) = 15.98, P < .001] and a significant interaction between diet group and time [F(2,370) = 7.43, P < .01] (Fig. 5). Serum from rats fed a CBB or INT diet significantly attenuated LPS-induced expression of iNOS in HAPI microglial cells after 8 weeks (POST) compared to pre-diet levels (PRE) (P < .05). Levels of iNOS were significantly higher post-diet compared to pre-diet in the CON-fed rats, and significantly higher post-diet in CON-fed rats compared to those rats fed with either CBB or INT diets (P < .05), which were not different from each other.
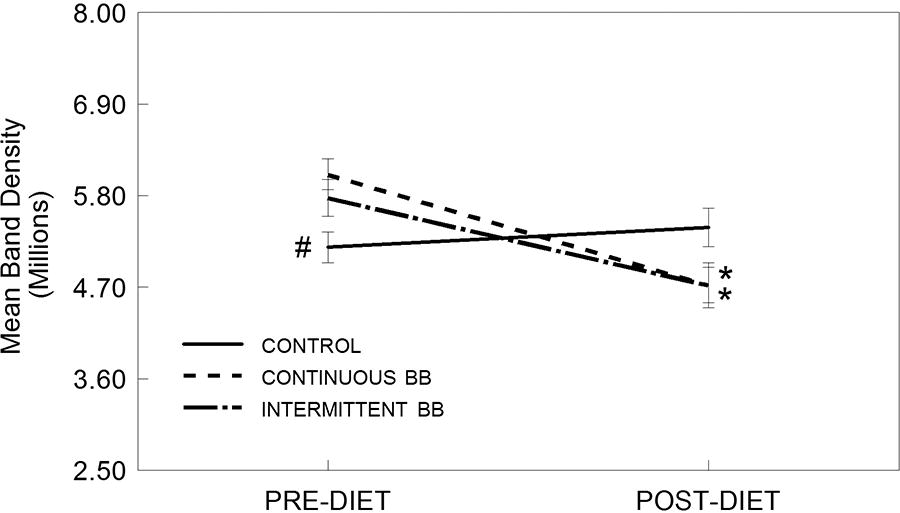
Expression of inducible nitric oxide synthase (iNOS, mean band density) in HAPI rat microglial cells pre-treated with serum from rats both pre-diet (PRE) and after 8 weeks on a control (CON), intermittent (INT), or continuous blueberry (CBB) diet (POST) and stimulated overnight with LPS (100 ng/mL). Data are expressed as mean ± SEM of the immunoreactive band density per 20 µg total protein as measured by Western blot. Asterisk (*) indicates a significant difference from baseline (PRE, *P < .05); pound (#) indicates a significant difference between diet groups at the same time point (#P < .05) as measured by Fisher’s LSD post-hoc test.
DISCUSSION
Overall, results from this study showed that continuous or intermittent feeding of wild blueberries had beneficial effects on activated microglia in vitro, with serum from these rats reducing protein expression of inflammatory enzymes COX-2 and iNOS, but only continuous feeding attenuated nitrite production. As blood was collected ∼48 h after the last blueberry exposure in the intermittent group, it is possible that nitrite in this group may have been improved if blood was collected during one of the days the rats were consuming the blueberry diet, as nitrite is measured in the cell-conditioned supernatant. Intermittent feeding with blueberry improved motor function as measured by the rotarod test, and cognitive function was shown as reduced errors in the water maze on Day 2 only. Surprisingly, continuous blueberry feeding did not improve motor or cognitive function as was seen in previous studies. 7,11,12 It could be that this version of the RAWM was too difficult for the aged rats, as minimal learning was seen from Day 1 to Day 5 (i.e., no significant change in latency). Even though intermittent BB consumption did not lead to improvements in more behavioral measures, the fact that some tests demonstrated beneficial effects and inflammation was reduced by serum in vitro shows that BBs might not have to be consumed daily to attain beneficial effects.
In this study, we examined the in vitro effects of pre-treatment with serum on microglia in an attempt to determine if blueberry metabolites present in the circulating blood may be mediating the anti-inflammatory effects. One of the explanations for improvements with intermittent feeding might be the retention of polyphenols or their metabolites in tissue following consumption. It is thought that polyphenols have low bioavailability because of poor absorption from the intestine and/or extensive metabolism following absorption, however, their metabolites are found in blood and target organs such as the brain. 29 Anthocyanins, one of the main components of blueberries, are relatively unstable and degrade easily. 12 However, long-term consumption of anthocyanins is likely to facilitate their transport, possibly through increased glucose transporter 2 expression, 30 which may account for their beneficial effects in the present study. Additionally, studies have shown that the coupling of efflux transporter and metabolic enzymes controls the disposition of polyphenols in vivo when there is an excess capacity to metabolize them. For polyphenols, these coupled processes enable a dual recycling scheme, which allows the polyphenols to be reabsorbed and results in longer than expected apparent plasma half-lives for these compounds, possibly enhancing their bioavailability. 29
Several studies have shown that polyphenols can cross the blood–brain barrier and accumulate in brain tissue. In one study, pigs who were fed diets supplemented with blueberries for 4 weeks had no detectable anthocyanins in their plasma or urine following an 18–21 fast, yet intact anthocyanins were seen in all tissues measured, including the eye, cortex, and cerebellum. 31 In another study in which rats were fed a 2% blueberry diet for 10 weeks, anthocyanins from the diet were able to cross the blood-brain barrier in glycosylated form, localize in brain regions that mediate cognitive behavior (e.g., cortex and hippocampus), and therefore may have affected cognitive performance by enhancing neuronal signaling pathways associated with learning and memory. 9 In that study, as in the present study, there were no differences between the dietary groups as both the control and BB-fed rats showed equal improvement in learning performance, and similar performance on the probe trials in the Morris water maze, even though there was a relationship between total number of different anthocyanins detected in the brain and maze performance in the blueberry-fed rats. 9
This animal evidence supports the role of polyphenols in human health, and although polyphenols have not been measured in human tissue, studies have compared their fate following acute and chronic consumption. After consumption of just one single dose of 250 mL blueberry juice following a 5-day washout period, anthocyanin metabolites were abundant and well-retained in urine, suggesting enterohepatic recycling leading to prolonged residence time. 32 A study using plasma from healthy men fed wild blueberries acutely or daily for 30 days showed blueberry polyphenols are absorbed and extensively metabolized by phase II enzymes and by the gut microbiota, leading to an array of metabolites that may be individually or collectively responsible for the beneficial effects observed after blueberry consumption. 8 Furthermore, chronic consumption of BB led to the retention and persistence of more phenolic acid compounds over a longer period than was observed following acute consumption. 8 The fact that the inflammatory markers of COX-2 and iNOS were decreased in LPS-activated microglia in the current study following pre-treatment with serum from the intermittent group, which had not consumed blueberry in the previous 48 h, is further evidence for the retention of polyphenols in the body.
Retention of polyphenols in the body has also been shown to be associated with improvements in cognition. In two human clinical berry trials, both blueberry and strawberry consumption for 90 days showed that their polyphenols are absorbed and extensively metabolized, resulting in the production of phenolic acid derivatives and their conjugates. 33,34 Although most phenolic compounds were metabolized and returned to baseline concentrations following an overnight fast, phenolic acids, particularly hippuric acid, were retained in circulation, suggesting potential adaptation. However, an additional 45 days of supplementation did not further increase the concentrations of anthocyanins in plasma. Furthermore, altered plasma concentrations of individual blueberry and strawberry polyphenolics were associated with improvements in cognition. 33,34 In a randomized, double-blind, placebo-controlled trial with blueberry supplementation in older adults with mild cognitive impairment (MCI), cognitive performance was assessed, as were urinary levels of parent anthocyanins and phase 2 anthocyanin metabolites. 35 Even though there was no group difference in urinary excretion of total anthocyanins (parent plus metabolite forms) due to an abundance of phase 2 metabolites in both groups, parent anthocyanins were significantly greater in the blueberry group and were correlated with neurocognitive benefit in semantic access and visual-spatial memory. 35
As suggested in a recent review focusing on anthocyanins and their C6-C3-C6 metabolites, anthocyanin uptake into enterocytes could be impeded by an anthocyanin association with mucin. 36 Evidence for mucin’s effects includes a study showing that when a large blackberry dose preceded a small dose, cyanidin-3-glucoside absorption from the small dose was reduced compared to when the first dose was small. 37 Further evidence is that urinary excretion of anthocyanins declined during a 36-day study of blueberry juice consumption, and excretion was greater for total anthocyanins, but not for parent anthocyanins (which were retained better with once/day dosing) when daily blueberry juice was taken all at once as a 250 mL dose in the morning rather than as three 83 mL doses three times/day over 12 h. 38 These results suggest saturation or inhibition effects of anthocyanins on xenobiotic machinery may contribute to the urinary decline in anthocyanins during prolonged intake and that greater intestinal coverage improved the capacity for anthocyanin absorption. 38 It is possible that in the present study, with the 3-day break from anthocyanin intake, the saturation or inhibition effects of anthocyanins were not reached and absorption was increased overall.
There is an increasing amount of evidence showing that polyphenols exhibit physiological functions via hormesis, wherein there is a stimulatory or beneficial effect at low doses and an inhibitory or toxic effect at high doses. 39 In other words, short-term exposure to blueberry phytochemicals could increase reactive oxygen/reactive nitrogen species or inflammation which, in turn, would lead to upregulation of antioxidant and anti-inflammatory defense mechanisms to enhance long-term protection. 15,39 It is possible that the intermittent diet in this study triggered more hormetic mechanisms than the continuous feeding, therefore it was able to better protect against age-related deficits in behavior. This finding would be in agreement with a study that showed that consuming blueberries for two days induced oxidative stress but continuous feeding ultimately led to reduced oxidation in rats. 15
CONCLUSIONS
In conclusion, BBs may not have to be consumed daily to elicit beneficial effects, possibly due to the retention of polyphenols and their metabolites in tissue and circulation. Future studies should measure tissue and/or serum levels of BB metabolites following continuous vs. intermittent feeding to determine if there is a difference. Because of the lack of availability of older female rats, this study was only conducted in male rats. It is unknown if these results will translate to female rats, so they should be included in future studies. Additionally, other mechanisms for the beneficial effects of blueberries should be studied in relation to frequency of consumption, such as changes to the gut microbiome or brain functions.
Footnotes
DATA AVAILABILITY STATEMENT
All data are available upon request.
AUTHORS’ CONTRIBUTIONS
Conceptualization: M.G.M. and B.S.-H.; Methodology: M.G.M., A.N.C., T.Z., and B.S.-H.; Formal analysis: D.R.F. and B.S.-H.; Investigation: D.R.F., D.S.C., A.N.C., T.Z., and B.S.-H.; Resources: D.R.F. and B.S.-H.; Writing—original draft preparation: B.S.-H.; Writing—review and editing: M.G.M., A.N.C., T.Z., D.S.C., and B.S.-H.; Visualization: D.R.F.; Supervision: B.S.-H.; Project administration: D.R.F. and B.S.-H.; Funding acquisition: B.S.-H. and T.Z. All authors have read and agreed to the published version of the article.
AUTHOR DISCLOSURE STATEMENT
The authors declare to have no conflict of interest.
Disclaimer
Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the USDA. The USDA is an equal opportunity provider and employer. Any opinions, findings, conclusions, or recommendations expressed in this publication are those of the authors and do not necessarily reflect the view of the USDA.
FUNDING INFORMATION
This research was funded by the United States Department of Agriculture (USDA), Agriculture Research Service (ARS) under project 8050-51000-107-000D, and an agreement between the USDA and the