
Abstract
The human leukocyte-associated Ig-like receptor (LAIR) family contains two members: LAIR-1 (CD305) and LAIR-2 (CD306). Among them, LAIR-1 is a transmembrane glycoprotein bearing two intracellular immunoreceptor tyrosine-based inhibition motifs (ITIM) and LAIR-2 is soluble. Both molecules bind collagen and LAIR-2 has higher affinity than LAIR-1. LAIR-1 can mediate strong inhibitory signal but the functions of leukocytes expressing LAIR-1 are unclear because of the absence of an effective method to isolate them with resting status. In this study, we generated a monoclonal antibody (MAb) by immunizing BALB/c mice with the recombinant LAIR-2-GST fusion protein, which we termed 3G4. The subclass of 3G4 was identified as IgG1. Specificity analysis by Western blotting demonstrated 3G4 could react with both LAIR-1 and LAIR-2. Unlike another LAIR-1-specific MAb (9.1C3), 3G4 did not inhibit the lysis of target cells P815 by NK cells in a redirected cytotoxicity assay. Preincubation of LAIR-1-transfected K562 cells with 3G4 mildly prevented the binding of LAIR-1 to collagens I and III in a dose-dependent manner. Taken together, the novel MAb 3G4 provides a useful tool to isolate LAIR-1-positive cells without changing their resting state for further application.
Introduction
A
As an ITIM-bearing transmembrane molecule, LAIR-1 functions as an inhibitory receptor on many immune cells by recruiting the Src homology 2 (SH2) domain-containing phosphatase (SHP)-1 or SHP-2. In cells lacking both SHP-1 and SHP-2, LAIR-1 can also mediate inhibitory signals by recruiting C-terminal Src kinase (Csk).(12) Collagen is identified as a ligand for LAIR-1,(13) and the arginine 65 residue in LAIR-1 is critical for the functional interaction of LAIR-1 with collagens.(14) LAIR-1 is reported to inhibit NK cell-mediated cytotoxicity,(4) prevent TCR-mediated activation in effector T cells,(15) attenuate BCR-induced calcium mobilization in primary B cells,(16) decrease FcγRII-induced calcium mobilization in U937 cells,(17) down-regulate FcɛR-induced degranulation of RBL-2H3 cells,(18) and inhibit granulocyte-monocyte colony-stimulating factor (GM-CSF)-induced monocyte differentiation into dendritic cells (DCs).(7) Furthermore, in human myeloid leukemia cells, LAIR-1 can prevent GM-CSF-dependent proliferation(19) and induce programmed cell death.(20) By expressing the LAIR-1 ligand collagen, tumor cells also possess the ability to inhibit immune responses.(21) The similar inhibitory functions of LAIR-1 are also observed in mice.(10) Interestingly, LAIR-2 is a secreted protein that shares 84% homologous with LAIR-1 but lacks a transmembrane and intracellular domain.(4,22) Unlike LAIR-1, LAIR-2 is absent in mouse and rat.(10,23,24) In humans, LAIR-2 can act as a more efficient antagonist of the LAIR-1–collagen interaction than soluble LAIR-1(22,25) shedding from immune cells upon cellular activation.(26)
Although the wide expression of LAIR-1 on most immune cells has been well documented, the functions of these cells expressing LAIR-1 are still unclear because an effective method to isolate them with resting status is lacking. In this study, we generated a novel monoclonal antibody (MAb 3G4) against both LAIR-1 and LAIR-2. Unlike other MAbs targeting LAIR-1, administration of 3G4 did not activate the inhibitory immunoreceptor LAIR-1 on NK cells.
Materials and Methods
Cell lines and antibodies
Mouse Sp2/0 myeloma cells, mastocytoma cell line P815, Jurkat cells, Burkitt lymphoma Raji cells, and green monkey kidney COS7 cells were all purchased from American Type Culture Collection (ATCC, Rockville, MD). The human erythroleukemia K562 cells were also purchased from ATCC (Rockville, MD) and stably transfected with human LAIR-1, as described previously.(13) Mouse anti-LAIR-1 MAb (9.1C3) was purchased from Abcam (Cambridge, MA). Mouse anti-GST MAb (E43.1), anti-C-myc MAb (9E10), and anti-SEB MAb (B4) were generated previously in our laboratory.
Molecular cloning of LAIR-2 cDNA
Total RNA was isolated from peripheral blood mononuclear cells (PBMCs) of a patient with hemorrhagic fever and renal syndrome (HFRS) using a TRizol LS reagent kit (Life Technologies, Carlsbad, CA). The cDNA was obtained by an Advantage RT-for-PCR kit (Clontech, Palo Alto, CA). cDNA encoding LAIR-2 was amplified using the primers (P1: 5′-CAGGAGGGGGCCCTTCCC-3′; P2: 5′-TGGTGCATCAAATCCGGA-3′) and pfu DNA polymerase (Takara, Dalian, China) by PCR. The reaction mixture was subjected to pre-denaturation at 94°C for 5 min followed by 30 amplification cycles each consisting of denaturation at 94°C for 1 min, annealing at 58°C for 30 s and extension at 72°C for 1 min, and a final elongation cycle at 72°C for 10 min that was performed with Taq DNA polymerase (Takara, Dalian, China). Products of PCR were electrophoresed on 10 g/L agarose gels, purified using an Agarose Gel DNA Fragment Recovery kit (Takara), ligated into pMD18-T vector (Takara), and then transformed into Escherichia coli JM109 competent cells for sequencing.
Expression and purification of recombinant LAIR-2-GST and LAIR-2-myc-6×His fusion protein
The reading frame encoding LAIR-2 was amplified using the primers (LAIR-2-forward: 5′-CGGGATCCCAGGAGGGGGCCCTTCCC-3′; LAIR-2-reverse: 5'-CGGAATTCCTGGTGCATCAAATCCGGA-3′) with pfu DNA polymerase and subcloned into a pGEX-4T-3 vector (Amersham Pharmacia Biotech, Piscataway, NJ) by BamHI and EcoRI sites. The corresponding protein was expressed in E. coli BL21 cells as glutathione S-transferase (GST) fusion protein induced by isopropyl-β-D-thiogalactopyranoside (IPTG, Sigma Chemical Co., Saint Louis, MO) and purified with Glutathione Sepharose 4B (Amersham Pharmacia). Similarly, recombinant LAIR-2-myc-6×His fusion protein was expressed in COS7 by eukaryotic expression vector pSecTag2 B (Invitrogen, Carlsbad, CA) and purified with Ni Sepharose (Amersham Pharmacia).
SDS-PAGE and Western blot analysis
To identify the expression of recombinant proteins, SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and Western blot analysis were performed. For SDS-PAGE, the host cells were collected and lysed with ultrasound. Then the lysates with 2× sample loading buffer were resolved on SDS-PAGE under denaturing condition and stained with Coomassie Brilliant Blue R-250 (Sigma). For Western blot analysis, the fractionated proteins on the SDS-PAGE were transferred onto the nitrocellulose membrane (Amersham Pharmacia) in a semi-dry transblot apparatus. The nitrocellulose membrane was blocked with 10% defatted milk in PBS at 37°C for 2 h, and then incubated with the primary antibody for 45 min. Later, the blots were washed twice in PBST (PBS with 0.1% Tween-20) and once in PBS, further incubated with horseradish peroxidase (HRP)-conjugated rabbit anti-mouse IgG for 45 min, followed by detection using ECL reagents (Boehringer Mannheim, Mannheim, Germany), according to the manufacturer's instructions, and exposed to photographic film (Kodak, Rochester, NY).
Hybridoma generation and screening
Monoclonal antibodies against LAIR-2 were generated from LAIR-2-GST-immunized BALB/c mice as described previously.(27) Briefly, female BALB/c mice (4 weeks old) were subcutaneously immunized at a couple of sites in the back with 10 μg purified recombinant LAIR-2-GST fusion protein emulsified in complete Freund's adjuvant (Sigma), followed by two sequential boosts every 3 weeks with the same dosage of LAIR-2-GST fusion protein, the former in incomplete Freund's adjuvant and the latter in normal saline. Ten days later, mice were bled from caudal vein and the serum titers were detected by indirect enzyme-linked immunosorbent assay (ELISA) with purified LAIR-2-myc-6×His. The mouse with the highest antibody titer to LAIR-2-myc-6×His was boosted with 50 μg of LAIR-2-GST by intravenous injection. Three days later, the mouse was sacrificed for spleen harvest and hybridoma generation. The splenocytes were fused with Sp2/0 mouse myeloma cells at a ratio of 5:1 in polyethylene glycol (PEG) 4000 (Merck, Rahway, NJ), according to the general procedure.(28) The fused cells were propagated in 96-well microculture plates (Nunc, Roskilde, Denmark) using hypoxanthine-aminopterin-thymidine (HAT) medium (Sigma). Twelve to 15 days later, culture supernatants were employed to screen the positive hybridomas using the indirect ELISA with LAIR-2-myc-6×His. Selected hybridomas were subcloned three times by limiting dilution prior to ascites production. The MAbs were purified using fast protein liquid chromatography (FPLC) according to previously published protocol,(29) and immunoglobulin (Ig) subclass was identified by using an isotyping kit (Pierce, Rockford, IL). Specificity of MAbs for LAIR-2 was assessed by Western blotting with purified LAIR-2-GST, LAIR-2-myc-6×His, and the chimeric protein composed of the extracellular domain of LAIR-1 fused with human immunoglobulin Fc fragment (LAIR-1-Fc; a gift from Dr. Ouyang,(26) Department of Immunology, Fourth Military Medical University, China).
Isolation of LAIR-1-expression cells by magnetic beads
LAIR-1-positive cells were isolated using immunomagnetic beads with 9.1C3 or 3G4, according to the routine method. Briefly, cells were harvested and washed twice with buffer (30% FCS in PBS) and then resuspended with the buffer (30% FCS in PBS) at a concentration of 1×107cells/mL. After incubation with 9.1C3 MAb for 10 min at 37°C, goat anti-mouse IgG-conjugated magnetic beads (0.5 mg per 10×106 cells; Immunotech, Marseille, France) were added and incubated for 10 min at 37°C. Cells bound to beads were captured with magnetic force. Unbound cells were collected for flow cytometry staining assay.
Flow cytometry staining assay
Cells to be evaluated were incubated with 9.1C3 or 3G4 at 4°C for 30 min followed by washing three times with FACS washing buffer (1% FCS and 0.1% NaN3 in PBS). Then cells were incubated with PE-conjugated goat anti-mouse IgG at 20 μg/mL. After washing three times, cells were analyzed using a flow cytometer (Becton Dickinson, San Jose, CA).
Antibody-induced redirected cytotoxicity assay for the effect of MAbs on cytotoxicity
To investigate the effect of 3G4 on cytotoxicity, antibody-induced redirected cytotoxicity assay (RCA) was performed according to the methods described previously.(26,30) Human peripheral blood mononuclear cells pre-incubated with irradiated Raji cells were used as effector cells and FcγR-positive mouse-derived mastocytoma P815 were used as target cells. P815 cells were loaded with 51Cr via incubating with Na251CrO4 (Amersham Pharmacia, Bucks, England) solution for 1 h at 37°C. Cells were then washed three times with PBS, resuspended in culture medium, and planted in triplicate in 96-well plates at 1×104 cells/well. The effector cells (106 cells in 100 μL PBS) were incubated with CD16 MAb, CD56 MAb, 9.1C3 MAb, or 3G4 MAb at 37°C for 30 min followed by one washing with RPMI 1640 medium containing 10% FCS. Then the effector cells were added to the 51Cr-loaded target cells in wells at indicated effector-target (E:T) ratios. After 4 h incubation in a CO2 incubator at 37°C, the quantity of 51Cr released into the culture supernatants was determined by liquid scintillation counter (LS6000, Beckman, Fullerton, CA) immediately and the specific lysis was estimated using the following formula:
where the experimental release was defined as mean counts per minute (cpm) of gamma radiation released by target cells in the presence of effector cells and/or MAb and the spontaneous release was defined as the mean cpm released by target cells alone.
Effect of MAb 3G4 on binding of collagens to LAIR-1
To analyze the interference of 3G4 on the binding of collagens to LAIR-1, LAIR-1-transfected K562 cells were generated. The binding analysis was performed according to the previously published protocol with a minor modification.(13) Briefly, 96-well flat-bottom plates (Nunc) were coated overnight at 4°C with purified collagens I and III (Sigma) or bovine serum albumin (BSA) in PBS, respectively, supplemented with 2 mM acetic acid. After washing twice, wells were blocked with BSA. K562 stably transfected with LAIR-1 were fluorescently labeled with 5 μM calceine AM (Invitrogen) in PBS for 30 min at 37°C. After washing twice with RPMI 1640 containing 1% FCS, cells were added to each well and plates were incubated at 37°C for 2 h. For the analysis of the interferential effect of 3G4, cells were preincubated with 3G4 for 30 min at room temperature. After four washings, their capacity to adhere to the collagens in the 96-well plates was determined by using a Fluoroskan Ascent fluorescent spectrophotometer (Thermo Labsystems, Franklin, MA).
Statistical analysis
All data were derived from at least three independent experiments. The statistical significance was analyzed using the Mann-Whitney test. Data were represented as mean±SEM. A value of p<0.05 was regarded as significant.
Results
Expression and purification of LAIR-2-GST fusion protein
For expression of LAIR-2-GST fusion protein, a DNA fragment of LAIR-2 with GST tag was cloned into the expression vector pGEX-4T-3. After induction by IPTG, a protein with an apparent molecular mass of 43 kDa was overexpressed in E. coli BL21, which was consistent with the relative molecular weight (Mr) of the LAIR-2-GST fusion protein. The optimal conditions for LAIR-2-GST expression was culturing at 30°C and inducment by the addition of IPTG to a final concentration of 50 nM 2h before harvest. Western blotting with anti-GST MAb confirmed that this overexpressed band was LAIR-2-GST (data not shown). The LAIR-2-myc-6×His fusion protein was also successfully expressed in COS7 cells (data not shown).
Generation and characterization of MAbs to LAIR-2
After fusion of spleen cells from the LAIR-2-GST-immunized BALB/c mouse and myeloma cells, four hybridomas with high affinity to LAIR-2-myc-6×His were obtained from these clones using LAIR-2-myc-6×His fusion protein by indirect ELISA and were designated as 1A7, 3G4, 3G12, and 4A9. The titers of the ascitic fluid of the four hybridomas were all 1:106 determined by indirect ELISA. Immunoglobulin class and subclass assay revealed that 3G4 was IgG1. Specificity of these MAbs for LAIR-2 was evaluated by their binding to LAIR-2-myc-6×His and LAIR-1-Fc. Western blot analysis showed that 1A7, 3G12, and 4A9 were all LAIR-2 specific. However, MAb 3G4 could react with both LAIR-2 and LAIR-1 fusion proteins (Fig. 1A).
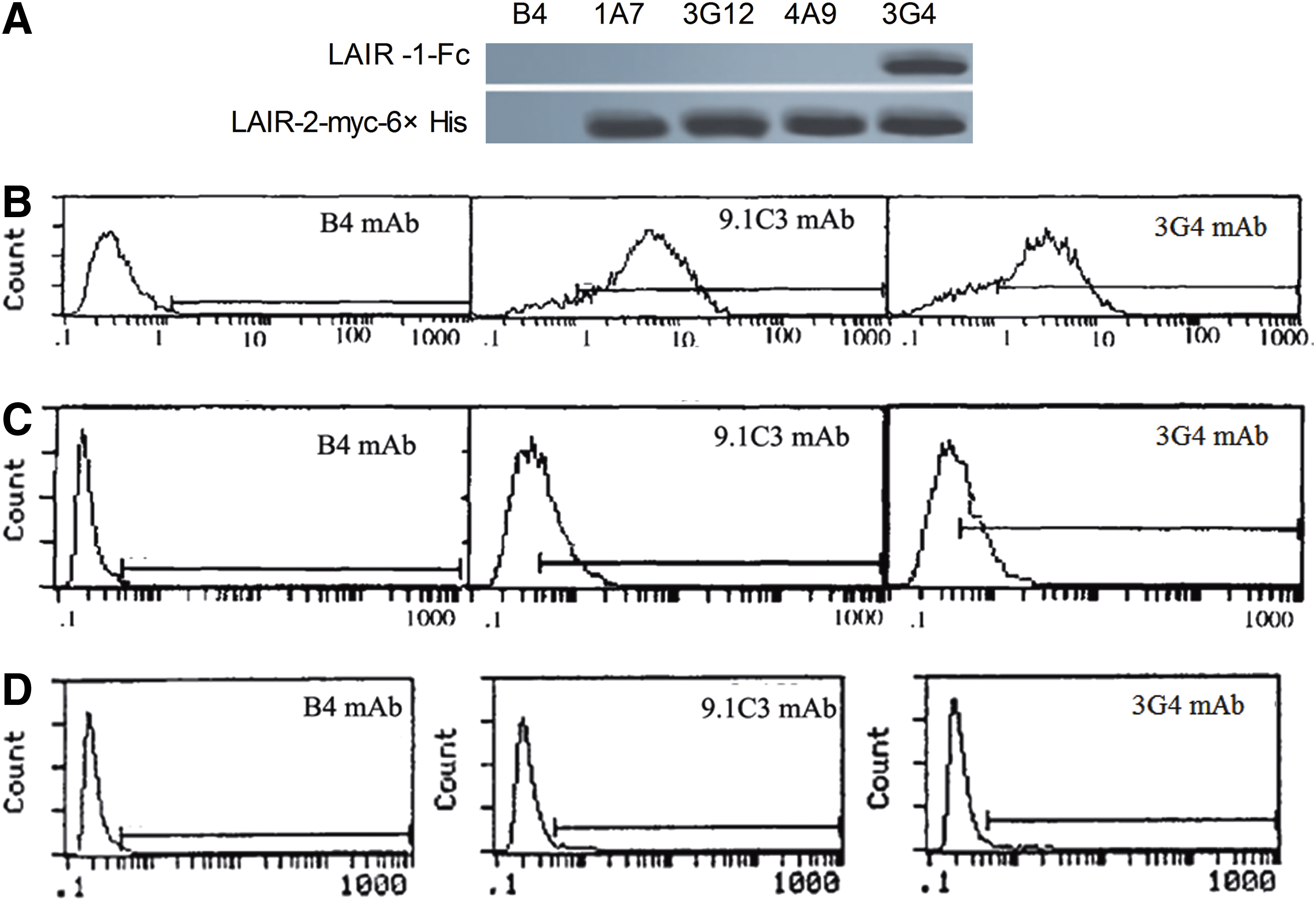
Characterization of MAbs to LAIR-2. (
Further study by flow cytometry showed that 3G4 could stain both PBMCs from healthy donors and Jurkat cells with similar peaks to that of 9.1C3 (Fig. 1B,C). When the LAIR-1 positive Jurkat cells were eliminated using immunomagnetic beads with 9.1C3, the residual Jurkat cells demonstrated both 9.1C3 and 3G4 negative (Fig. 1D), suggesting that 3G4 and 9.1C3 recognize the same molecule on Jurkat cells.
MAb 3G4 demonstrated no inhibition of lymphocyte killing ability
It was previously shown that cross-linking of LAIR-1 on human NK cells or T cells delivers a potent inhibitory signal that is capable of inhibiting target cell lysis.(4,31) In the current study, the potential role played by 3G4 in cytotoxicity was identified with RCA, which was a test to identify the function of a receptor on the cell surface with its specific MAb. CD16 and CD56 receptors were known as molecular markers of NK cells. CD16 was well known as a lysis receptor mediating direct NK cell cytotoxicity.(32) As shown in Figure 2, CD16 MAb administration significantly enhanced the cytotoxicity of PBMCs, while CD56 MAb exhibited no significant effect on the cytotoxicity. The 9.1C3 led to down-regulation of the activation effector cells as expected, even in the presence of CD16 MAb. However, 3G4, as another MAb to LAIR-1, showed an unanticipated result by exhibiting no effect on cytotoxicity although it could also bind to LAIR-1 as 9.1C3 MAb. This indicated that 3G4 binding did not activate LAIR-1 signaling.
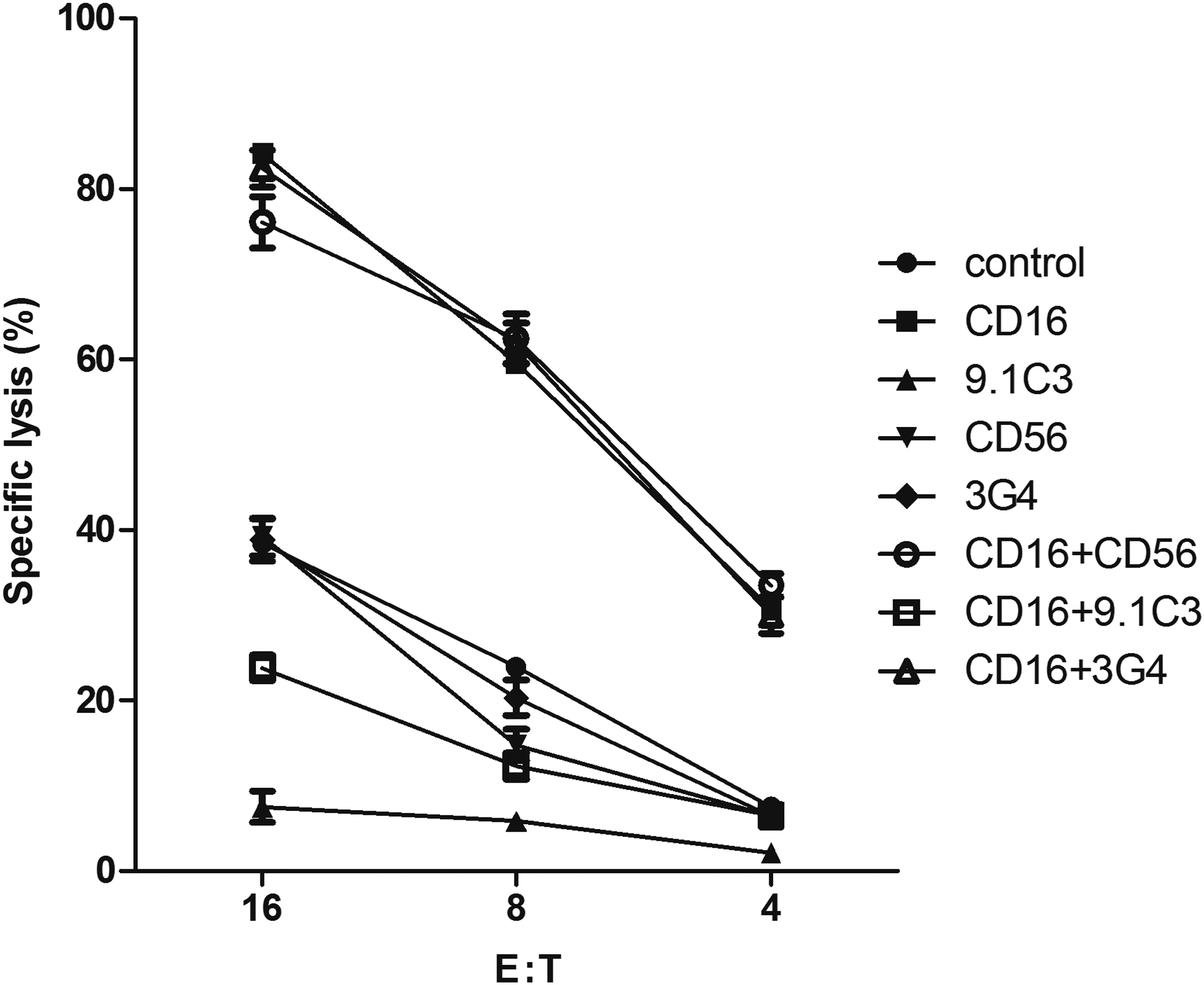
Effect of 3G4 on killing ability of effector cells was determined by RCA. The effector cells were incubated with various MAbs to be evaluated prior to co-incubating with 51Cr-loaded target cells. Specific lysis was assayed by detecting 51Cr release from P815 target cells. Data were represented as mean±SEM (n=3). The 3G4 demonstrated no inhibition to the cytotoxicity.
MAb 3G4 mildly prevented binding of LAIR-1 to collagen
As documented by Meyaard and colleagues,(13) the K562 cells stably transfected with human LAIR-1 could bind firmly to plate-bound collagens I and III. To further investigate whether 3G4 treatment disturbs the interaction between LAIR-1 and its ligand, human LAIR-1-transfected K562 cells were employed in this study. As shown in Figure 3, preincubation of LAIR-1-transfected K562 cells with 3G4 did not fully block the interaction of LAIR-1 with collagens, but only mildly reduced the percentage of cells binding to collagen I or III in a dose-dependent manner. This indicated that the 3G4 treatment partly prevented the binding of LAIR-1 to collagen.
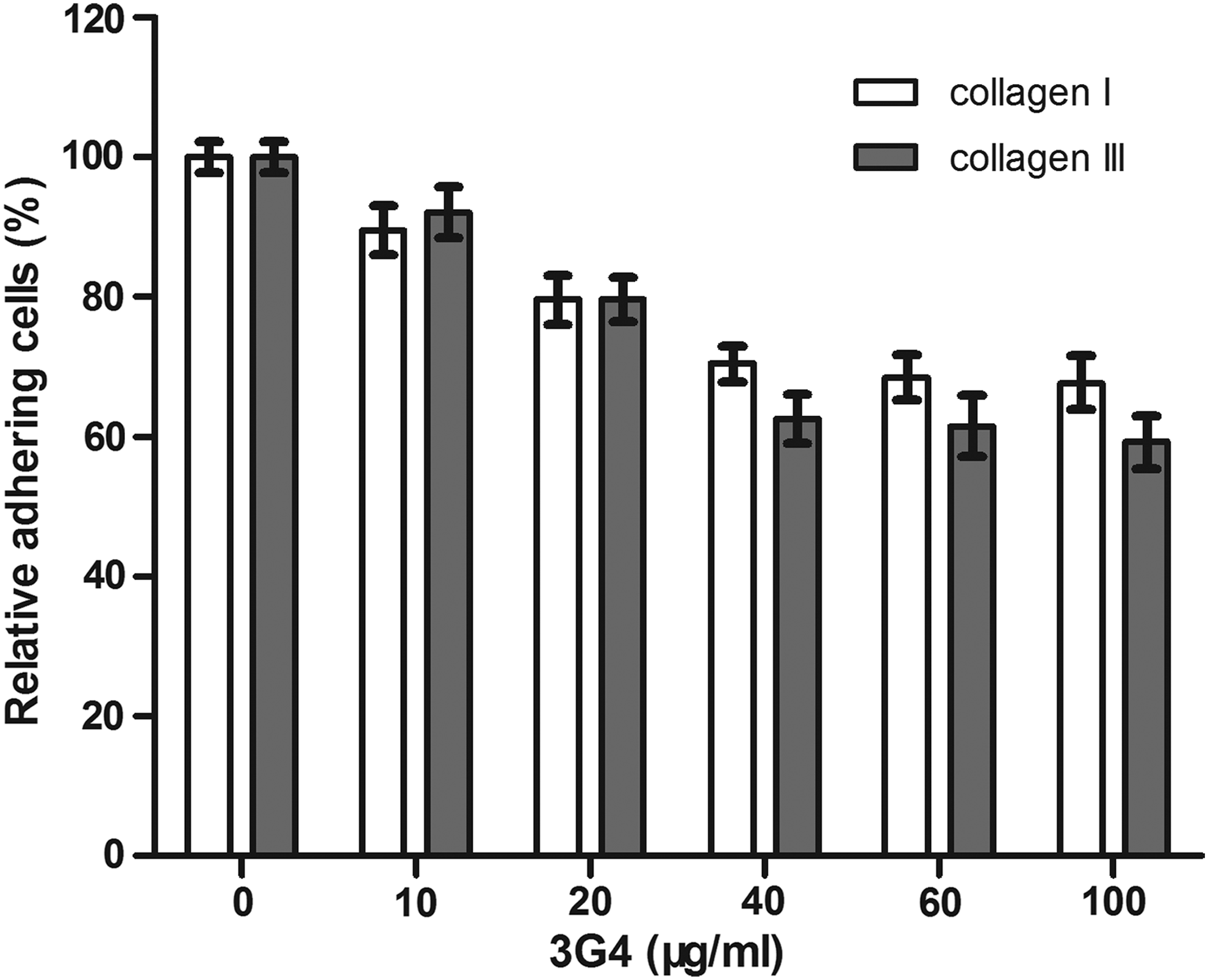
The 3G4 mildly prevented the binding of LAIR-1 to collagen. 96-well plates were coated with collagens I (
Discussion
Inhibitory signals help to attenuate or terminate the unwanted immune response triggered by activating ones in order to maintain an adequate immune response.(14,33,34) LAIR-1, an ITIM-bearing inhibitory receptor, was widely expressed on most immune cells. In vitro, cross-linking of LAIR-1 with MAb could deliver a potent inhibitory signal,(4) which was capable of downregulating activating signals. However, the exact role of LAIR-1 in vivo remains largely a mystery. In the present study, we generated four MAbs to LAIR-2 from LAIR-2-GST-immunized BALB/c mice. One of the four MAbs, the one designated 3G4, could react with LAIR-1 and LAIR-2, verifying that there was at least one common epitope in LAIR-1 and LAIR-2 molecules that could be processed by antigen-processing cells. Usually, the specific antibody is an effective tool to study the function of a receptor. In the RCA test, the fab region of a MAb bound to the receptor to be evaluated on the NK cell, and the Fc portion of the MAb bound to the Fc receptor on the target cell. When the inhibitory receptors were cross-linked by the corresponding MAb, the cytotoxicity would be inhibited. Likewise, the cytotoxicity would be enhanced by the cross linkage of activated receptors. In MAbs to LAIR-1 prepared previously, such as 9.1C3, once bound to the LAIR-1 protein, LAIR-1 would be activated and thus lead to downregulation of cell activation. Therefore, when purified using these MAbs, the LAIR-1-positive cells we collected were activated by the MAbs. Distinct from 9.1C3, MAb 3G4 did not deliver inhibitory signals in the RCA, and 3G4 partly inhibited the binding of LAIR-1-transfected K562 cells to plate-bound collagens. Although the epitope of 3G4 recognized on LAIR-1 awaits further study, we speculate that it might be a non-functional epitope in the interaction with collagens.
In conclusion, we briefly outlined the generation and characterization of MAbs to LAIR-2 in this study. Particular emphasis was given to the MAb 3G4, which could cross-react with LAIR-1. When binding to LAIR-1, MAb 3G4 did not mediate inhibitory signals. This feature of 3G4 allows us to develop a method to isolate LAIR-1-positive cells without changing their status, which may pave a way for the functional study of LAIR-1-positive cells.
Footnotes
Author Disclosure Statement
The authors have no financial interests to disclose.