
Abstract
Abstract
Primary open-angle glaucoma (POAG) is a leading cause of blindness worldwide, and intraocular pressure (IOP) is an important modifiable risk factor. IOP is a function of aqueous humor production and aqueous humor outflow, and it is thought that prolonged IOP elevation leads to optic nerve damage over time. Within the trabecular meshwork (TM), the eye's primary drainage system for aqueous humor, matricellular proteins generally allow cells to modulate their attachments with and alter the characteristics of their surrounding extracellular matrix (ECM). It is now well established that ECM turnover in the TM affects outflow facility, and matricellular proteins are emerging as significant players in IOP regulation. The formalized study of matricellular proteins in TM has gained increased attention. Secreted protein acidic and rich in cysteine (SPARC), myocilin, connective tissue growth factor (CTGF), and thrombospondin-1 and -2 (TSP-1 and -2) have been localized to the TM, and a growing body of evidence suggests that these matricellular proteins play an important role in IOP regulation and possibly the pathophysiology of POAG. As evidence continues to emerge, these proteins are now seen as potential therapeutic targets. Further study is warranted to assess their utility in treating glaucoma in humans.
Introduction
P
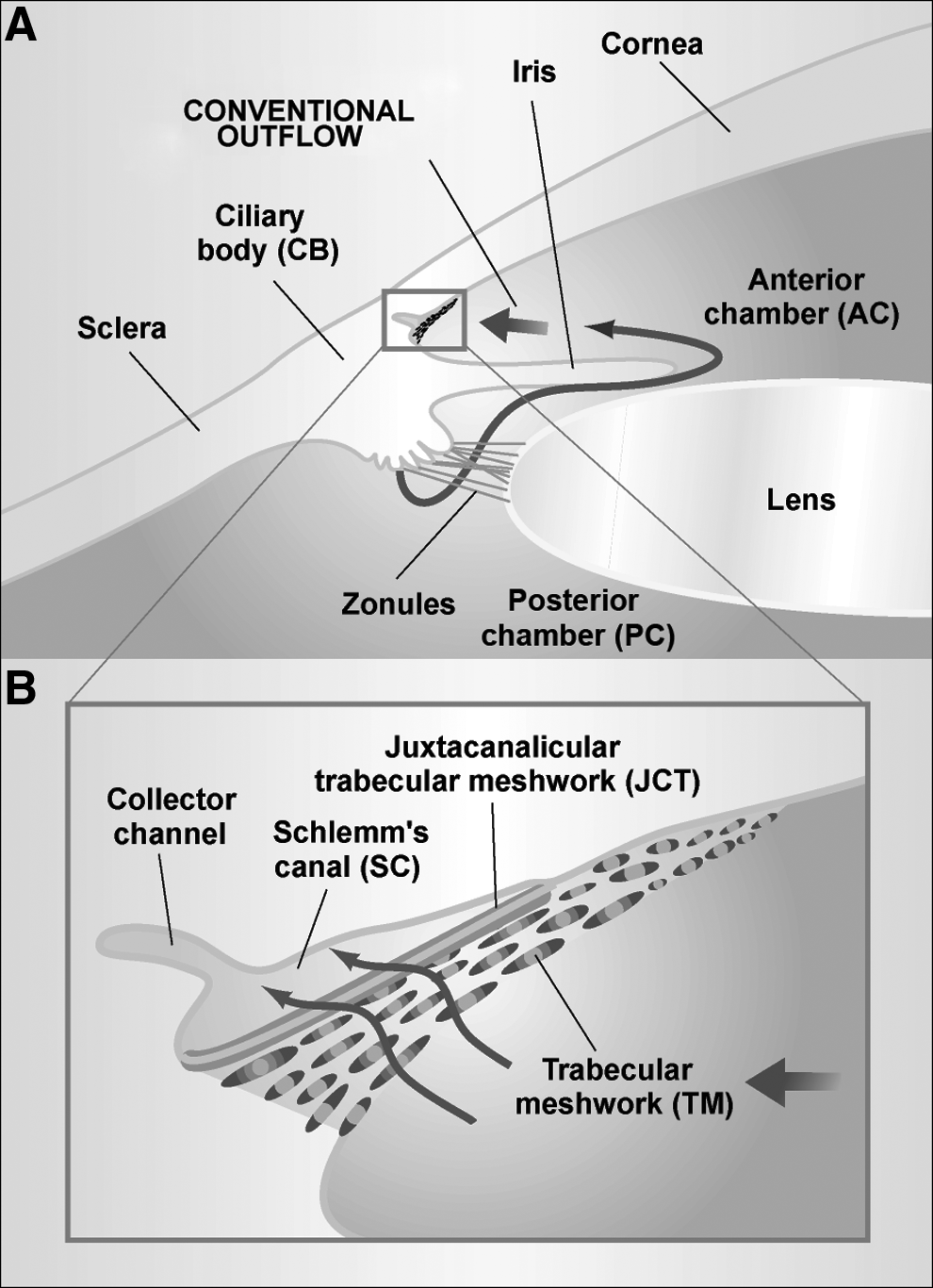
Schematic diagram showing key ocular structures involved in aqueous humor inflow and outflow (adapted, with permission, from Tomarev 2001, Nature Med. 7, 294–295; Copyright © 2001 Nature Publishing Group).
Matricellular proteins are nonstructural secreted glycoproteins that aide in cellular communication with and control over surrounding ECM. This expanding family of proteins includes the prototypical SPARC (secreted protein, acidic and rich in cysteine), also known as osteonectin and BM-40, myocilin, connective tissue growth factor (CTGF), and other CCN family members, thrombospondins-1 and −2 (TSP-1 and −2), tenascins-C and–X, SC1/hevin, and osteopontin.35–38 Galectin family proteins perform similar functions, meeting several matricellular protein criteria, but they lack a high degree of structural similarity to the proteins above. Matricellular proteins are generally characterized by their importance in regulating interactions between structural proteins and signaling molecules such as cytokines and matrix proteases.
Due to the vital role that ECM turnover plays in managing outflow facility, matricellular proteins are of great interest. A growing body of evidence suggests that some matricellular proteins may serve as promising therapeutic targets. This article represents a comprehensive update on recent progress in the rapidly developing field, expanding upon an initial review of matricellular proteins in the TM from several years ago. 39
Refining Our Understanding
Matricellular proteins in the TM
Secreted protein, acidic and rich in cysteine also known as osteonectin or BM-40
The prototypical matricellular protein SPARC was first studied in bone in 1981. 40 Mammalian, amphibian, and avian SPARC demonstrate greater than 70% amino acid sequence homology. 41 Wherever and whenever there is a significant degree of ECM turnover, SPARC is likely to be highly expressed, playing notable roles during both embryogenesis and tissue repair.42,43 In a number of conditions involving aberrant tissue remodeling—such as hepatic fibrosis,44,45 cholangitis, 46 renal interstitial fibrosis, 47 systemic sclerosis, 48 osteoporosis, 49 and certain carcinomas 50 —elevated SPARC expression levels have been noted. Through its interactions with integrin-linked kinase, SPARC has been shown to enhance fibronectin-mediated ECM assembly.51,52 SPARC suppression has been shown to decrease the expression of multiple collagens and tissue inhibitor of metalloproteinase (TIMP)-3.53,54 Mice harboring single gene deletions of Sparc exhibit accelerated dermal wound closure, decreased levels of dermal collagen, elevated adiposity, decreased bone mineral density, early-onset cataractogenesis, and a suppressed reaction to implanted synthetic materials.55–61 In addition, SPARC-null mice exhibit decreased laminin and collagen (types I and IV) deposition within renal tissue, decreasing the risk for damage consistent with diabetic nephropathy. 62 Taken together, these studies of tissues demonstrating suppressed SPARC levels suggest that this particular matricellular protein normally acts to promote ECM deposition.
SPARC induces changes in matrix metalloproteinases (MMPs) in various human tissues and cell types. For example, in malignant human breast cancer (MDA-MB-231, BT549) and glioblastoma-astrocytoma cell lines (U87MG), SPARC increases MMP-2 activation by decreasing TIMP-2 and upregulating membrane-bound 1-MMP (MT1-MMP or MMP-14), the principle inhibitor and activator of MMP-2, respectively, presumably affecting the tumor's tissue invasiveness.63,64 In monocytes, SPARC upregulates MMP-1 and gelatinase B (MMP-9) via prostaglandin synthase-dependent signaling, thereby facilitating leukocytic movement through ECM. 65
In humans, SPARC localizes to a variety of ocular tissues and its levels can act as a marker for processes such as corneal wound repair and cataractogenesis.66–68 Corneal epithelial cells, retinal pigment epithelium, and lens epithelial cells have all been demonstrated to produce SPARC.68–70 Within lens tissue, tight regulation of SPARC levels is required to maintain lens transparency given that both increased SPARC production 71 and complete absence of SPARC60,72 are associated with cataractogenesis. SPARC is also produced by both TM and CB smooth muscle cells73,74 and aqueous and vitreous humor have been noted to contain significant SPARC levels. 68 Immunofluorescence studies of ex vivo human anterior segment tissues reveal that SPARC is distributed throughout the TM and JCT region. SPARC is one of the most highly expressed genes in primary cultured human TM cells, and its levels are elevated in response to mechanical stretch, suggesting that SPARC plays an important role in normal TM function.75,76
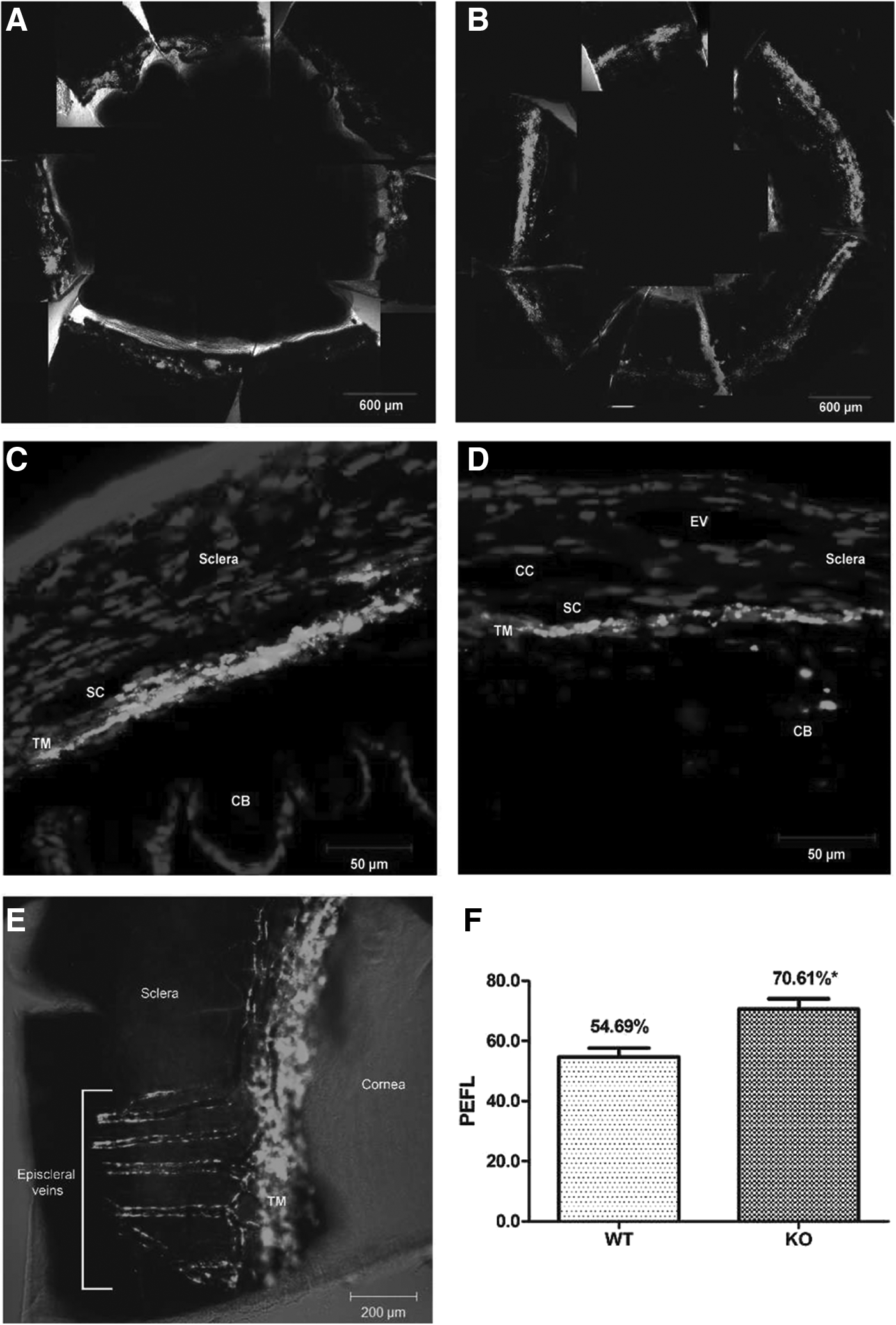
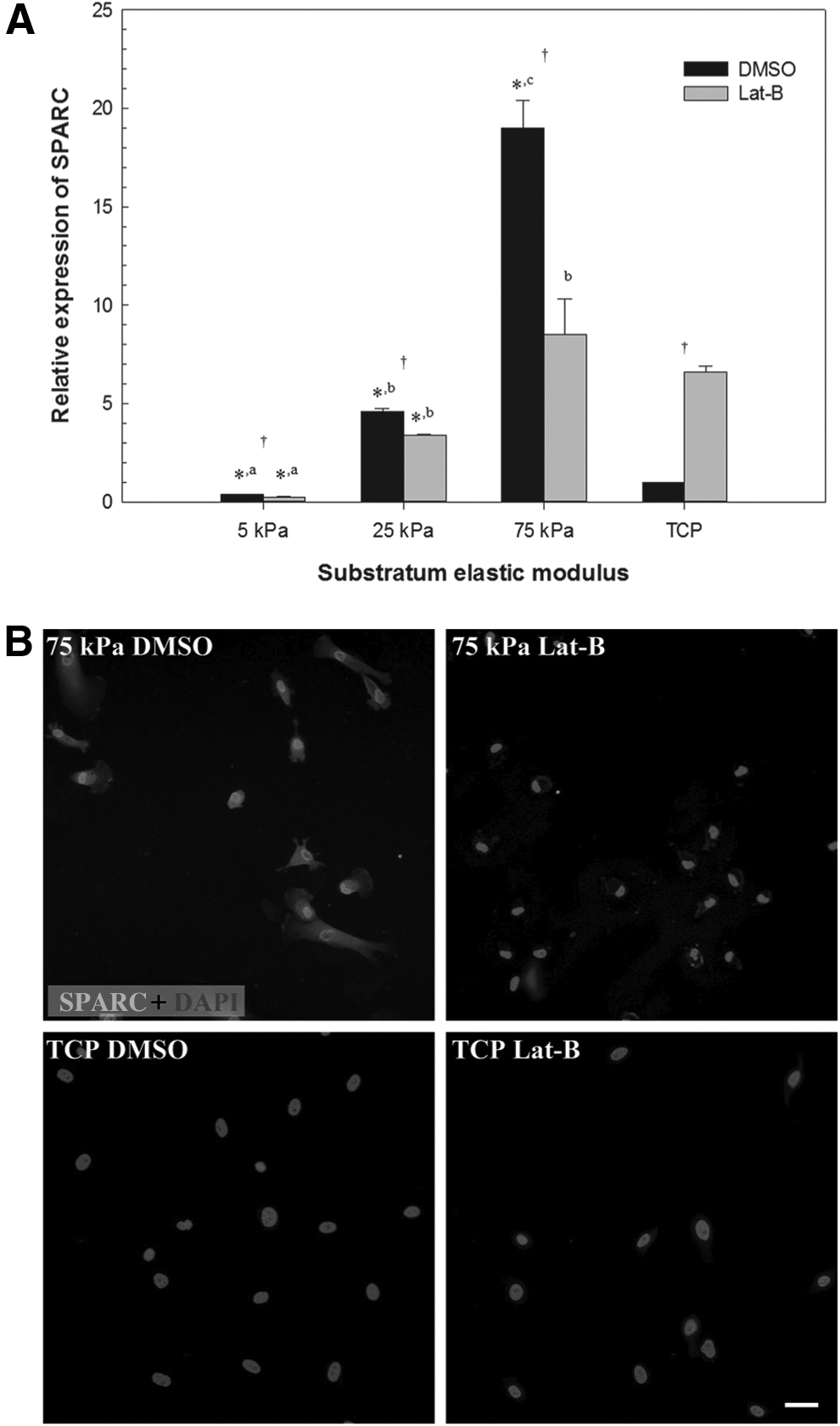
Interest in SPARC as a regulator of ECM homeostasis in the TM continues to grow. Our laboratory has shown that SPARC-null mice exhibit 15%–20% lower IOP compared with their wild-type counterparts77,78 and that these mice demonstrate a more uniform pattern of aqueous humor outflow (Fig. 2). 34 SPARC overexpression in human TM cells increases IOP in an ex vivo perfusion chamber model, likely by increasing the production of various ECM components (Fig. 3). 79 Upstream regulation of SPARC is complex and may involve a combination of microRNA (miRNA) elements,80,81 pro-fibrotic cytokines,82,83 isoprenoid precursors, 84 AMP-activated protein kinase (AMPK), 85 and even dynamic responses to TM stiffness. 86 transforming growth factor (TGF)-β2 cytokine levels are significantly elevated in eyes with POAG,87,88 and treatment with activated TGF-β2 upregulates SPARC in cultured human TM cells (Fig. 4). 83 Of note, preliminary evidence suggests that the TGF-β2-mediated ocular hypertension observed in wild-type mice is not as pronounced in their SPARC-null counterparts. 89 TM cells cultured on synthetic hydrogels mimicking the stiffness found in glaucomatous meshwork show significantly greater SPARC and myocilin mRNA expression than those cultured on hydrogels that approximate the relatively reduced stiffness of normal meshwork (Fig. 5). 86
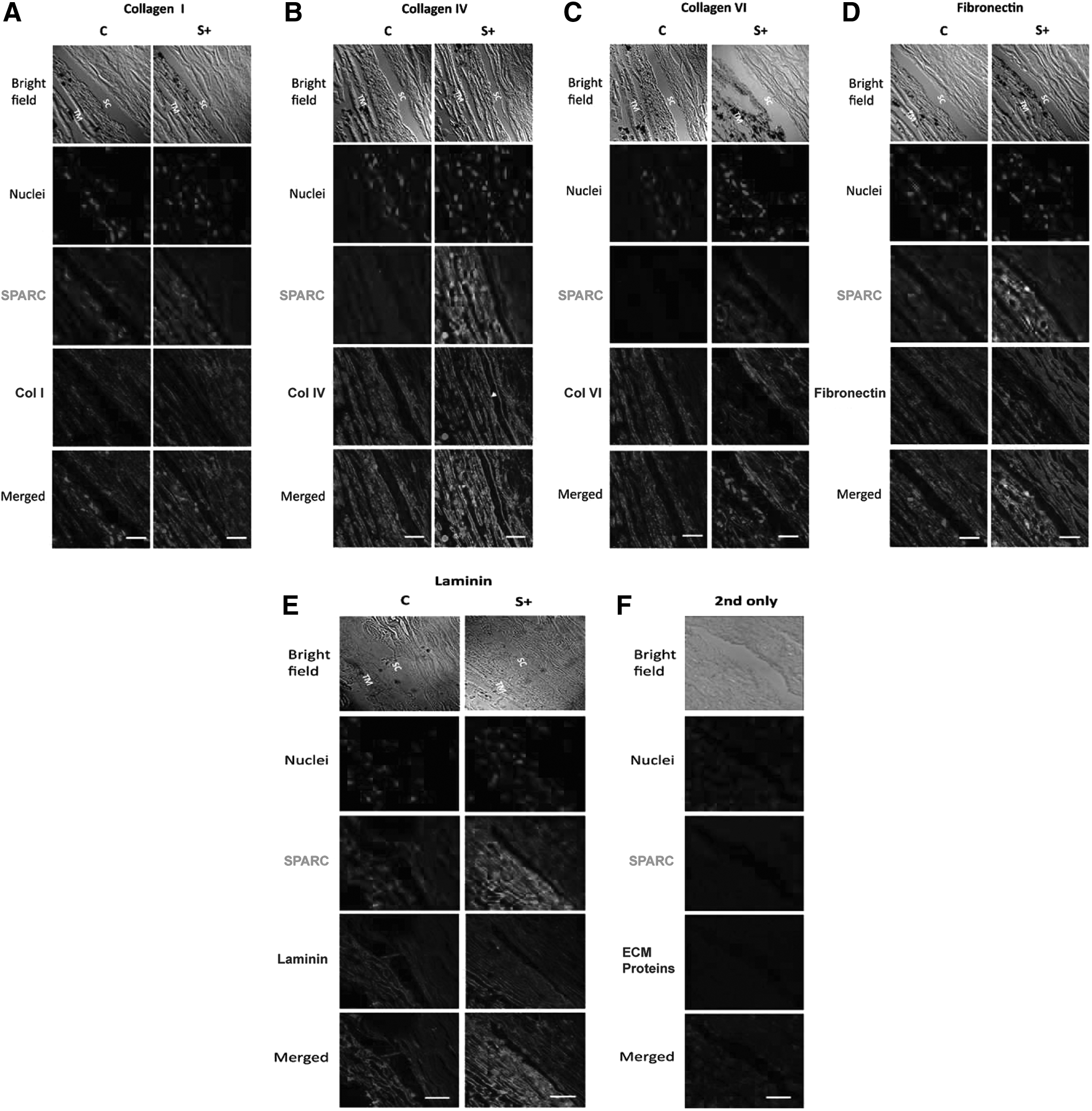
Representative immunolabeling of ECM proteins after infection with adenovirus overexpressing human SPARC versus vector only control (adapted, with permission, from Oh et al.,
79
Copyright © 2013 Association for Research in Vision and Ophthalmology). Collagen I
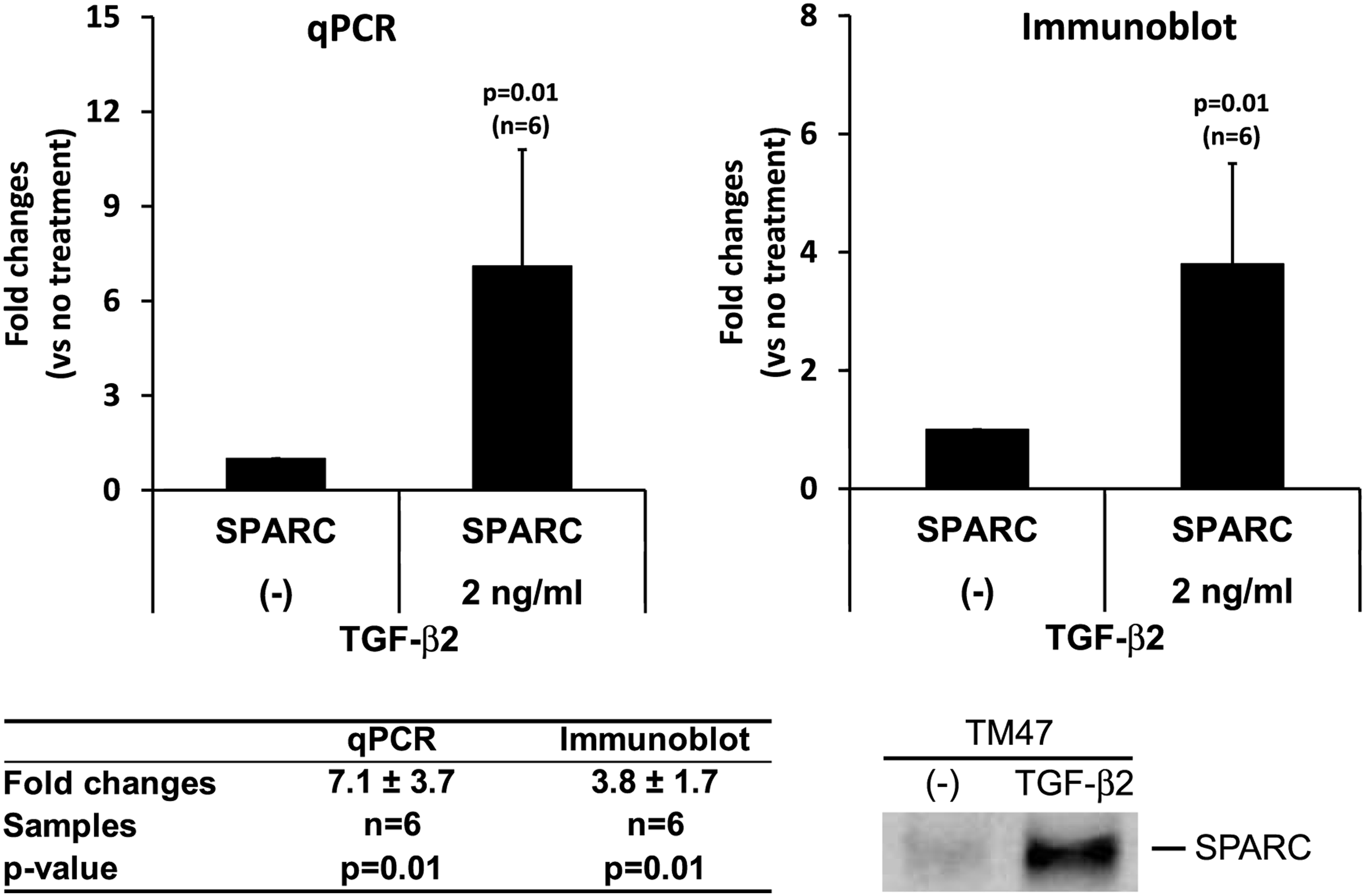
Analysis of SPARC levels following TGF-β2 treatment (2 ng/mL) in cultured TM cells, n=6 (adapted, with permission, from Kang et al., 83 Copyright © 2013 Association for Research in Vision and Ophthalmology). SPARC mRNA was analyzed by qPCR and normalized with β-actin. SPARC protein levels in conditioned media were analyzed by immunoblotting and calculation of integrated band intensity. Representative immunoblot shown.
Myocilin
Myocilin—a secreted protein found in human TM and thought to be involved in ECM regulation 90 —has demonstrated de-adhesive properties that have earned it classification as a matricellular protein.91,92 The gene that encodes myocilin was one of the first to be linked to POAG, 93 and myocilin's C-terminal domain interacts with that of SPARC. 94 Previous studies have suggested that most disease-causing myocilin gene mutations interfere with myocilin protein production 93 or proper folding and secretion,95–97 giving rise to an “intracellular sequestration” model of disease pathogenesis. However, one report found that approximately twenty percent of disease-causing mutants yielded protein products that were still successfully secreted and recovered from culture media. 97 Furthermore, half of those involved mutations in the olfactomedin-like domain near the C-terminal region, which harbors a demonstrated interaction site for SPARC protein. 94 It is therefore reasonable to theorize that altered ECM in the JCT as a result of mutated myocilin may in some cases derive from abnormal interactions with SPARC or other matricellular proteins and may partially explain the pathophysiology of increased resistance to aqueous humor outflow. Taken together, these studies further strengthen the notion that SPARC, like myocilin, may be a key regulatory node that governs ECM homeostasis in the TM and IOP.
Connective tissue growth factor
The CCN family of 6 multifunctional matricellular proteins derives its name from 3 prototypical members named in order of discovery (
Knockout mutations of the gene that encodes CTGF are fatal in mice, and initial work revealed that the protein is crucial for bone and soft tissue development. 102 Studies in human dermal fibroblasts revealed that CTGF may potentiate a cell's ability to spread on fibronectin, a process required for adhesion during tissue development and wound healing. 103 Interestingly, CTGF was also found to play a critical role in regulating fibrosis through its interactions with TGF-β signaling. 104 Given its profibrotic tendencies, it became a protein of interest in surgical models, demonstrating increased expression following glaucoma filtration surgery in rabbits and being associated with an elevated risk of bleb failure.105,106
The putative role of CTGF in human glaucoma pathophysiology was first identified in pseudoexfoliation syndrome,107,108 and attention soon focused on how CTGF regulates TM matrix deposition,109,110 cellular tone, 111 and aqueous outflow facility.112,113 In one promising recent study, the prostaglandin analog fluprostenol was found to mitigate CTGF-induced fibrogenesis in cultured human TM cells. 114 Pharmacological targeting of CTGF therefore shows great therapeutic potential and remains an active area of research. Further work elucidating the potential role of other CCN family members is required.
Thrombospondins
First reported as a thrombin-sensitive protein, TSP—also known as glycoprotein G or GPG—was first described in 1971. 115 Subsequent investigation has led to the adoption of nomenclature that identifies this protein as TSP-1,116,117 one member of a family of proteins that are now classified into subgroups A (TSP-1 and TSP-2) and B (TSP-3, TSP-4, and TSP-5).118–120 The C-terminal regions of TSPs demonstrate shared calcium-, ECM-, and cell membrane-binding properties. Subgroup A TSPs, which all contain an antiangiogenic domain at the N-terminal region, have been found in both nonocular and ocular tissues.119,121 Subgroup B TSPs, which do not clearly belong to the matricellular family, lack antiangiogenic regions and have variable N-terminal domains. 119 Although they are not well characterized, it is believed that TSP-3 may be involved in bone growth plate maintenance 122 ; TSP-4 may be involved in stretch-mediated contractility enhancement within cardiac muscle tissue 123 and spinal sensitization in models of neuropathic pain.124,125 TSP-5, also known as cartilage oligomeric matrix protein, is found in cartilaginous tissue and is extensively reviewed elsewhere.126–128
As they more clearly belong to the matricellular family, TSP-1 and TSP-2 are reviewed here. In vitro studies show that these TSPs, like SPARC, regulate cellular tone, shape, adhesion, proliferation, and apoptosis.129,130 Subgroup A TSPs also play important roles in tissue formation and remodeling.38,129,131 For example, both TSP-1 and TSP-2 have been shown to interact with MMPs, with TSP-1 inhibiting MMP-3-dependent activation of pro-MMP-9 and thrombin-induced activation of pro-MMP-2, 132 and TSP-2 forming a complex with and internalizing MMP-2. 133 Under physiologic conditions, TSP-1 is also capable of binding and activating the pro-fibrotic cytokine TGF-β1,134,135 in addition to binding ECM components such as collagen types I-V, laminin, and fibronectin.136–139 Its role both in physiologic and aberrant tissue repair and fibrosis is reviewed elsewhere. 140 TGF-β1 treatment has been shown to upregulate TSP-1 expression, suggesting the possibility of a reciprocal relationship. 141
Mice harboring single gene deletions of either Thbs1 or Thbs2 have yielded information regarding the 2 protein's functional similarities and differences. TSP-1-null mice demonstrate phenotypes that are generally consistent with states of decreased TGF-β1 activity,142,143 with TSP-1-null and TGF-β1-null mice exhibiting histopathological similarities in at least 9 organ systems. 142 For example, the 2 strains both suffer from reduced dermal matrix, hyperplasia of various epithelial cells, and pulmonary inflammation. Further strengthening the link, cultured TSP-1-null cell isolates exhibit one eighth the amount of active TGF-β1 of their wild-type counterparts, and actively replacing TSP-1 metabolites restores TGF-β1 levels and bronchial epithelial histology in the TSP-1-null cultured cells and mice, respectively. 142 Despite the TSP subgroup A sequence homology, TSP-2-null mice demonstrate phenotypic characteristics that are distinct from their TSP-1-null counterparts. The former demonstrate frail skin, tendon laxity, disordered collagen fibrillogenesis, accelerated wound healing, increased bone density, increased vascular density, and a distinct bleeding diathesis.119,144,145 TSP-1 and TSP-2 also differ in their temporal and spatial expression in knockout mouse models of wound healing. 146 Interestingly, one member of the related ADAMTS (a disintegrin and metalloproteinase with TSP type 1 motifs) family may play a role in the pathogenesis of ectopia lentis,147,148 while another appears to regulate outflow facility in ex vivo perfused human anterior segments. 149
Subgroup A TSPs are expressed throughout a variety of ocular tissues and appear to play a role in several pathologies involving angiogenesis including diabetic retinopathy, age-related macular degeneration, corneal wound repair, pseudoexfoliation syndrome, and uveal melanoma.121,150,151 TSP-1 and TSP-2 are both expressed in human TM,75,121 but TSP-1 localizes to the TM and JCT regions, whereas TSP-2 predominates in the uveal meshwork. 121 In one study, TSP-1 exhibited more intense immunoreactivity in one-third of patients with POAG, 152 making it of particular interest. Of note, both TGF-β1 and TGF-β2 upregulate TSP-1 in human TM cells,152–154 and treatment with TGF-β2 treatment generally favors ECM accumulation and an increase in resistance to aqueous outflow.153,155–158 Furthermore, TSP-1—like SPARC—may play a role in TM cellular tone, as it has been shown to be antagonized by the actin cytoskeletal disruptor latrunculin A.
We have begun preliminary investigations into the ocular phenotypes of TSP-1-null and TSP-2-null mice, given the demonstrated importance of the subgroup A TSPs in matrix remodeling throughout the body and, in particular, within the TM. The average IOPs of TSP-1-null and TSP-2-null mice are approximately 10% and 7% lower than that of their corresponding wild-type mice, respectively, while maintaining similar central corneal thickness (CCT) and iridocorneal angle architecture. 159 These findings were confirmed in a study summarizing IOP changes in all matricellular null strains, which also demonstrated that CCT need not be measured to evaluate IOP differences in these strains (Table 1). 78 Interestingly, aqueous humor turnover rates—as measured by fluorophotometric techniques—are enhanced in both TSP-1- and TSP-2-null mice, suggesting that both may play roles in regulating aqueous outflow facility.
Adapted, with Permission, from Chatterjee et al., 78 Keller et al. 196 Copyright © 2013 Elsevier B.V.
P<0.0001.
WT, wild-type; KO, knockout; n, sample size.
Tenascin-C and tenascin-X
The first in its family to be characterized, Tenascin-C was initially described in the stroma of gliomas and in tendons.160,161 Since then, the vertebrate tenascin family has grown to include 3 additional members. Each of them is comprised of linked subunits that each contain EGF-like and heptad repeats, fibronectin type III domains, and a C-terminal globular domain shared with the fibrinogen family.36,162 Because tenascin-R has thus far only been described in the central nervous system 163 and tenascin-W has yet to be well characterized, 164 tenascin-C and tenascin-X are the only 2 family members currently classified as matricellular proteins.38,130 Generally, tenascins are involved in tissue remodeling and are highly expressed during development and processes involving injury and wound repair. Although they both demonstrate cellular adhesion-modulating properties and the ability to affect cellular tone, migration, and growth, tenascin-C and tenascin-X exhibit significant differences in their organ distribution and overall function.
Tenascin-C is primarily expressed in tendons, ligaments, cartilage, and the ECM of tumor stroma and blood vessels.160,161,165–167 Like its fellow matricellular proteins SPARC and TSP-1, tenascin-C regulates cellular adhesion and migration in a tissue-dependent manner. 168 One mechanism by which this occurs may include interference with integrin-mediated cellular interaction with fibronectin.169,170 At sites of inflammation, tenascin-C is found to be expressed relatively early,171,172 and it appears to be upregulated by TGF-β2 and other anti-inflammatory cytokines173–176 Enhanced expression of tenascin-C has also been observed in response to mechanical stretch,177,178 for example, in models of cardiomyocyte damage and regeneration.179,180 Mice deficient in tenascin-C have abnormal neurochemistry and demonstrate abnormal dermal and corneal would healing with reduced fibronectin.181–184 These phenotypes presumably result from aberrant cell migration and adhesion and disruption of ECM formation.
In healthy mammalian adults, tenascin-C is expressed at the limbus, at the scleromuscular junctions, and in the JCT region of the TM.185–187 It has also been shown to be expressed during normal corneal development. 188 In disease states, the protein is found in the vitreous of patients with vitreoretinopathy and proliferative diabetic retinopathy189,190—likely contributing to disease progression 191 —and in the optic nerve head of rat glaucoma models. 192 Within the TM, tenascin-C is specifically present in sheaths and sheath-derived plaque cores 186 and co-localizes with fibronectin in vitro. 193 Mechanical stress of human TM leads to upregulation and alternative splicing of tenascin-C.76,194 Initial evidence from cultured human TM cells also suggested that tenascin-C may contribute to Rho GTPase-mediated resistance to aqueous humor drainage. 195 One recent study in perfused ex vivo human and porcine anterior chambers has confirmed its predominance in the JCT region and revealed a potential role in the physiologic maintenance and remodeling of TM but found that its RNA interference-based suppression did not affect outflow facility significantly. 196 Furthermore, tenascin-C-null mice exhibit IOPs and CCTs similar to their wild-type counterparts, supporting the notion that the protein—while likely to be involved in dynamic TM maintenance—may not significantly contribute to IOP regulation. 196
Tenascin-X is generally found in the connective tissue of cardiac, skeletal muscle, and skin, and its distribution is often distinct from that of tenascin-C. 197 Patients harboring mutations and deletions of tenascin-X exhibit phenotypic similarities with those who have Ehler–Danlos syndrome, including hyperelastic skin, increased joint and vascular laxity, and abnormal wound healing.198–201 Tenascin-X is believed to modulate cellular adhesion and collagen stiffness.202,203 Mice deficient in tenascin-X, like their human counterparts, exhibit skin hyperextensibility and fragility, with histologically normal-appearing collagen fibrils that are less densely packed. 204 This mouse strain is now recognized as a suitable model for Ehlers–Danlos syndrome. 205 Dermal fibroblasts lacking the gene that encodes tenascin-X demonstrate normal levels of collagen production in vitro but poor collagen deposition in the cell-associated matrix. Although it is not completely understood how tenascin-X mediates ECM deposition, it is thought that its binding to decorin may be crucial, since deficiency of this proteoglycan is also associated with Ehlers–Danlos-like phenotypes.162,206
In ocular tissues, tenascin-X is expressed both during development and in the normal adult cornea, corneoscleral junction, and limbal and conjunctival blood vessels. 207 Of note, the region encompassing its gene has been implicated in one recent genome-wide association study of age-related macular degeneration. 208 Tenascin-X-null mice are now well characterized,186,209–211 and similar to tenascin-C-null mice, they exhibit IOPs and CCTs comparable to their wild-type counterparts, making it less likely that deficiency of this protein significantly contributes to IOP regulation. 196 For this reason, SPARC, TSP-1, and TSP-2 appear to be more promising with regard to potential therapeutic intervention.
Hevin
Hevin was originally identified from among other synaptic junction glycoproteins in the rat brain and was found to share a high degree of homology with SPARC. 212 Like SPARC, hevin demonstrates anti-adhesive properties, 213 which may potentially be mediated by antagonizing fibronectin-mediated adhesion. 214 Furthermore, it has been shown to modulate decorin and collagen fibrillogenesis and to bind collagen I.215,216 Hevin has been shown to help mediate the progression of foreign body-induced inflammation while SPARC directs the formation of a collagenous capsule.51,52 This suggests that hevin may have similar pro-fibrotic functions as its matricellular family member SPARC.
Hevin has been found to be expressed in murine ocular tissue. 217 It was originally perceived as a potential key player in regulation of ECM turnover within the TM and JCT, in part due to its notable C-terminal interaction with myocilin, 218 something it shares in common with SPARC. 94 In human 293-T cells, Li et al. showed that hevin interacts with myocilin and that myocilin causes accumulation of hevin. 218 Of note, patients with myocilin-associated POAG—which demonstrates an over 90% penetrance in heterozygous individuals—generally present with high IOP. 219 Since our original review of matricellular proteins, we have demonstrated that both human and murine TM tissue contain detectable levels of hevin protein. 220 Hevin-null mice exhibited similar IOPs and CCTs to their wild-type counterparts, with similar aqueous humor turnover rates. Furthermore, in cultured human TM cells, TGF-β2 did not induce hevin; this is in contrast to SPARC, which exhibits a reproducible dose-dependent induction. 220 Taken together, these results diminish the likelihood that hevin plays a critical role in glaucoma pathophysiology by altering resistance to aqueous outflow.
Osteopontin
Originally called sialoprotein, 221 osteopontin owes its present name to its potential role as a “bridge” (from the French, pont) between cells and hydroxyapatite.221,222 Osteopontin is now widely considered a member of the matricellular protein family, owning to its observed role in regulating ECM turnover in various tissues.38,130 It still, however, has a structural role in bone, 223 as evidenced by its various binding sites for integrin, hydroxyapatite, and calcium.224,225 In addition to these active domains, it harbors bindings sites for heparin, putative thrombin cleavage sites, and an N-terminal region that is both chemotactic for neutrophils and associated with carotid plaque formation.224–227 Osteopontin's diverse effects—including during development, bone resorption and calcification, wound healing, mucosal protection, and immunological responses, all of which are reviewed elsewhere223,228—have been conceptually grouped into 2 broad functions: regulation of acute and chronic inflammation, and regulation of macrophage-derived cells such as osteoclasts. 229 These putative functions are more or less confirmed by phenotypic traits found in osteopontin-deficient mice, including reduced bone resorption, increased dystrophic calcification, altered sympathetic control of bone mass, impaired periprosthetic osteolysis, abnormal matrix reorganization following tissue injury, and reduced T-cell-mediated immunity.230–235
Within the eye, osteopontin is involved in corneal and choroidal wound healing, lens epithelial injury, autoimmune uveitis, and proliferative diabetic retinopathy.236–242 Using cDNA microarrays, TM cells have been shown to express osteopontin. 243 Given that TM cells have phagocytic activity that may alter outflow facility, osteopontin may be involved in this process. Interestingly, the relative concentrations of osteopontin in the aqueous humor of patients with pseudoexfoliation syndrome is lower than those found in control patients. 244 Additionally, osteopontin exhibits an age-dependent increase in aqueous humor and abnormal levels are a risk factor associated with degeneration of the optic nerve in D2J mice. 245 We found that osteopontin is present in human TM and exhibits lower concentrations in the aqueous humor of POAG patients. 246 There was, however, no significant IOP difference between osteopontin-null mice and their wild-type counterparts. Further investigation is warranted to investigate whether manipulation of osteopontin levels affect outflow facility.
Lectins and galectins
Lectins represent a family of proteins—each containing a subtype-specific carbohydrate recognition domain (CRD)—that are neither catalytic enzymes nor immunoglobulins.247,248 Lectins that specifically bind β-galactosides are now termed galectins, having previously been known as S-type lectins. Galectins are thought to be involved with cell adhesion, migration, and survival.249,250 Thus far, 15 galectins have been identified and are subdivided according to structural characteristics: (1) Monovalent (prototypic) galectins contain a single CRD; (2) bivalent galectins contain tandem-repeats; and (3) chimeric galectins contain a single CRD and a unique N-terminal region rich in proline and glycine. Galectins, which are nonstructural components of the ECM that direct cell-to-cell and cell-to-matrix interactions, are now widely accepted as matricellular proteins.249–251 The viability, fertility, and relatively normal appearance of mice with genetic deletions of single galectins suggests that there is a certain degree of redundancy within the family.252,253
To date, galectin-1 and galectin-3 are the most well described in humans, although progress is being made on all fronts. 254 Galectin-1 is the prototypical galectin and is capable of binding numerous carbohydrate residues255,256 and fibronectin and laminin,257,258 mediating both intra- and intermolecular cross-linking. TGF-β2 increases galectin-1 expression in endothelial cells,255,256 and the effects of galectin-1 on cell adhesion are dose-dependent 259 while its effects on cell growth are biphasic.260,261 The only chimeric galectin identified thus far, 262 galectin-3 binds to a variety of ECM components such as fibronectin, laminin, and collagen IV.263,264 Of note, galectin-3 has also been shown to be either a substrate or an upstream regulator for MMP-1, MMP-2, and MMP-9. These MMPs are important regulators of ECM turnover during pathologic tissue remodeling events, such as those that occur during modulation of the tumor microenvironment or in response to cardiomyocyte damage in heart failure patients.265–270 With regard to cellular proliferation in neoplastic and autoimmune disease, galectin-1 and galectin-3 often promote antagonistic agendas, with galectin-1 mediating apoptosis and galectin-3 mediating cell survival.250,271,272
Galectin family proteins have been implicated in a variety of ocular disorders, including inflammatory conditions of the conjunctivita and cornea, proliferative vitreoretinopathy, and diabetic retinopathy.273–278 Numerous studies suggest that galectins are important in the maintenance of a healthy ocular surface at rest and after injury,279,280 with galectins-1 and −9 specifically playing roles in the response to bacterial and viral pathogens that damage the corneal surface.281–283 The healthy human lens 284 also expresses galectin-3, and a galectin-like molecule called GRIFIN (galectin-related inter-fiber protein).285,286 In depth proteomic analysis of normal human CB tissue—which participates in aqueous humor production and aqueous humor outflow through the unconventional pathway—has revealed expression of galectin-1. 287 Additionally, there is now evidence that suggests that galectin-3 may play a role in neuroprotection against glaucomatous damage.288,289
Galectins—specifically galectins-1, -3, and -8—may be involved in the TM.290–292 Fautsch et al. have confirmed the presence of galectins-1 and -3 RNA and protein using PCR, western blot analysis, and immunohistochemistry. 290 Both galectins were observed to stain throughout the TM, including the uveal, limbal, and JCT regions, and the inner and outer walls of Schlemm's canal. Of note, the authors found no visible difference in the galectin-1 or galectin-3 labeling distributions of glaucomatous versus normal TM tissues. Diskin et al. found that TM cells express and adhere to galectin-8-coated wells but do not adhere to galectin-1 or −3-coated wells, which was supported by their discovery that TM cell β1-integrins carry predominantly α2-3-sialylated glycans, which exhibit high affinity for galectin-8 specifically. These studies provided the foundation for a recent investigation that revealed that galectin-8 promotes cytoskeletal rearrangement in TM cells through activation of Rho signaling, which presents the intriguing possibility that galectin-8 may indeed participate in the regulation of aqueous outflow facility. 292 Mice with deletions of the genes that encode galectin-1 and -3 have been thoroughly characterized since being engineered over 2 decades ago,252,253,293 but galectin-8-null mice have not been widely studied or described. Future studies aimed at assessing outflow facility and IOP in galectin-null mice may provide insight into the functional importance of galectin family proteins. Of the matricellular proteins reviewed in this article (Table 1), 78 galectins present the greatest number of unanswered questions in this regard.
Conclusions
Matricellular proteins represent an intriguing family of nonstructural proteins that affect ECM turnover within the TM, just as they do in nonocular tissues. Some of these proteins antagonize fibronectin binding, thereby interfering with cellular adhesion. All of them share a certain pro-fibrotic theme, interacting with cytokines such as TGF-β2, whose link to glaucoma pathogenesis is now widely accepted. Over the past 5 years, significant strides have been made in our understanding of the role of these matricellular proteins in the promotion of ECM deposition and regulation of outflow facility within the TM. With recent studies into SPARC, myocilin, CTGF, TSP-1, TSP-2, tenascin-C, tenascin-X, hevin, osteopontin, and galectins, it is clear that several of these proteins show promise as possible therapeutic targets for glaucoma. SPARC, myocilin, CTGF, TSP-1, TSP-2, and galectins show the greatest promise thus far and warrant further study.
Footnotes
Acknowledgments
Howard Hughes Medical Institute Research Training Fellowship for Medical Students (A.C.), National Eye Institute EY 019654-01 (D.J.R.) and EY 014104 (MEEI Vision-Core Grant).
Author Disclosure Statement
No competing financial interests exist.