
Abstract
Diacylglycerol kinase (DGK) is responsible for the enzymatic conversion of diacylglycerol to phosphatidic acid. Since both diacylglycerol and phosphatidic acid serve as signaling molecules, DGK is regarded as a hub between diacylglycerol-mediated and phosphatidic acid-mediated signaling. One of the 10 DGK isozymes, DGKα, is shown to be involved in T cell function. Transfection studies using tagged expression vectors revealed that DGKα localizes to the cytoplasm and nucleus and translocates to the plasma membrane in response to T cell receptor stimulation. However, a limited number of studies reported the localization of native protein of DGKα in tissues and cells. In this study, we immunized mice with recombinant DGKα and developed several anti-DGKα monoclonal antibodies (mAbs). One of the established anti-DGKα mAbs is a clone DaMab-2 (mouse IgG1, kappa). In enzyme-linked immunosorbent assay, DaMab-2 recognized only DGKα, and did not react with the other isozymes, such as DGKγ, DGKζ, DGKη, and DGKδ. Importantly, DaMab-2 is very useful in immunocytochemical analysis of human cultured cells, indicating that DaMab-2 is advantageous to analyze the localization and function of DGKα.
Introduction
D
DGK constitutes an enzyme family composed of 10 isozymes in mammalian species.(1–3) Each isozyme has a distinct molecular structure and subcellular localization pattern. Of DGKs, 80-kDa DGKα is the first identified enzyme, which contains EF-hand motif (a Ca2+-binding site), Zn-finger (C1 domain, diacylglycerol-binding site), and catalytic domain. A wide range of cell types and tissues express DGKα, including lymphocytes, glial cells, vascular cells, hepatocytes, adrenal medullary cells, fibroblasts, sarcoma cells, and melanoma cells.(3–11)
Functional significance of DGKα has been intensely investigated in T lymphocytes. In T cells, DGKα regulates cell proliferation in response to IL-2 stimulation(3) and is involved in T cell receptor (TCR) signaling through modulating RasGRP activity.(12) T cells isolated from DGKα-deficient mice show altered activity of TCR signaling and hyperproliferation.(13) Transfection studies in T cells demonstrated that DGKα localizes in the cytoplasm and translocates to the plasma membrane in response to the external stimuli.(14)
A limited number of studies have been reported about the localization of native protein using specific monoclonal antibodies (mAbs) because no mAb is available to detect human DGKα by immunostaining. Herein, we report a novel anti-human DGKα mAb, DaMab-2, which is very useful in immunocytochemical analysis of cultured cells.
Materials and Methods
Cell lines
P3 U1 and HeLa were obtained from the American Type Culture Collection (Manassas, VA). P3 U1 was cultured in the RPMI 1640 medium (Nacalai Tesque, Inc., Kyoto, Japan), supplemented with 10% heat-inactivated fetal bovine serum (FBS) (Thermo Fisher Scientific, Inc., Waltham, MA) at 37°C in a humidified atmosphere of 5% CO2 and 95% air. HeLa was cultured in DMEM (Wako Pure Chemical Industries Ltd., Osaka, Japan) supplemented with 10% FBS. Antibiotics, including 100 U/mL of penicillin and 100 μg/mL of streptomycin were added to media.
Plasmid preparation
Human DGKα cDNA(15) was synthesized, and subcloned into an expression vector, pMAL-c2 (New England Biolabs, Inc., Beverly, MA), with PA tag (GVAMPGAEDDVV)(16–18) using the In-Fusion PCR Cloning Kit, named pMAL-c2-DGKα-PA. DGKα with PA tag was also subcloned into an expression vector, pET41a(+) (Novagen; EMD Millipore Corp., Billerica, MA), with PA tag of N-terminus and RAP tag (DMVNPGLEDRIE)(19) of C-terminus, named pET41a-PA-DGKα-RAP. This construct was verified by direct DNA sequencing.
Production of recombinant protein of DGKα
Competent Escherichia coli TOP-10 cells (Thermo Fisher Scientific, Inc.) were transformed with the plasmid pMAL-c2-DGKα-PA. They were then cultured overnight at 37°C in the LB medium (Thermo Fisher Scientific, Inc.) containing 100 μg/mL ampicillin (Sigma-Aldrich Corp., St. Louis, MO). Cell pellets were resuspended in phosphate-buffered saline (PBS) with 1% Triton X-100 and 50 μg/mL aprotinin (Sigma-Aldrich Corp.). After sonication, the crude extracts were collected by centrifugation (9000 g, 30 min, 4°C). The recombinant PA-DGKα-RAP was expressed in transformed E. coli BL21 (DE3) cells that were cultured for 3 h at 37°C in the presence of 1 mM of IPTG, and the bacterial pellet from a 400 mL culture was suspended in 10 mL TBS (pH 7.5). Bacteria were lysed by sonication, and the soluble fraction was obtained by centrifugation at 15,000 g for 15 min.
The lysates were passed through a 0.45-μm filter to remove any trace amounts of insoluble materials. Cleared lysates were mixed with NZ-1-Sepharose (1 mL bed volume) and incubated at 4°C for 2 h under gentle agitation. The resin was then transferred to a column and washed with 20 mL TBS (pH 7.5). The bound protein was eluted with PA tag peptide at room temperature in a stepwise manner (1 mL × 10).
Hybridoma production
Female BALB/c mice (4 weeks old) were purchased from SLC Japan (Shizuoka, Japan). Animals were housed under pathogen-free conditions. The Animal Care and Use Committee of Tohoku University approved the animal experiments described herein. BALB/c mice were immunized using an intraperitoneal (i.p.) injection of MBP-DGKα-PA (100 μg) together with Imject Alum (Thermo Fisher Scientific, Inc.). After three additional immunizations, a booster injection was administered through i.p. two days before the spleen cells were harvested. The spleen cells were fused with P3 U1 cells using PEG1500 (Roche Diagnostics, Indianapolis, IN). The fused cells were grown in the RPMI medium with hypoxanthine, aminopterin, and thymidine selection medium supplement (Thermo Fisher Scientific, Inc.). Culture supernatants were screened using enzyme-linked immunosorbent assay (ELISA) for binding to recombinant MBP-DGKα-PA and PA-DGKα-RAP.
Enzyme-linked immunosorbent assay
Recombinant MBP-DGKα-PA and PA-DGKα-RAP were immobilized on Nunc MaxiSorp 96-well immunoplates (Thermo Fisher Scientific, Inc.) at a concentration of 1 μg/mL for 30 min. After blocking with 1% bovine serum albumin in 0.05% Tween20/PBS, the plates were incubated with culture supernatant followed by 1:3000-diluted peroxidase-conjugated anti-mouse IgG (Dako, Glostrup, Denmark). The enzymatic reaction was performed with a 1-Step Ultra TMB-ELISA (Thermo Fisher Scientific, Inc.). The optical density was measured at 655 nm using an iMark microplate reader (Bio-Rad Laboratories, Inc., Hercules, CA).
Transfection of siRNA against human DGKα
Silencing RNA duplexes directed against human DGKα (siDGKα1, 5′-UUAGGAGGUUGAACACCUGUCGAGG-3′ and siDGKα2 5′-CCUGGCCUCUGGACCGGAUCGUAAA-3′) were purchased from Thermo Fisher Scientific, Inc. Scrambled siRNA duplexes (AllStars Negative Control siRNA) (Qiagen, Valencia, CA) were used as control. HeLa cells were transfected with siRNAs against human DGKα or scrambled control using Lipofectamine RNAi MAX (Thermo Fisher Scientific, Inc.) according to the manufacturer's instruction. After 48 h of transfection, cells were fixed and stained by DaMab-2 as described above.
Immunocytochemical analysis
HeLa cells were fixed with 2% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) for 20 min on ice. Cells were perforated with 0.3% Triton-X 100/PBS for 20 min at room temperature, followed by treatment with 5% normal goat serum in PBS (NGS/PBS) to block nonspecific binding sites, and were incubated with 20 μg/mL of DaMab-2 or control (NGS/PBS) overnight at room temperature in a moist chamber. They were incubated with goat anti-mouse IgG-Alexa 488 (dilution 1:300; Thermo Fisher Scientific, Inc.) for 30 min at room temperature. Cells were also treated with DAPI (Thermo Fisher Scientific, Inc.) to stain cell nuclei. They were examined using confocal laser scanning microscopy (LSM700; Carl Zeiss, Inc., Jena, Germany).
Results
The recombinant MBP-DGKα-PA was captured using NZ-1-Sepharose and efficiently eluted with PA tag peptide.(16–18) Mice were then immunized with MBP-DGKα-PA to develop novel anti-DGKα mAbs. Using ELISA, the culture supernatants of hybridomas were screened for binding to recombinant MBP-DGKα-PA or PA-DGKα-RAP that was purified from E. coli. As a result, DaMab-2 (mouse IgG1, kappa) was established after limiting dilution. Western blot analysis demonstrated that DaMab-2 detected MBP-DGKα-PA, but not MBP-IDH1,(20–22) indicating that DaMab-2 reacts with DGKα, but does not react with MBP tag in western blot analysis (Supplementary Fig. S1). ELISA demonstrated that DaMab-2 reacted only with DGKα, and did not react with the other isozymes, such as DGKγ, DGKζ, DGKη, and DGKδ (data not shown).
Immunocytochemical analyses revealed that DaMab-2 reacted with HeLa cells (Fig. 1). The enlarged view clearly demonstrated a diffuse staining pattern in both the nucleus and cytoplasm. This pattern of staining was identical to the previous studies using rabbit polyclonal antibody on rat aortic smooth muscle and endothelial cells.(8,9) In addition, DaMab-2 reveals faint immunoreactions in the plasma membrane. Previous cDNA transfection studies described that DGKα translocates to the plasma membrane in response to TCR stimulation.(14) Negative control (without primary antibody) showed an absence of immunoreactions, suggesting that images obtained from DaMab-2 antibody were authentic.
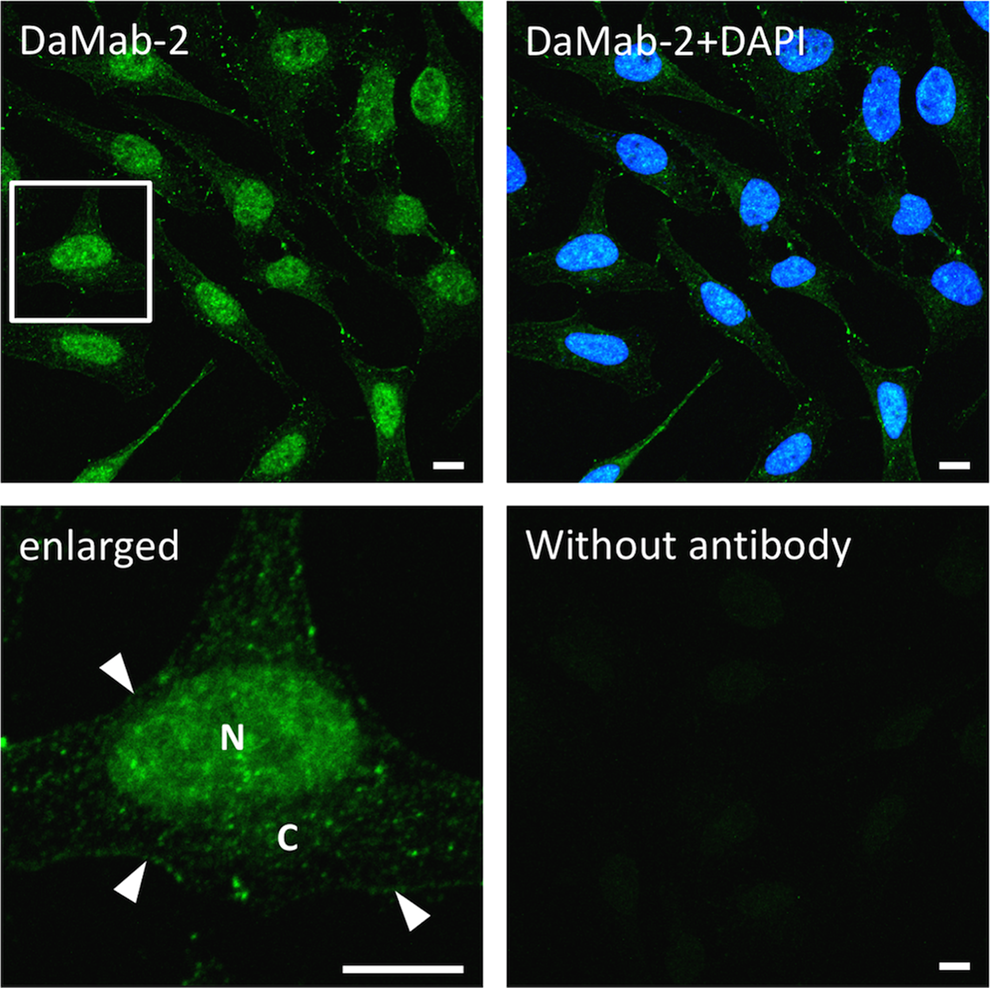
Immunocytochemical analysis using DaMab-2. HeLa cells were stained with DaMab-2 at a concentration of 20 μg/mL. Nuclear staining was performed with DAPI. As a negative control, cells were stained without primary antibody (lower right). Enlarged view of the square is shown on the lower left panel. N; Nucleus, C; Cytoplasm, arrowheads; plasma membrane. Bar = 10 μm.
To further confirm the accuracy of the immunoreactions of DaMab-2, we performed siRNA silencing experiments. As shown in Figure 2, transfection with siDGKα1 significantly reduced the immunoreactivity compared with the transfected scramble control siRNA (siCont). Similar results were obtained in the experiment using siDGKα2 (data not shown). These results confirm that DaMab-2 reacts with DGKα specifically and correctly in human cell line HeLa cells.
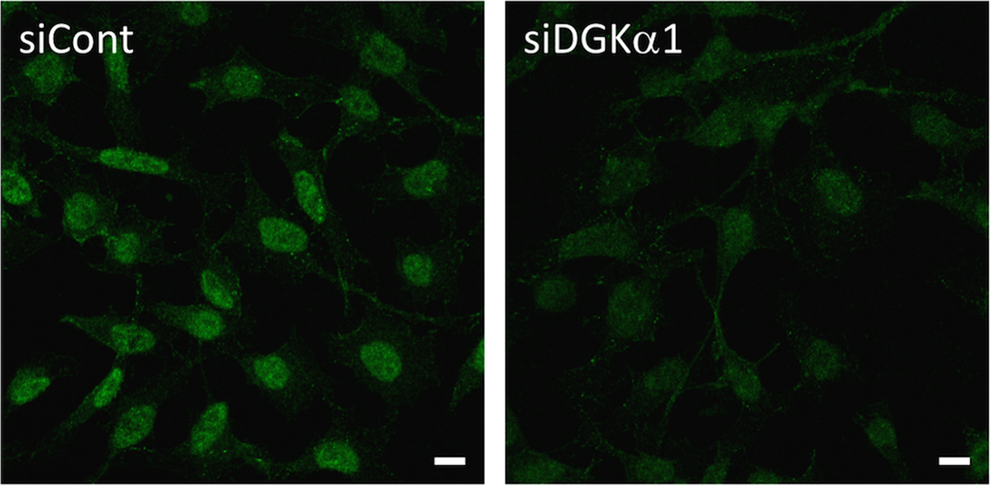
Knockdown of DGKα using siRNA. DaMab-2 immunocytochemistry was performed after siRNA treatment for human DGKα (siDGKα1). AllStars Negative Control siRNA (siCont) was used as a control. Bar = 10 μm.
Discussion
DGKα protein was purified from porcine brain in 1983,(23) followed by cDNA cloning from porcine,(24) human,(15) and rat brains.(4) In the rat study, a rabbit polyclonal antibody against rat DGKα was generated to determine DGKα-expressing cells in the rat brain. Fibrous and dot-like immunoreactions were detected in white matter by the polyclonal antibody, suggestive of an oligodendrocyte pattern. Using this polyclonal antibody, DGKα was shown to localize diffusely in the cytoplasm of hepatocytes and medullary chromaffin cells,(10,11) and in both the nucleus and cytoplasm of vascular cells(8,9) of rat tissues. However, highly specific and sensitive mAbs against human DGKα remain to be established.
Herein, we immunized mice with recombinant DGKα and successfully developed a clone DaMab-2 (mouse IgG1, kappa). ELISA demonstrated that DaMab-2 recognized only DGKα, and did not react with the other isozymes, such as DGKγ, DGKζ, DGKη, and DGKδ (data not shown). Furthermore, DaMab-2 is very useful in immunocytochemical analysis of HeLa cells (Fig. 1). Unfortunately, DaMab-2 did not react with endogenous DGKα of HeLa cells in western blot (data not shown). In this study, we focused on the development of mAbs against DGKα for immunocytochemistry. It might often be difficult to establish mAbs, which are useful for both western blot analysis and immunocytochemical analysis. The design of immunogen and screening methods to produce specific mAbs are totally different when we aim to develop mAbs for western blot or immunocytochemical analysis. In particular, it is very difficult to produce specific and sensitive mAbs to detect endogenous proteins in every experiment. Indeed, DaMab-2 can detect recombinant DaMab-2 in western blot (Supplementary Fig. S1), although it did not react with endogenous DGKα of HeLa cells in western blot. Several anti-DGKα mAbs are commercially available, and have been reported to be useful in western blot analysis and immunocytochemical analysis. However, recombinant or overexpressed DGKα are usually used in those studies. We need to further develop clones for detecting endogenous DGKα in western blot analysis.
In conclusion, a novel anti-DGKα mAb, DaMab-2, sensitively and specifically reacted with human DGKα using immunocytochemical analyses. DaMab-2 could be advantageous to analyze the localization and function of DGKα.
Footnotes
Acknowledgments
This work was supported in part by the Platform for Drug Discovery, Informatics, and Structural Life Science (PDIS) from Japan Agency for Medical Research and Development, AMED (Y.K.), by the Regional Innovation Strategy Support Program from the Ministry of Education, Culture, Sports, Science, and Technology (MEXT) of Japan (Y.K.), and by the Grant-in-Aid for Challenging Exploratory Research from MEXT (K.G.).
Author Disclosure Statement
Fumio Sakane, Akinobu Taketomi, Kaoru Goto, and Yukinari Kato received research funding from Ono Pharmaceutical Co., Ltd. The other authors have no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.