
Abstract
Abstract
Purpose:
The involvement of local and systemic oxidative stress in intraocular pressure (IOP) elevation and optic nerve damage has been hypothesized in the pathogenesis of glaucoma. In this study, we aim to evaluate the antioxidant effects of curcumin in BV-2 microglia oxidative damage and assess its neuroprotective effects in a chronic high IOP rat model.
Methods:
BV-2 microglia cell line was used in an in vitro study and Wistar rats were used in an in vivo study. Cultured BV-2 microglia cells were pretreated with 10, 1, or 0.1 μM curcumin for 1 h, and sustained oxidative stress was induced by subjecting BV-2 microglia to 200 μM hydrogen peroxide (H2O2) for 24 h. MTT assay was used to determine cell viability. Changes of intracellular reactive oxygen species (ROS) and apoptosis were analyzed by flow cytometry. Three episcleral veins were cauterized to induce high IOP in Wistar rats and measured by Tonopen. After 6 weeks of treatment with curcumin (10 mg/kg/day) by intragastric administration, surviving of retinal ganglion cells was quantified. Activation of caspase 3, cytochrome c, BAX, and BCL2 was quantified by Western blotting both in BV-2 microglia and in animal model. Data were analyzed with the GraphPad Prism 5.0 software, and P<0.05 was considered to be statistically significant.
Results:
The in vitro study showed that when BV-2 microglia was pretreated with curcumin, the cell viability increased and the intracellular ROS and apoptosis significantly decreased. In the in vivo study, chronic mild IOP elevation was induced for 4 weeks. In the curcumin-treated group, curcumin protected rat BV-2 microglia from death significantly. In both H2O2-treated BV-2 microglia and glaucoma models, caspase 3, cytochrome c, and BAX were downregulated and BCL2 was upregulated in the curcumin-treated group.
Conclusions:
Curcumin affords neuroprotective effects by inhibiting oxidative damage and could be a new or adjunctive treatment for glaucoma.
Introduction
G
Curcumin (diferuloylmethane), is the principal curcuminoid of the popular Indian spice turmeric, which belongs to the ginger family (Zingiberaceae). 7 In recent studies, curcumin showed antioxidant, antitumor, chemopreventive, and anti-inflammatory action in acute and chronic inflammation, both in clinical practice and in experimental models.8–10 The anti-inflammatory and antitumor properties of curcumin were linked to its ability to downregulate NF-κB activation, IκBα gene expression, cyclooxygenase-2 gene, prostaglandin E-2, interleukins, tumor necrosis factor-α, and so on. 11 Also, the effect of curcumin on amyloid beta (Aβ) plaque buildup, a major cause of Alzheimer's, has been studied and showed protective effects.12,13 Curcumin's anti-inflammatory properties are involved in the neuroinflammatory cascade of neurotic plaque pathogenesis. 14 In some respects, glaucoma is similar to Alzheimer's disease. 15 The potential of curcumin in the treatment of glaucoma has not yet been studied.
In the present study, we evaluate the neuroprotective efficacy of curcumin both in vitro and in vivo. We show that curcumin protects BV-2 microglia cell viability and inhibits intracellular reactive oxygen species (ROS), apoptosis, and the activation of caspase 3 under hydrogen peroxide (H2O2) oxidative damage. In the high IOP rat model, curcumin is found to protect against the loss of RGCs also. All of these results suggest that curcumin could be a potential treatment strategy for glaucoma.
Methods
Cells and animals
BV-2 microglia was presented by the School of Basic Medicine, Peking Union Medical College, 16 and were grown in Dulbecco's modified Eagle's medium: Nutrient Mixture F-12 (Gibco, Grand Island, NY) containing 10% fetal bovine serum (Gibco) and 1% penicillin–streptomycin (100 U/mL penicillin and 100 ng/mL streptomycin) at 37°C and 5% CO2.
All work with animals respected the guidelines set out by the ARVO Statement. Wistar rats (250–300 g, female) were obtained from the animal center of the Capital Medical University and raised in the animal room of the university with food and water ad libitum. Deep anesthesia (ketamine, xylazine, acepromazine injected intraperitoneally; 50/5/1 mg/kg, as per IACUC recommendations) was used for the high IOP model, RGC labeling, and euthanasia. For measuring IOP, light anesthesia was used (a gas mixture of oxygen, 2% isoflurane mixture, at a rate of 2.5 L/min, as per IACUC recommendations).
Curcumin pretreatment and chronic oxidative stress
BV-2 microglia cells at ∼80%–90% confluence were pretreated with curcumin (10, 1, and 0.1 μM) and incubated at 37°C for 1 h. Oxidative stress was induced by treating BV-2 microglia cells with 200 μM H2O2 in a serum-free medium for 24 h. 17 Untreated cells and cells incubated with H2O2 alone were used as normal control and H2O2 control, respectively.
Evaluation of cell viability
Cell viability was determined by an MTT test. BV-2 microglia cells were plated in a 96-well plate at a density of 1×104 per well for 24 h. After pretreatment with curcumin for 1 h, cells were incubated with 200 μM H2O2 for 24 h. After incubation, the cells were washed twice with phosphate-buffered saline (PBS), and the MTT was added to each well. The plates were incubated at 37°C for 4 h, 100 μL of dimethyl sulfoxide (DMSO) was added, and the plates were shaken gently for 15 min to solubilize the formazan blue crystals. Absorbance at 570 nm was measured with an ELISA plate reader (Dynatech Medica, Guernsey, United Kingdom). Each experiment was performed in 3 wells and was duplicated at least 3 times.
Measurement of intracellular ROS
Intracellular ROS of BV-2 microglia was evaluated with the fluorescent probe H2DCFDA according to the manufacturer's instructions. 17 Cells were pretreated with curcumin for 1 h and incubated with H2O2 for 24 h. Cells were harvested and suspended in freshly prepared 1 μM H2DCFDA at 37°C for 30 min in the dark. After rinsing with PBS, cells were analyzed immediately by flow cytometry (ex/em=488/530 nm).
Flow cytometry analysis of BV-2 microglia apoptosis by Annexin V and PI
Apoptosis of the BV-2 microglia cells (FITC Annexin V Apoptosis Detection Kit; BD Science, Franklin Lakes, NJ) was evaluated according to the manufacturer's instructions. 18 The BV-2 microglia cells (1×106) were seeded in 6-well plates and incubated with H2O2 for 24 h after pretreatment with curcumin. The samples were analyzed by flow cytometry (FACSCalibur; BD Biosciences), and the experiments were performed in triplicate.
High IOP rat models and drug treatment
As previously described, the high IOP rat model made by cauterizing 3 episcleral veins (2 dorsal episcleral veins located near the superior rectus muscle and 1 temporal episcleral vein located near the lateral rectus muscle) with a 30" cautery tip under deep anesthesia. 19 The contralateral control eyes had sham surgery to isolate the 3 veins only but without cauterization. The rats were intragastrically administrated with curcumin (10 mg/kg/day), once a day for 42 consecutive days. 20 During the experiments, no abnormal behavior or symptoms were observed and no significant changes in the body weight. The animals were divided into 4 groups (each 6 rats): normal control group, a high IOP control group, PBS-treated high IOP group, and curcumin-treated experimental group (10 mg/kg).
IOP measurements
IOP was measured by Tonopen XL applanation tonometer immediately after the episcleral vein cauterization surgery and every week until the endpoint of each experiment. Normal IOP was 10–14 mm Hg according to other reports 21 and our experiments. In our experiments, 3 consecutive readings were obtained from each eye, and the average number was taken as the IOP. In this high IOP animal model, the IOP was elevated to ∼1.7-fold for use in this study, and cauterized eyes in which an average IOP of<1.4-fold or >2.8-fold occurred at any point during the study were excluded. We also discarded the animals in rare cases, in which the retinal vasculature showed signs of ischemia.
RGC retrograde Fluorogold labeling
RGCs were retrogradely labeled with a 4% Fluorogold solution (Fluorochrome, Englewood, CO) applied bilaterally to the superior colliculus (SC), as described earlier. 21 Rats were mounted on stereotactic apparatus, holes were drilled besides the lambda suture on each side, and Fluorogold (1 μL) was injected into the SC. In the high IOP model, retrograde labeling was performed at day 35 after ocular hypertension (7 days before the experimental endpoint).
RGC survival quantification
Quantification of surviving RGCs was performed as reported previously. 21 On day 42 of high IOP, both eyes were enucleated, the anterior parts were cut out, and the remaining part was fixed in 4% paraformaldehyde (PFA) for 30 min. Then, retinas were flat-mounted on a glass slide and dissected by 4 radial cuts to facilitate flattening of the retinas. Using a Zeiss fluorescence microscope (Carl Zeiss Meditec, Jena, Germany), 3 pictures (at a distance of 1, 2, and 3 mm radially from the optic nerve, indicated as areas 1, 2, and 3) were taken in each retina quadrant (superior, temporal, inferior, nasal) 20 ; therefore, 12 pictures/retina at 20× magnification were taken of the flat-mounted retinas. In all cases, manual RGC counting was performed by 2 independent persons. Standardized %RGC survival in each rat was calculated as the ratio of the experimental right eye (OD) to the control left eye (OS) (RGCexperimental/RGCcontralateral).
Measurement of caspase 3, cytochrome c, BAX, and BCL2 activity by Western blot in BV-2 microglia and glaucoma animal model
Cells were pretreated with curcumin for 1 h and incubated with 200 μM H2O2 for another 12 h. Retinas were dissected and lysed in 80 μL sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) sample buffer containing 2% SDS. Twenty micrograms of samples was loaded onto a 10% SDS-PAGE gel and electrophoresed at 150 V for 1 h. Proteins were then transferred to a polyvinylidene difluoride (PVDF) membrane (Bio-Rad, Hercules, CA). After blocking, anti-caspase 3, anti-cytochrome c, anti-BAX, and anti-BCL2 monoclonal antibodies (1:1,000; Cell Signaling Technology, Danvers, MA) and anti-β-actin antibody (1:5,000; Cell Signaling Technology) were applied. Bands were visualized by incubation with a horseradish peroxidase-conjugated goat anti-rabbit or anti-mouse secondary antibody (1:10,000; Multiscience Biotech, Hangzhou, China) and chemiluminescence substrates (ECL Plus; PerkinElmer, Inc., Covina, CA). Blots were developed and analyzed with the ImageJ software. The protein levels were normalized by β-actin.
Statistical analysis
The data analysis was performed using the statistical software Prism 5 (GraphPad Software, Inc., San Diego, CA). All of the data were presented as the mean±SD. The differences were evaluated using ANOVA followed by the Student–Newman–Keuls test for multiple comparisons and the Student's t-test for pairwise comparisons. A P<0.05 was considered to indicate a statistically significant difference.
Results
Curcumin protected BV-2 microglia cells against H2O2-induced cytotoxicity
The MTT assay is used for assessing BV-2 microglia cell viability. Incubation of BV-2 microglia cells with 200 μM H2O2 for 24 h resulted in significant cell death. BV-2 microglia cells pretreated with curcumin were found to protect BV-2 microglia cells against H2O2-induced cytotoxicity dose dependently (10, 1, and 0.1 μM) compared with H2O2-treated controls (P<0.05, Fig. 1).
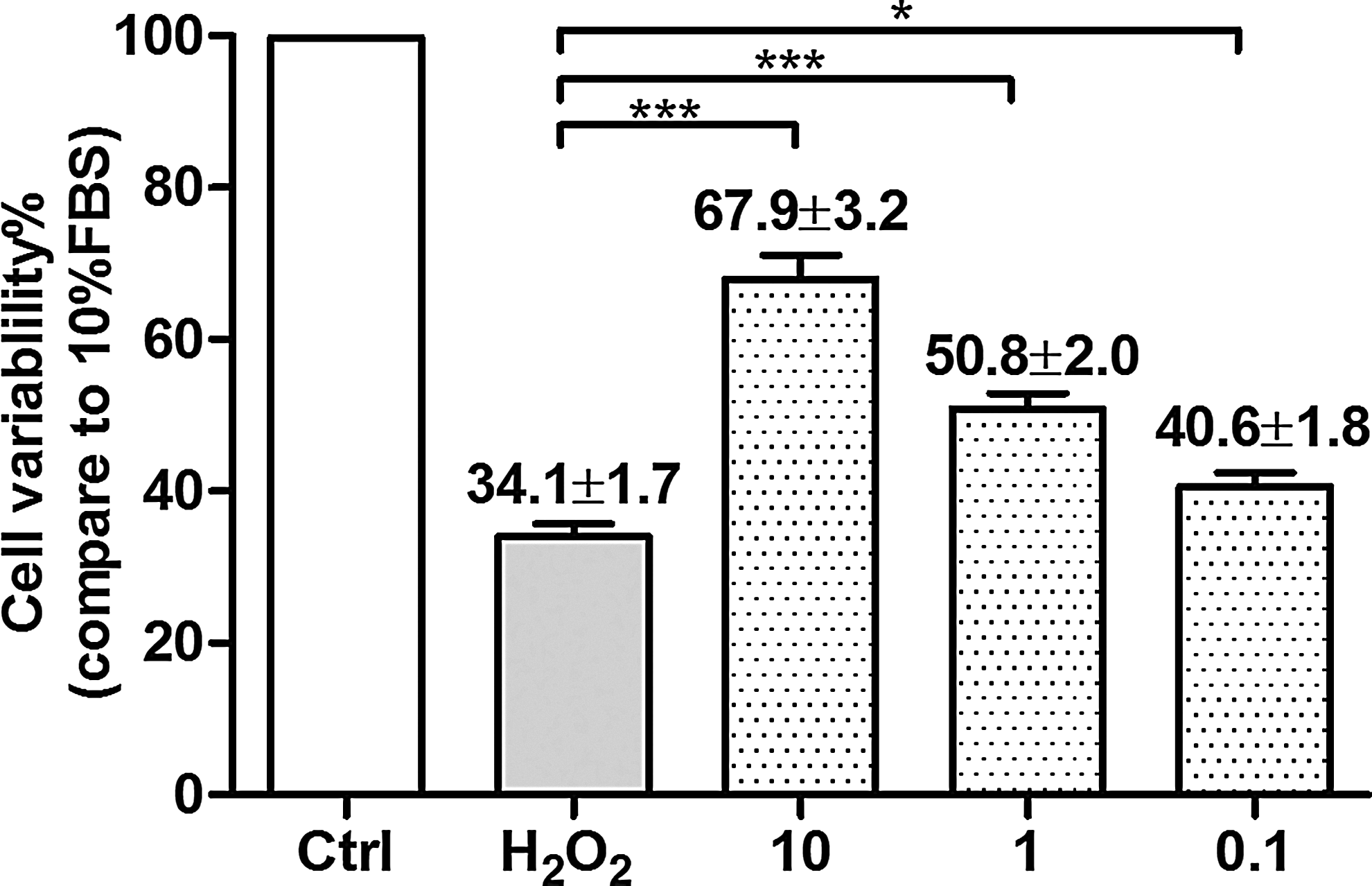
Curcumin protected BV-2 microglia cells against hydrogen peroxide (H2O2)-induced cytotoxicity. Incubation of BV-2 microglia cells with 200 μM H2O2 for 24 h resulted in a significant cell death with only 34.1% cell live. Curcumin protected BV-2 microglia cells against H2O2-induced cytotoxicity dose dependently by pretreatment with 10, 1, and 0.1 μM curcumin. The x-axis represents different treatment groups, whereas the y-axis is the ratio of cell variability of treatment groups compared with 10% fetal bovine serum (FBS) containing culture medium treatment group (Ctrl). Each experiment was repeated at least 3 separate times. The data were presented as mean±SD. *P<0.05, ***P<0.001.
Curcumin inhibited H2O2-induced increase in BV-2 microglia intracellular ROS
ROS are generated as by-products of cellular metabolism. When cellular production of ROS overwhelms its antioxidant capacity, damage to cellular macromolecules may ensue. To evaluate the antioxidant capacity of curcumin, flow cytometry analysis was used to evaluate the intracellular ROS. Our results showed that curcumin pretreatment dose dependently (10, 1, and 0.1 μM) reduced the ROS production in BV-2 microglia cells compared with H2O2-treated controls (P<0.05, Fig. 2).
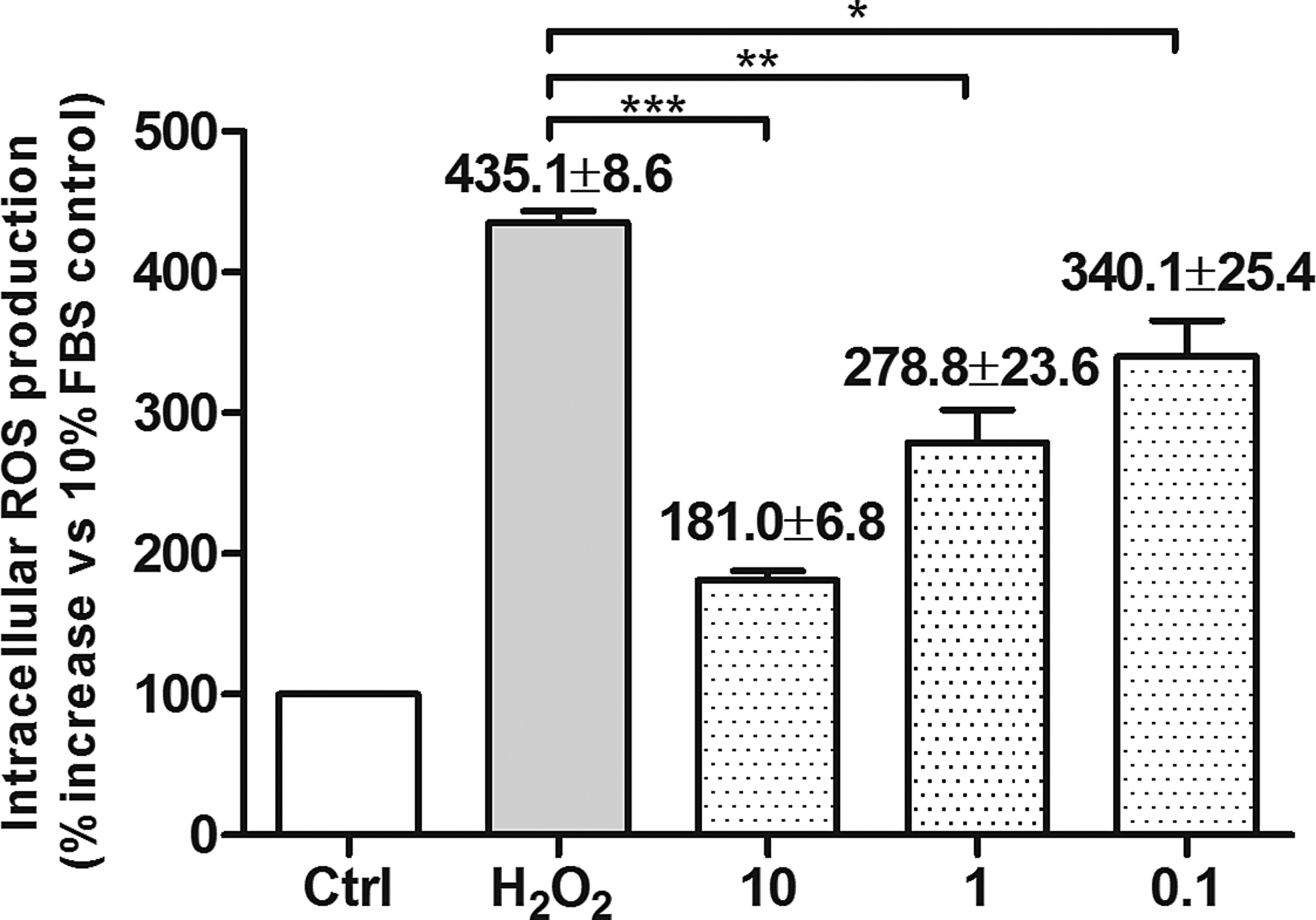
Curcumin inhibited H2O2-induced increase in BV-2 microglia intracellular reactive oxygen species (ROS). Flow cytometry analysis illustrated that 200 μM H2O2 induce the release of ROS significantly. Curcumin pretreatment dose dependently reduced the ROS production in BV-2 microglia cells. Each experiment was repeated at least 3 separate times. The x-axis represents different treatment groups, whereas the y-axis is the ratio of ROS production in different treatment groups compared with 10% FBS treatment group (Ctrl) (by percentage). The data were presented as mean±SD. *P<0.05, **P<0.01, ***P<0.001.
Curcumin protected against H2O2-induced apoptosis
H2O2 is known to induce cell apoptosis in vitro. To evaluate the effects of curcumin in early and late apoptosis, flow cytometry was used in our study. Collected cells were divided into 4 groups: dead cells (Annexin V−/PI+, upper left, UL), late apoptotic cells (Annexin V+/PI+, upper right, UR), viable cells (Annexin V−/PI−, lower left, LL), and early apoptotic cells (Annexin V+/PI−, lower right, LR). The apoptotic rate was calculated as the percentage of early apoptotic cells (LR) plus late apoptotic cells (UR). Pretreatment with curcumin dose dependently inhibited apoptosis, and the percentage of apoptotic cells (UR+LR%) was significantly lower in H2O2-treated cells compared with untreated cells at 3, 6, 12, and 24 h after incubation (P<0.05, Fig. 3).
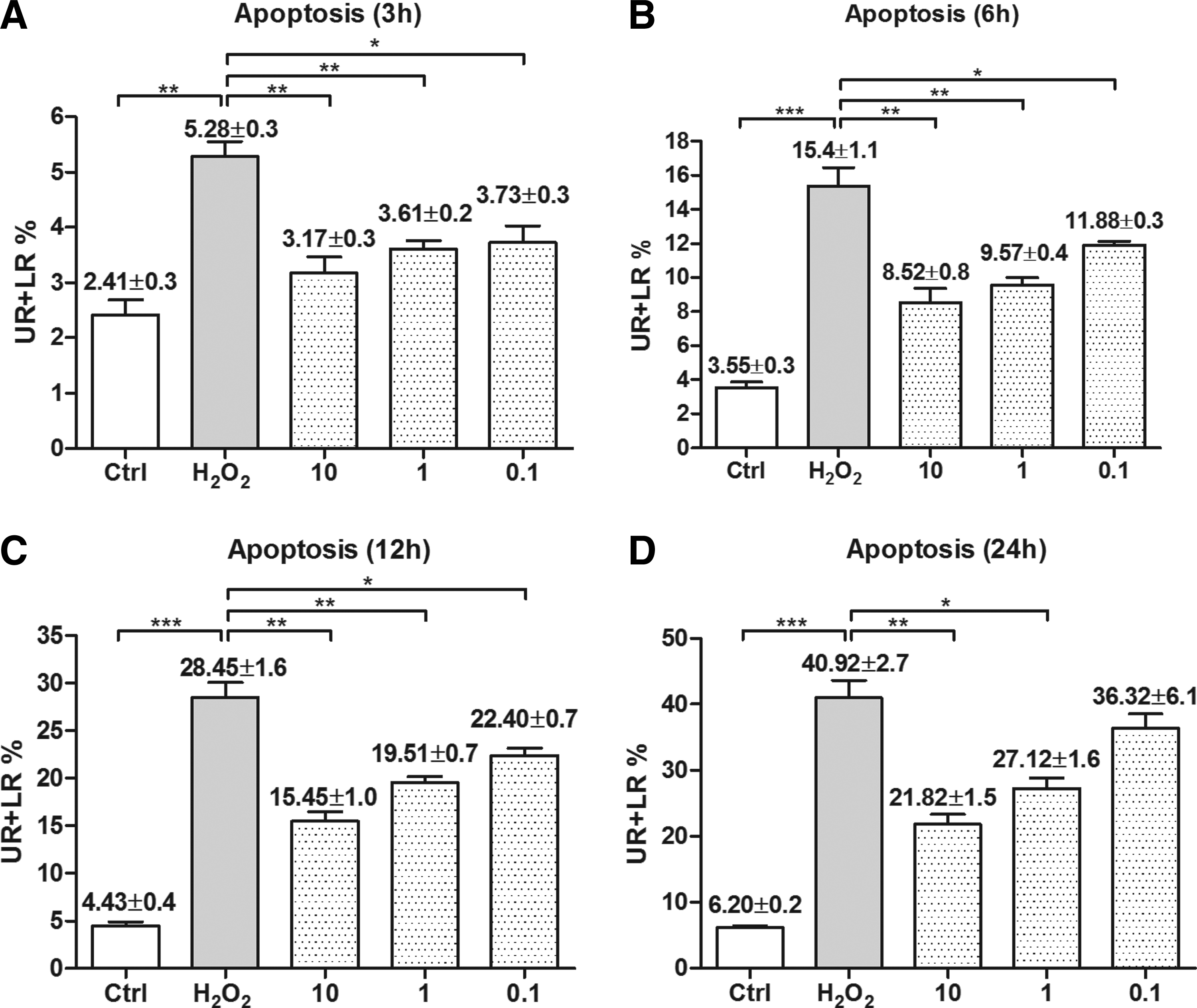
Curcumin protected against H2O2-induced apoptosis. Flow cytometry analysis was performed at the time point of 3 h
Curcumin inhibited H2O2- and high IOP-induced caspase 3, cyto c, and BAX activation while increased BCL2 activation in BV-2 microglia and animal model
Caspase 3, cyto c, BAX, and BCL2 activation is related to apoptosis and mitochondria oxidative damage, which are important indicators for cell damage. In this study, Western blot analysis was performed to evaluate the expression of these proteins in both H2O2- and high IOP-induced oxidative damage in BV-2 microglia and glaucoma models. Compared with controls, caspase 3, cyto c, and BAX were upregulated. The treatment of curcumin could inhibit the activation of these apoptosis-related proteins (P<0.05, Figs. 4 and 5), whereas the treatment of curcumin can upregulate the antiapoptotic factor BCL2 in both BV-2 microglia and animal models.
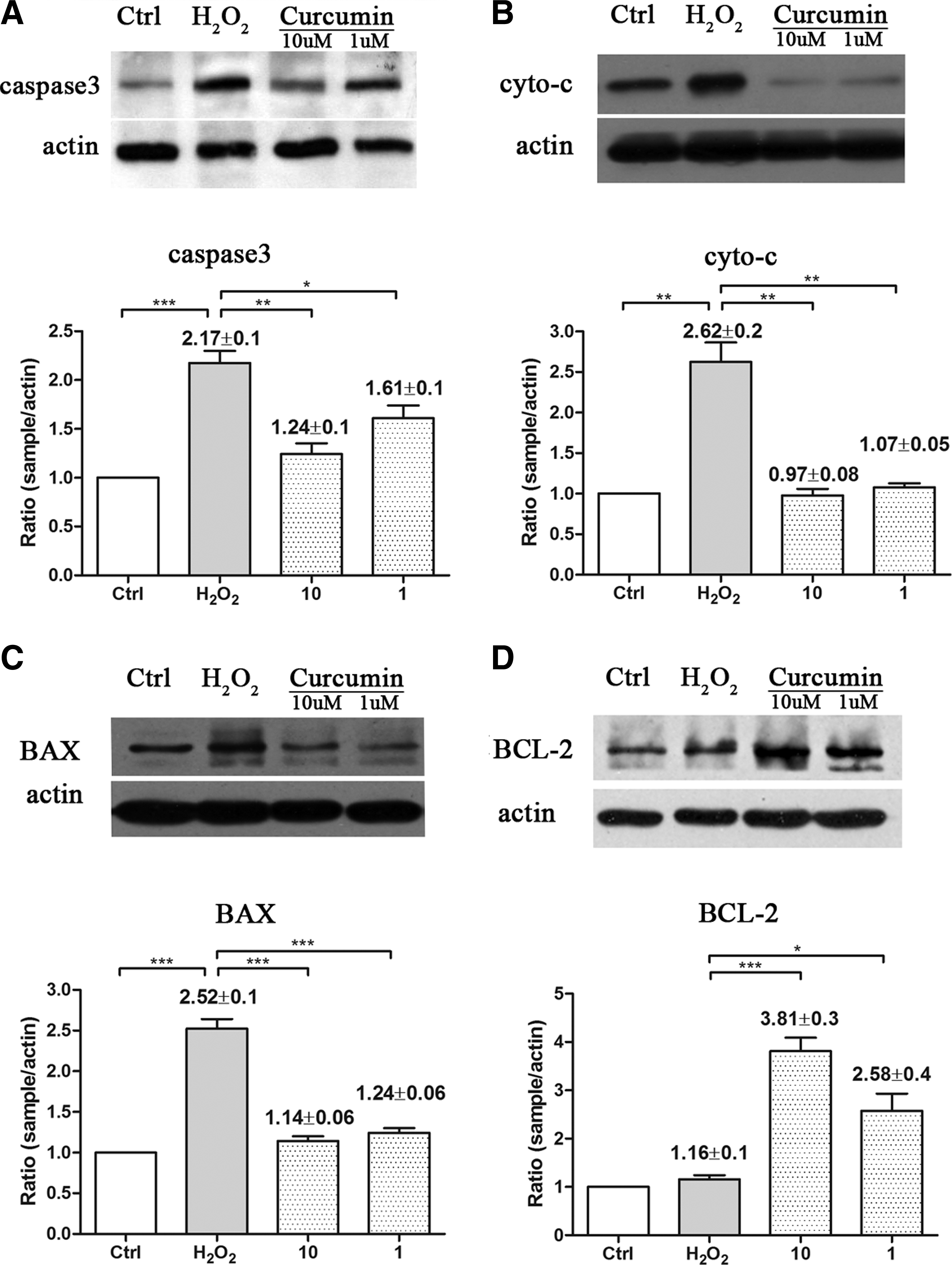
Curcumin inhibited H2O2-induced proapoptotic factors caspase 3, cyto c, and BAX activation and increased antiapoptotic factor BCL2 activation.
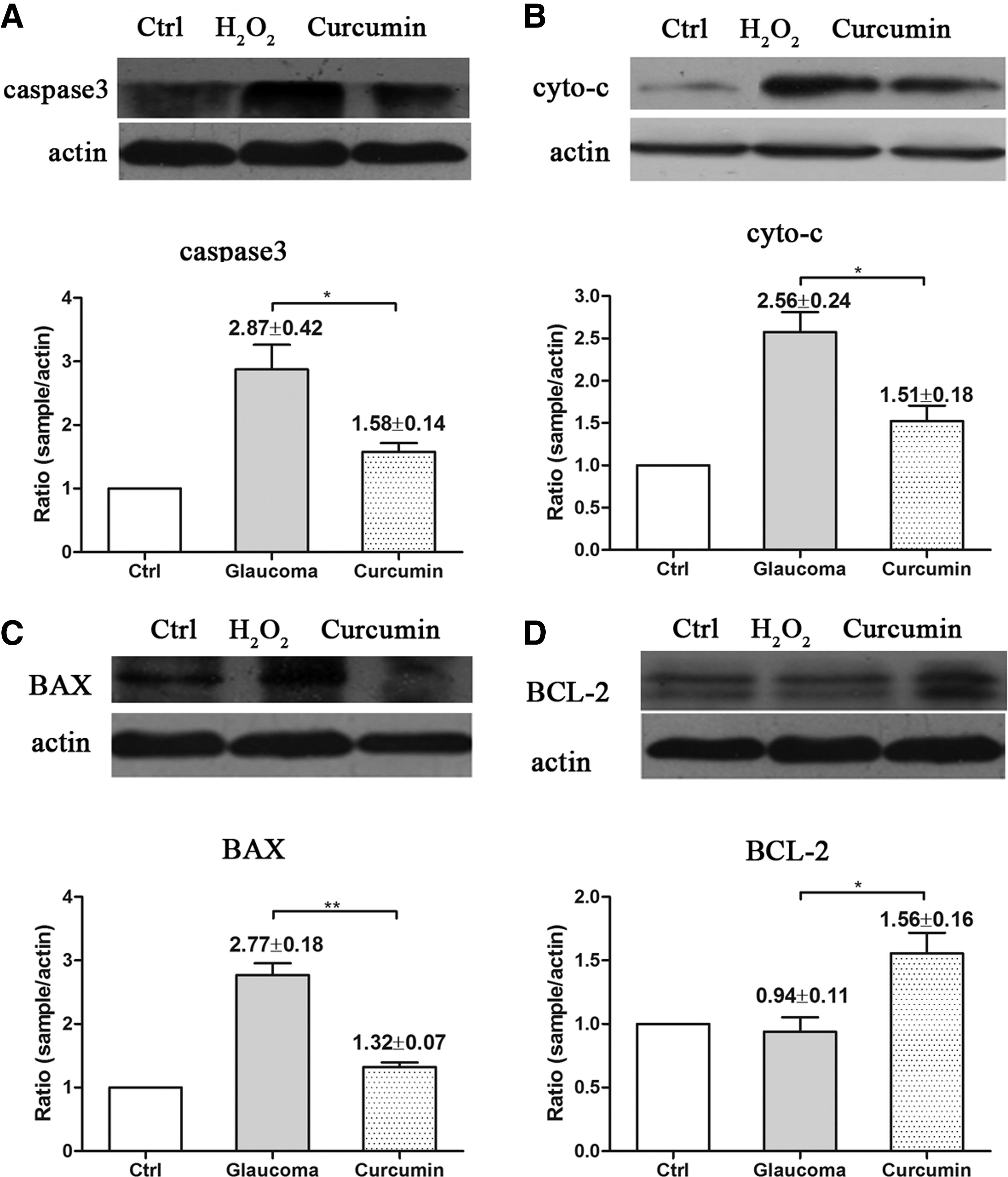
Curcumin inhibited high intraocular pressure (IOP)-induced proapoptotic factors caspase 3, cyto c, and BAX activation and increased antiapoptotic factor BCL2 activation.
Curcumin protected RGCs from death in high IOP rat model
To evaluate the effects of curcumin in vivo, high IOP rat model was used in this study. Cauterization of 3 episcleral veins in Wistar rats can produce sustained elevation of IOP and consecutive loss of RGCs during the 6 weeks of treatment (Fig. 6A, B). The RGCs of the rats fed with curcumin survived longer than RGCs of the untreated group. The untreated and PBS-treated high IOP rat model showed RGCs death rate of ∼26%, and curcumin-treated rats (10 mg/kg/day) of about 19%, which is significantly different from the control group (curcumin-treated group vs. the PBS and untreated group, P<0.05, Fig. 6B, C).

Effects of curcumin in high IOP rat model.
Discussion
In the present study, we examined the effect of curcumin against H2O2-induced sustained oxidative stress in BV-2 microglia cells. Our results showed that pretreatment with curcumin could prevent oxidative damage measured by MTT, decrease intracellular ROS level, and alleviate apoptosis induced by oxidative stress. In addition, in a high IOP animal model, curcumin can prevent the death of RGCs better than untreated controls. In both in vitro and in vivo experiments, curcumin inhibited proapoptotic factors, such as caspase 3, cyto c, and BAX, whereas upregulated the antiapoptotic factor BCL2. All of the evidence points to the possibility of the use of curcumin for treatment in the future.
Glaucoma is characterized by the elevation of IOP, and increased IOP, vascular damage, and hypoxia are also often associated with glaucoma.22,23 Insufficient autoregulation increases the risk of unstable ocular perfusion, leading to an unstable oxygen supply.24,25 This unstable oxygen supply generates oxidative stress. Growing evidence suggests that oxidative stress is a common component of glaucomatous neurodegeneration. 26 ROS are generated as by-products of cellular metabolism, primarily in the mitochondria. Overexpressed ROS can damage cellular macromolecules, such as DNA, proteins, and lipids.27,28 Such a state of oxidative stress is thought to contribute to the pathogenesis of several neurodegenerative diseases, including high tension and normal tension glaucoma. 25 Besides the direct damage to RGCs by ROS, they also lead to glial dysfunction. 3 Oxidative stress-induced dysfunction of glial cells may contribute to spreading of neuronal damage by secondary degeneration. 29 In addition, oxidative stress plays a part in the activation of the immune response during glaucomatous neurodegeneration, which can accelerate the degeneration of the retina in glaucoma patients. 29 As a consequence of oxidative stress, the concentration of superoxide (O2−) within the axons of the optic nerve head increases. The activation of the neighboring astrocytes by mechanical or by ischemic stress induces excessive production of nitric oxide (NO), which diffuses into the axons and fuses with superoxide. 30 The resulting peroxynitrate diffuses within the axons toward the retina and the lateral geniculate nucleus and induces apoptosis. Therefore, antioxidant treatment in glaucoma patients is important, and many studies have demonstrated its benefit.
H2O2 is a key oxidant, widely used to induce oxidative stress in experimental studies, which can cause ROS generation and apoptosis. 17 It has been reported that the retinas of primary open-angle glaucoma patients were exposed to a high concentration of ROS compared with normal controls. Therefore, we used H2O2 to induce chronic oxidative stress in BV-2 microglia in our in vitro study. To avoid possible physical interaction between curcumin and H2O2, the cells were first pretreated with curcumin and then incubated with H2O2. As stated above, in both in vitro and in vivo animal studies, curcumin has shown antitumor, antioxidant, antiarthritic, antiamyloid, anti-inflammatory, and anti-ischemic properties. 8 In our study, we also showed the antioxidant property in BV-2 microglia under H2O2 stimulation. Although the detailed mechanism of curcumin remains unclear, we propose that it is involved in repairing the function of mitochondria in BV-2 microglia cells (Figs. 1–5).
In addition to our own findings, another study found that curcumin acts as a free radical scavenger and antioxidant, inhibiting lipid peroxidation and oxidative DNA damage, protecting against lead neurotoxicity. 31 Numerous studies have also demonstrated that curcumin has a positive effect on neurogenesis in the hippocampus and increases the levels of brain-derived neurotrophic factor in rats, which is important for the glaucoma mechanism.20,32 At the same time, Ravindran et al. proved that curcumin acts on cancer by modulating the growth of tumor cells through the regulation of multiple cell signaling pathways, including the cell proliferation pathway (cyclin D1, c-myc), cell survival pathway (Bcl-2, Bcl-xL, cFLIP, XIAP, c-IAP1), caspase activation pathway (caspase 8, 3, 9), tumor suppressor pathway (p53, p21), death receptor pathway (DR4, DR5), mitochondrial pathway, and protein kinase pathway (JNK, Akt, AMPK). 33 Although curcumin was effective in a broad-spectrum medication field, it has had poor systemic availability, but recent studies have shown that the phosphatidylcholine formulation (Meriva®) increases its oral bioavailability.34,35
In conclusion, in the present study, curcumin showed a significant antioxidant and antiapoptotic effects on BV-2 microglia cells and can inhibit the death of RGCs in a high intraocular animal model. Curcumin could be a new treatment or adjunctive method for treating glaucoma, but the detailed mechanism needs to be explored further.
Footnotes
Acknowledgment
This study was supported by Research Fund of the National Natural Science Foundation of China (No. 81070735).
Author Disclosure Statement
No competing financial interests exist.