
Abstract
Abstract
The structural and molecular properties of the human tonsil lymphatic microvascular system are important to understand as these features likely contribute to fluid balance, immunity, and tumor metastasis. The tonsil is a unique lymphoid organ in that it is in intimate contact with the contents of the upper aerodigestive tract and that there are no identifiable afferent lymphatics. Conventional immunofluorescence microscopy demonstrated a remarkable degree of lymphatic vessel architecture within the tonsil; LYVE-1-positive lymphatic vessels were detected around each germinal center and in the marginal regions between the follicles. High resolution confocal laser scanning immunofluoresence microscopy demonstrated that individual lymphatic endothelial cells had a classic ‘oak leaf’ shape and discontinuous expression of CD31 and VE-cadherin; characteristics hypothesized to be related to fluid and cellular transport. A comparative analysis demonstrated a dramatic increase in lymphatic but not blood vessel density and complexity in inflamed compared to noninflamed tonsil tissue. The results of this study describe the spatial organization of the tonsil lymphatic vasculature, discontinuous expression of CD31 and VE-cadherin in human lymphatic capillaries, and a change in lymphatic vessel morphology in response to inflammation.
Introduction
The association between inflammatory and malignant diseases and the lymphatic vasculature system has generated an interest in understanding the structure and function of the lymphatic system. Although it has not been demonstrated in clinical studies, the results of several preclinical models have demonstrated that the lymphatic system grows and remodels, a process termed lymphangiogenesis, in response to at least certain types of inflammatory stimuli,4,5
In the last 10 years, antibodies reagents and microscopy techniques have been developed to detect the lymphatic vasculature in preclinical models. For example, rabbit antibodies to the hyaluronic acid receptor, LYVE-1, have been shown to detect mouse or human lymphatic vessels reliably.6,7 The adhesion molecule PECAM-1 (CD31) is expressed on lymphatic and more significantly on vascular endothelial cells at the junctions between adjacent cells. Immunoreactivity with antibodies to LYVE-1 or CD31 and conventional immunofluorescence (IF) or confocal laser scanning microscopy (LSM) techniques were used to identified the lymphatic or blood vasculature, respectively, in human pediatric tonsil tissue. Remarkably conserved lymphatic vessel architecture was observed in noninflamed tonsil tissue. Lymphatic capillaries surrounded but did not extend into germinal centers.
Confocal LSM and image analysis acquires serial images in the ‘z’ plane that are stacked to create a three-dimensional reconstruction useful for evaluating vessel morphology and cell surface molecules that have spatially restricted expression. Discontinuous expression of the adhesion molecules CD31 and VE-cadherin is a phenotype of initial lymphatic capillaries.8,9 This expression pattern is hypothesized to facilitate fluid and cellular transport in lymphatic endothelial cells. This phenotype was detected in some of lymphatic vessels, suggesting the presence of initial lymphatic capillaries in human tonsil tissue. A dramatic increase in lymphatic but not blood vessel density and complexity was identified in inflamed tonsil tissue. The results of this study describe the spatial organization of the human tonsil lymphatic system, the discontinuous expression of CD31 and VE-cadherin in human lymphatic capillaries, and a remarkable change in lymphatic morphology in inflamed tonsil tissue.
Materials and Methods
Subjects
The clinical research described in this application was approved by the Boys Town National Research Hospital Institutional Review Board. Patients were recruited from the clinical practice of the senior author. Witnessed parent consent was obtained. Tonsil tissue was collected from patients 2–8 years old. Two groups were identified. The first group, identified as inflamed, had clinical signs of tonsil inflammation (erythema and exudate) at the time of tonsillectomy and a history of recurrent tonsillitis (n = 6). The second group, identified as noninflamed, was without clinical signs of tonsil inflammation and did not have a history of tonsillitis. These patients had tonsil hypertrophy and a diagnosis of sleep disordered breathing (n = 12). Categorization (inflamed or noninflamed) was made at the time of tonsillectomy by senior author. Tonsillectomy was performed using the established standard technique of pencil electrocautery.
Conventional immunofluorescence
Tonsil tissue was frozen in optimum cutting temperature (OCT) compound and 8 micron sections were cut with a cryostat and fixed with cold acetone. Samples were washed with phosphate buffer saline (PBS) pH 7.4. Nonspecific antibody reactivity was blocked by incubating the tissue with 10% fetal calf serum (FCS) in PBS, pH 7.4. Rabbit antibodies to LYVE-1 (Millipore, Temecula, CA) or purified negative control rabbit sera (Santa Cruz Biotechnology, Santa Cruz, CA) were used at 1/500 to detect the lymphatic vasculature. Mouse antibodies to human CD31 (BD Bioscience, San Jose, CA) or negative control mouse IgG (Invitrogen, Carlsbad, CA) were used at 1/300 to detect the blood vasculature. Fluorescently-labeled species specific secondary antibodies (1/1000) were used to detect the primary antibodies. The data was collected using the Zeiss Axioimager microscope and images were produced with Spot software (Diagnostic Instruments Inc., Sterling Heights, MI).
Tissue processing, antibody staining, and confocal LSM image acquisition
100–140 micron sections of the superior, inferior, and mid-portion of each freshly obtained tonsil specimen was fixed with 1% paraformaldehyde (PFA) for 1–2 h and permeabilized with PBS solution containing 0.3% triton-X, 0.2% FCS, and 0.02% sodium azide (PBS+). All antibody incubations were performed using PBS+ with 5% FCS. Washing buffer contained PBS with 0.3 % triton-X, 2% bovine serum albumin, and 0.02% sodium azide. Antibodies to LYVE-1 (Millipore, Temecula, CA) or purified negative control rabbit sera (Hyclone, Logan, UT) were used at 1/250 to detect the lymphatic vasculature. Antibodies to CD31 (BD Bioscience), VE-cadherin (BD Bioscience) or negative control mouse IgG1 (Invitrogen, Carlsbad, CA) were used at 1/50. Samples were washed with washing buffer three times. After washing, the appropriate secondary antibodies directly conjugated to fluorochromes (1/500) were used to detect the primary antibodies. The data were collected on a Zeiss confocal LSM 510 and the images were produced using Zeiss AimImageBrowser.
Statistical analysis
Representative microscopic fields of the superior, middle, and inferior portions of each tonsil were recorded. A Matlab application was designed to provide a quantitative assessment of lymphatic and blood microvascular density by counting the number of ‘red (lymphatic vascular)’ or ‘green (blood vascular)’ pixels of intensity greater than the threshold established by visual inspection. The percentage of red or green pixels for each tonsil image was calculated. The statistical difference between the two groups was calculated using Student's t test.
Results
Germinal centers and marginal or stromal regions are well visualized in human tonsil tissue stained with standard hematoxylin and eosin techniques (Fig. 1A). The lymphatic and blood vasculature was detected in noninflamed tonsil tissue using IF microscopy. Staining with antibodies to CD31 (Fig. 1B) or LYVE-1 (Fig. 1C) identified the blood or lymphatic vasculature, respectively, within the marginal zone of the tonsil and surrounding the germinal center.
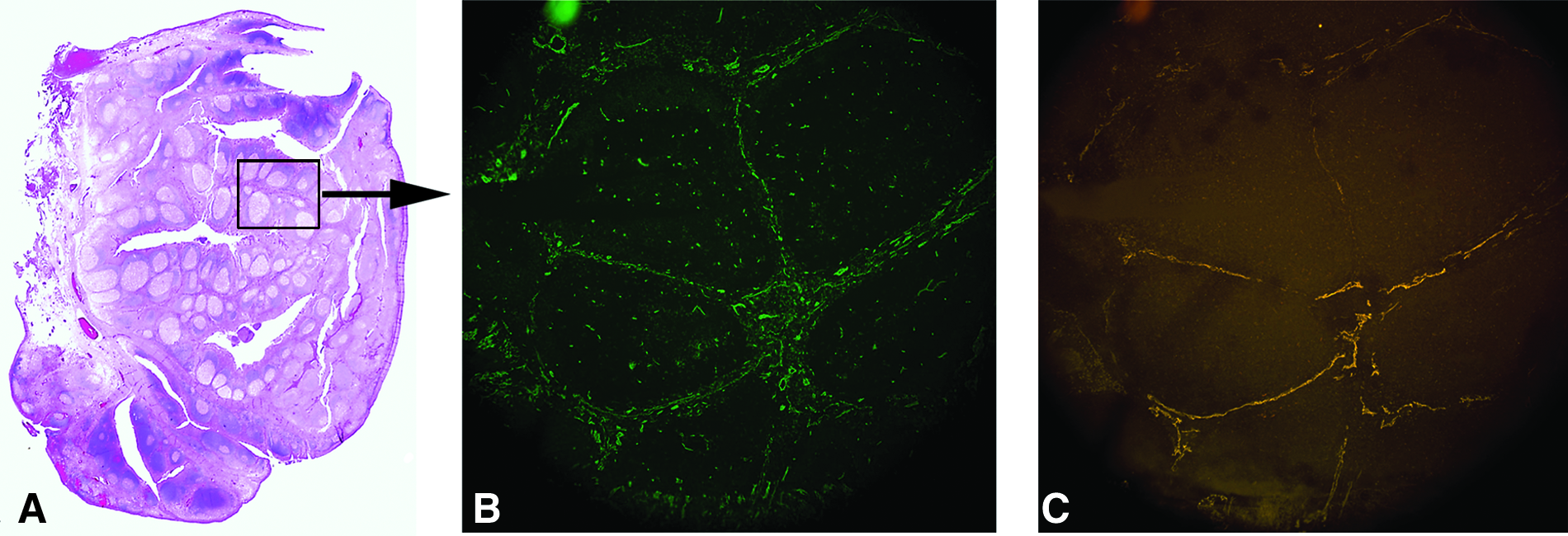
(
Confocal LSM and image analysis was used to visualize the complex morphology of the blood and lymphatic vasculature within the noninflamed tonsil tissue. Lymphatic vessels identified by LYVE-1 staining in noninflamed tonsil tissue were abundant, irregularly shaped, and in many cases contoured to the edge of the germinal center (Fig. 2). Blood vessels were detected in the marginal regions, and thin CD31-positive LYVE-1 negative vessels-like structures were detected within the germinal centers. They are presumably endothelial extensions of blood vessels and their significance is unknown. High magnification demonstrated the expression of LYVE-1 at the perimeter of the lymphatic endothelial cells outlining an ‘oak leaf’ pattern (Figs. 3A and 3B). 10 Discontinuous expression of CD31 (Fig. 3A) and VE-cadherin (Fig. 3B) was identified in both marginal lymphatics and in lymphatic vessels adjacent to the germinal center, suggesting the presence of initial lymphatic capillaries in noninflamed tonsil tissue. A restricted spatial pattern of CD31 or VE-cadherin expression on lymphatic vessels or within the tonsil could not be identified.
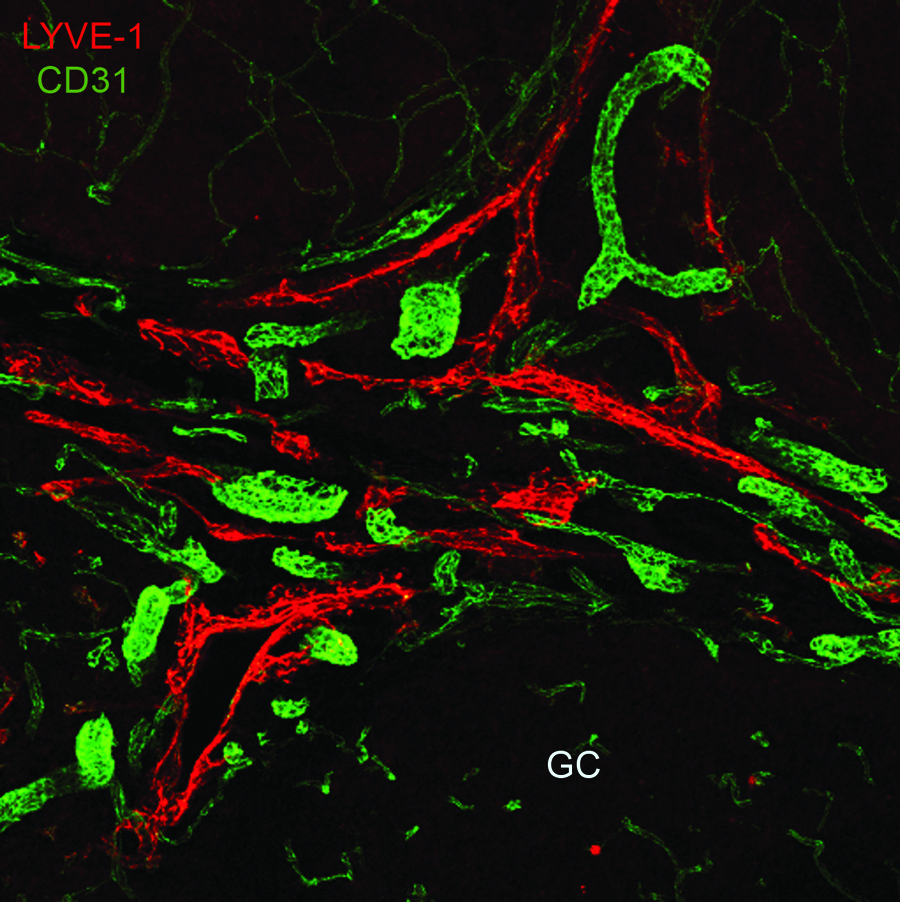
Lymphatic vessels in noninflamed tonsil tissue are relatively large, irregular, and surround the germinal center (GC) periphery. 100 μm noninflamed tonsil specimens were stained with antibodies to LYVE-1 or CD31 to detect the lymphatic or blood vasculature system, respectively. Confocal LSM and image analysis was used to acquire the data. Isotype specific control samples generated no specific signal (data not shown). (See Lymphatic Research and Biology at www.liebertonline.com for color figure.)
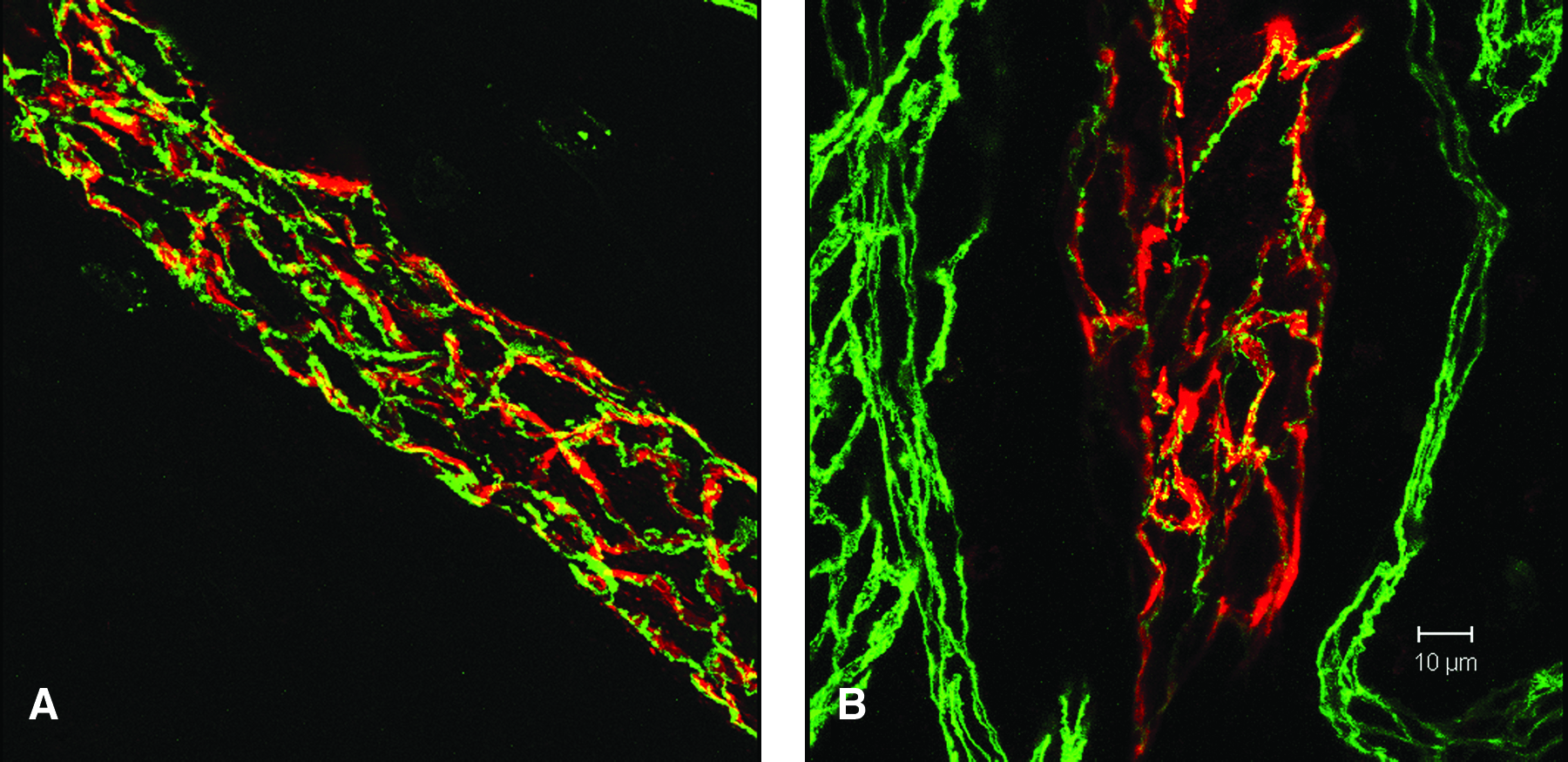
Discontinuous expression of CD31 and VE-cadherin in LYVE-1 positive lymphatic vessels. 100 μm noninflamed tonsil specimens were stained with antibodies to LYVE-1 (red) and co-stained with antibodies to CD31 (green;
A qualitative increase in lymphatic vessels in inflamed compared to noninflamed tonsil tissue was detected by IF microscopy (Figs. 4A–4D). This analysis revealed a remarkable degree of stromal hypertrophy in the inflamed compared to noninflamed tonsil tissue. Confocal LSM and image analysis demonstrated a similar qualitative increase in lymphatic density and complexity in inflamed (Figs. 5A–5C) compared to noninflamed (Figs. 5D–5F) tonsil tissue. Quantitative analysis using Matlab software demonstrated a statistically significant increase in lymphatic microvascular density in inflamed tonsil tissue compared to noninflamed tissue without inflammation (p = 0.03). A similar analysis demonstrated no significant difference in the blood microvascular density in inflamed tonsil tissue compared to noninflamed tonsil tissue without inflammation (Fig. 6).
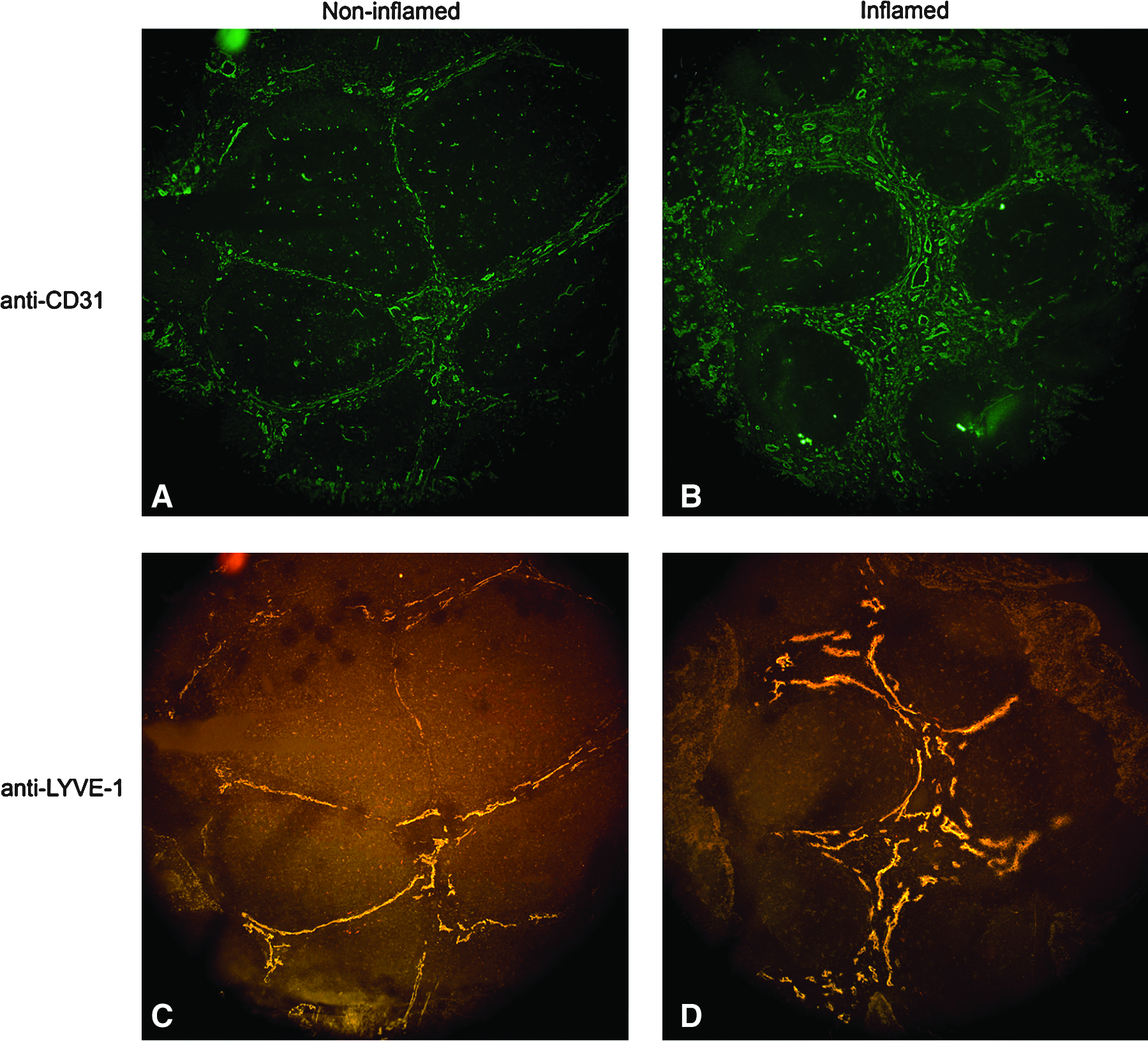
Lymphatic hypertrophy within the marginal zone of inflamed tonsil tissue. 8 micron noninflamed (
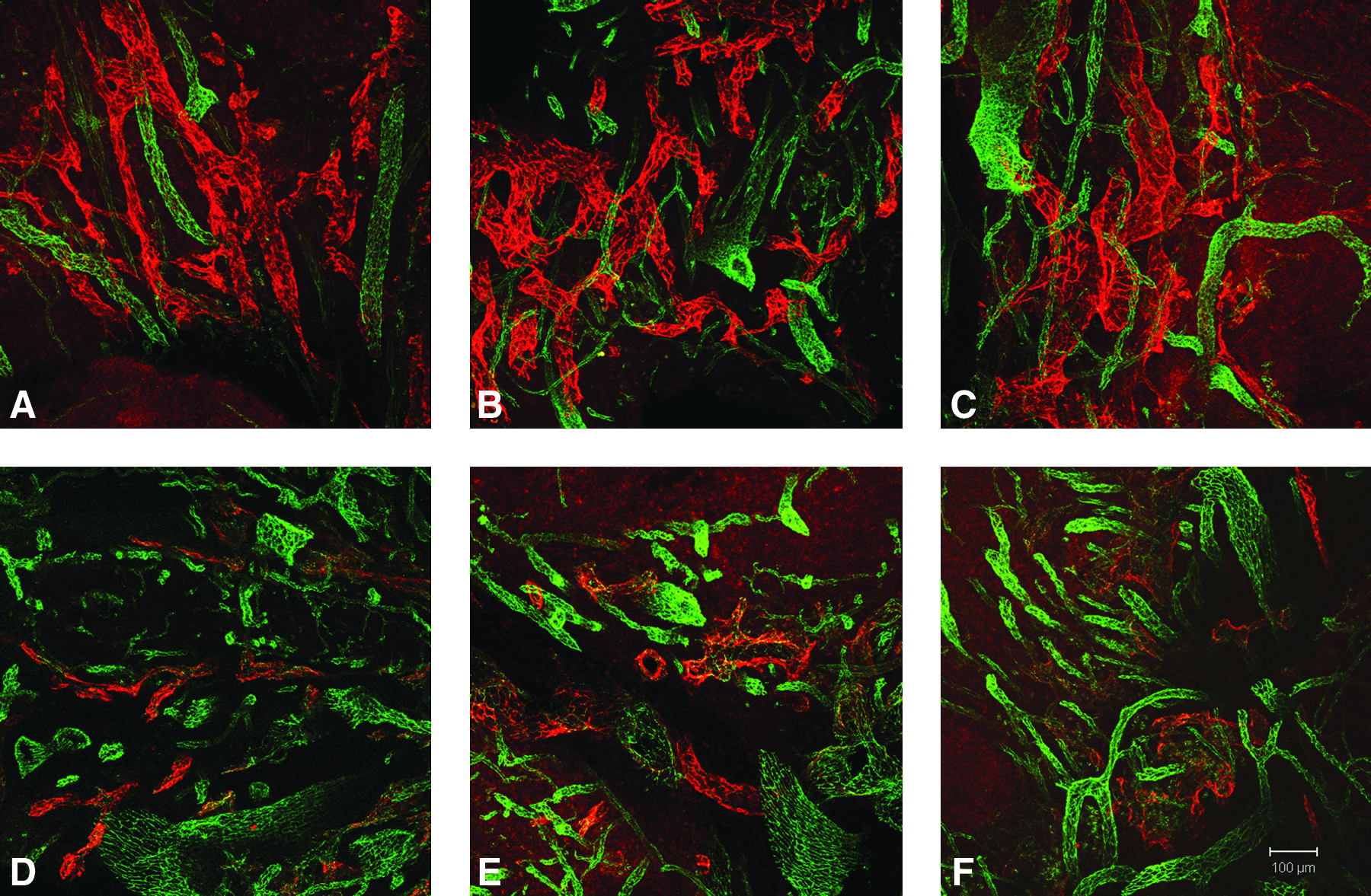
Lymphatic hypertrophy in inflamed tonsil tissue. 100 μm specimens were obtained from inflamed (
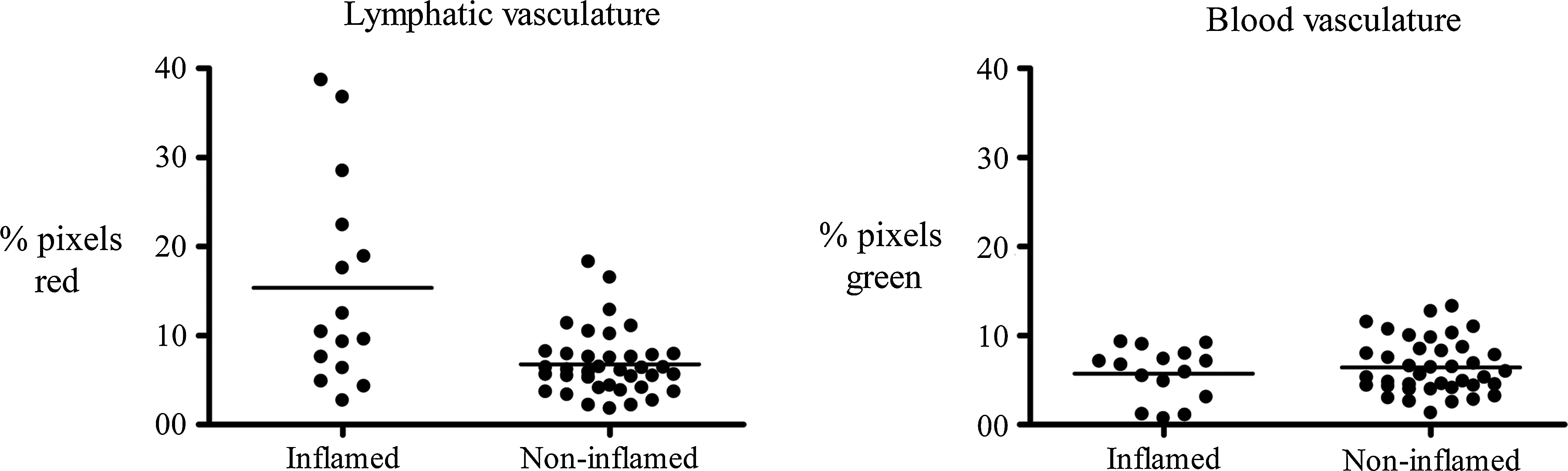
A statistically significant difference was detected in lymphatic but not blood vasculature density in inflamed tonsil tissue. The percentage of red (lymphatic) or green (blood) pixels for each tonsil image was calculated and is shown. The statistical difference between the two groups was calculated using Student's t test.
Conclusions
The results of this study demonstrate that the lymphatic vasculature is abundant and well organized within the marginal regions of the tonsil tissue but not within the germinal centers. The function of the tonsil is not known with certainty; however, it is suspected to contribute to immunity. The tonsil is one of the first lymphoid aggregates to encounter pathogens that enter the oral cavity and it is composed of leukocytes organized in germinal centers and stromal tissue. Interestingly, the tonsils do not have afferent lymphatics, a feature that suggests that tonsil lymph is generated intrinsically. The tonsil anatomy may be conceptualized as distal or terminal lymphatic capillaries positioned in intimate contact with lymphoid tissue. This is dissimilar to the conventional schemata of afferent lymphatic vessels draining into lymphoid aggregates (lymph nodes). The discontinuous expression of CD31 and VE-cadherin in human tonsil lymphatic capillaries suggested the presence of initial lymphatics capillaries, vessels hypothesized to have attributes of fluid and solute transport.8–10 Efferent tonsil lymph may generated intrinsically by passage of fluid, antigen, or cells conditioned by the constituents of the germinal centers into the initial lymphatic capillaries within the tonsil. This unique anatomic relationship could contribute to local and peripheral immunity. 11
Preclinical corneal and tracheal models have demonstrated lymphangiogenesis in response to inflammatory stimuli.4,5 Inflammation-mediated lymphangiogenesis is hypothesized to occur clinically; although this principle has not been tested in human subjects. Lymphatic morphology and density was increased in inflamed pediatric tonsil tissue compared to noninflamed tonsil tissue. IF microscopy and confocal LSM and image analysis techniques demonstrated a qualitative increase in lymphatic density in inflamed compared to noninflamed tissue. These results were consistent with a quantitative analysis that demonstrated a statistically significant increase in lymphatic but not blood vasculature in inflamed compared to noninflamed tonsil tissue. Additionally, remarkable stromal hypertrophy was detected, a common event in progressive inflammatory conditions. The findings reported here are consistent with the proposed model based on the results of preclinical studies demonstrating that certain inflammatory conditions stimulate lymphangiogenesis. Interestingly, no detectable increase in blood vascular density was identified in inflamed tonsil tissue. This was unexpected, as in many preclinical experimental systems angiogenesis occurs in concert with lymphangiogenesis.4,5 Inflammation-mediated lymphangiogenesis but not angiogenesis may be unique to the tonsil microenvironment or the pathogenesis of tonsillitis which in practice and in this study is often not identified specifically. Important questions to address are 1) whether lymphangiogenesis is common to all types of inflammatory responses or represents a subset of inflammatory processes; and 2) how lymphangiogenesis alters host immunity and the course of clinical disease.
One potential limitation of this study is subject group allocation and the disease state that is represented in each group. Nonhypertrophic noninflamed pediatric tonsil tissue is difficult to obtain, and this study does not include such a group. This study was designed such that tonsil inflammation (erythema and exudate) was identified at the time of tonsillectomy in individuals with a history of tonsillitis. A correlation between Streptococcal infection or a history of Streptococcal infection could not be made reliably. Noninflamed hypertrophic tonsils were identified by the absence of erythema, and mucopurulence. These patients had obstructive symptoms, for instance snoring, sleep apnea, and potentially a history of hypoxia. It is recognized that the etiology of tonsil hypertrophy is poorly understood and potentially could involve inflammation. Collectively, these issues may explain the variance in our statistical analysis.
The results of these studies establish useful microscopy techniques to visualize the human lymphatic microvascular system in lymphoid tissue. A highly organized lymphatic vasculature comprised in part of initial lymphatic capillaries was demonstrated to increase in complexity in inflamed tonsil tissue. These clinical findings are a prerequisite to understanding how the lymphatic vasculature interfaces with lymphoid tissue at the cellular and molecular level. The effects of lymphangiogenesis on the extracellular matrix, immunity, and end organ function are important unresolved questions. Understanding these principles may provide important insight towards the mechanism of progressive inflammatory disease.
Footnotes
Acknowledgments
The authors thank the patients and their families that participated in this study, Denis Fitzpatrick for his assistance developing the Matlab software, and Dominic Cosgrove for his critical review of this manuscript.
Author Disclosure Statement
No competing financial interests exist.
The project was funded by State of Nebraska Tobacco Research Initiative.