
Abstract
CD271 is a common receptor for all neurotrophins that is localized to neurons, endothelial cells, and the basal layer of the epithelium in normal tissue. Recently, we and others reported that CD271 plays essential roles in the development of squamous cell carcinoma, especially in tumor-initiating cells. Since little is known about how CD271 regulates cancer cell initiation and proliferation, antibodies that recognize different domains of CD271 are needed to enable investigation. Therefore, this study aimed to develop an antihuman CD271 antibody by immunizing mice with a CD271 antigen produced by a baculovirus. The antibody was named hCD271mAb#13, and it recognized cysteine-rich domain 1 with a higher affinity than the commercially available antibody ME20.4. We determined that hCD271mAb#13 is suitable for flow cytometry, Western blotting, immunocytochemistry, and immunohistochemistry of formalin-fixed paraffin-embedded tissue. Use of hCD271mAb#13 for CD271 labeling could enable detailed analyses of cancer cell regulation and other biological processes.
Introduction
CD271
CD271 is required for neural and vascular development(2) and is also recognized as a neural crest stem cell marker.(3) CD271 was recently identified as a tumor-initiating cell marker in human melanoma.(4) We previously reported that CD271 is important for tumor development by tumor-initiating cells in squamous cell carcinoma. We demonstrated that CD271 is expressed in proliferative cells of hypopharyngeal cancer and lung squamous cell carcinoma,(5–7) and localized to the basal layer of the normal epithelium. CD271 knockdown caused almost complete suppression of cell proliferation.(5,7) We also recently reported that humanized anti-CD271 antibodies exert antibody-dependent cellular cytotoxicity activity against hypopharyngeal cancer and melanoma cells using a xenografted mouse model.(8) These data suggest that CD271 is a potential therapeutic target for various cancers. Because the mechanism of CD271-mediated regulation of cancer cell proliferation is not fully understood, antibodies that recognize a specific domain of CD271 are needed. Therefore, we aimed to develop a novel antihuman CD271 antibody through this study.
Materials and Methods
Purification of CD271 ECD recombinant protein
Purification of recombinant protein was performed as described previously.(8) In brief, Escherichia coli DH10Bac (Invitrogen, Carlsbad, CA) was transformed using a pFastBac-CD271 ECD recombinant donor plasmid. To produce recombinant baculovirus, precultured Sf21 cells were transfected using recombinant bacmid DNA and Cellfection II reagent (Invitrogen) according to the manufacturer's instructions. A lysis buffer (20 mM Tris-HCl, 10% glycerol, 1% NP-40, 1 mM ethylene glycol tetraacetic acid, 1 mM dithiothreitol (DTT), and 150 mM NaCl, pH 8.0) with a protease inhibitor cocktail (Sigma-Aldrich, St. Louis, MO) was added to collected cells, and the mixture was then sonicated and centrifugated. The supernatant was then mixed with glutathione sepharose 4B (Sigma-Aldrich) for 60 minutes at 4°C. PreScission Protease (GE Healthcare, IL) with PreScission Cleavage buffer (50 mM Tris-HCl, 150 mM NaCl, and 1 mM DTT, pH 7.5) was added to the mixture.
Establishment of hybridoma cell lines
BALB/c mice were immunized using subcutaneous injections of 40 μg of recombinant CD271-ECD together with complete adjuvant (Freund) (BD Biosciences, CA). After several additional immunizations, a booster injection was intravascularly administered 6 and 3 days before harvesting spleen cells. Spleen cells were fused with SP2/0 cells using an HVJ Envelope Cell Fusion Kit (GenomeOne-CF; ISHIHARA SANGYO KAISHA, Osaka, Japan) and grown in GIT medium (Wako, Osaka, Japan) supplemented with 10% fetal bovine serum (FBS), hypoxanthine, aminopterin, and thymidine selection medium supplement (Thermo Fisher Scientific, MA).
A culture supernatant of hybridoma cells was screened using enzyme linked immunosorbent assay (ELISA). HPCM2-CD271 and HPCM2-EV cells were seeded in plates and fixed in 4% paraformaldehyde. Fixed cells were incubated with supernatant and antimouse horseradish peroxidase-conjugated immunoglobulin G (IgG) was applied. Enzyme activity was detected using a tetramethyl benzidine substrate.
For epitope determination, transiently transfected 293T cells (CD271-full length, Δ1, Δ1–2, Δ1–3, and Δ1–4) were seeded in polyethylenimine-coated plates and incubated for 24 hours.(8) For determination of antibody concentration, plates were coated with antimouse IgG2a (ISO2-1 kit; Sigma-Aldrich), washed, and incubated with the culture supernatant of hybridoma cells or antimouse IgG2a antibody as a standard (200 μg/mL, anti-CA IX, H-11; Santa Cruz Biotechnology, TX). The antibody isotype in the hybridoma supernatant was determined using mouse monoclonal antibody isotyping reagents (Sigma-Aldrich) according to the manufacturer's protocol.
Cell lines
Two human hypopharyngeal patient-derived xenograft (PDX) cell lines (HPCM1 and HPCM2, maintained in RPMI-1640 medium [Wako, Japan] supplemented with 10% FBS and 100 U/mL penicillin, and 100 μg/mL streptomycin), the SK-OV-3 cell line (ovarian adenocarcinoma, maintained in Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% FBS and penicillin/streptomycin, purchased from American Tissue Culture Collection, VA), and the MeWo-Luc and SK-MEL-2-Luc cell lines (malignant melanoma, maintained in DMEM supplemented with 10% FBS and penicillin/streptomycin, provided by Japanese Collection of Research Bioresources Cell Bank, Osaka, Japan) were used. The MCC148c cell line was established by PDX of cancer tissue derived from a patient with lung squamous cell carcinoma(7) and maintained in DMEM (Wako) supplemented with 10% FBS, 0.4 μg/mL hydrocortisone, 2.5 μM Y-27632 (Focus Biomolecules, PA), and penicillin/streptomycin.
Flow cytometry and Western blotting
Flow cytometry was performed as described previously.(5) Fluorescence data were collected using a cell analyzer (SA3800; SONY, Tokyo, Japan). Western blotting was performed as described previously.(5)
Small interfering RNAs
CD271 small interfering RNAs (siRNAs) #1 (siCD271#1, HSS107179) and #2 (siCD271#2, HSS107181), and nonsilencing control siRNA (siControl, 12935-300) were purchased from Invitrogen. siRNA transfections were performed using Lipofectamine® RNAiMAX Reagent (Life Technologies, CA) in antibiotic-free medium for 48 hours. The siRNA knockdown efficiencies were confirmed using flow cytometry analysis.
Establishment of human CD271-overexpressing cells
Human CD271-overexpressing cells (SP2/0-hCD271 and SK-OV-3-hCD271) were established using the retroviral vector pBABE-puro-CD271, as described previously.(8)
Immunohistochemical staining
Immunostaining of human CD271 was performed as previously described.(5) The anti-CD271 antibody (C40-1457) was purchased from BD Biosciences (Franklin Lakes, NJ). Cell blocks were prepared as follows: a piece of paraffin was set at the bottom of a 1.5-mL tube to which cells were added. The tube was centrifuged and the supernatant formed was removed; agar was, then, added to the tube. A 10% formaldehyde solution was, then, added to the cells for 24 hours at room temperature to fix them. Next, the blocks containing the cells were processed to paraffin blocks.
Results and Discussion
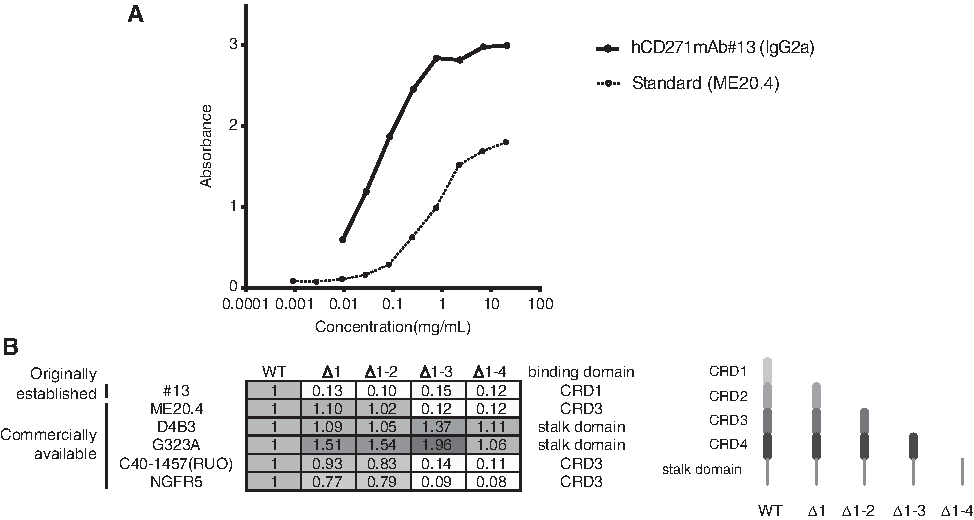
To develop a novel anti-CD271 mAb, we immunized mice with purified CD271 ECD protein. Culture supernatants were screened using ELISA for the ability to bind to hCD271-overexpressing HPCM2 cells. After limiting the dilution of the hybridomas, hCD271mAb#13 (IgG2a) was established. The dissociation curve of hCD271mAb#13 was analyzed, and we found that hCD271mAb#13 possessed higher affinity than the commercially available antibody ME20.4 (Fig. 1A). Next, we tried to determine which domain of CD271 was recognized by hCD271mAb#13 using ELISA and found that hCD271mAb#13 did not bind CRD1-deleted CD271, suggesting that the recognition domain of hCD271mAb#13 was specific for CRD1. The recognition domains of five other commercially available antibodies have specificity for CRD3 or the stalk domain, but not CRD1.
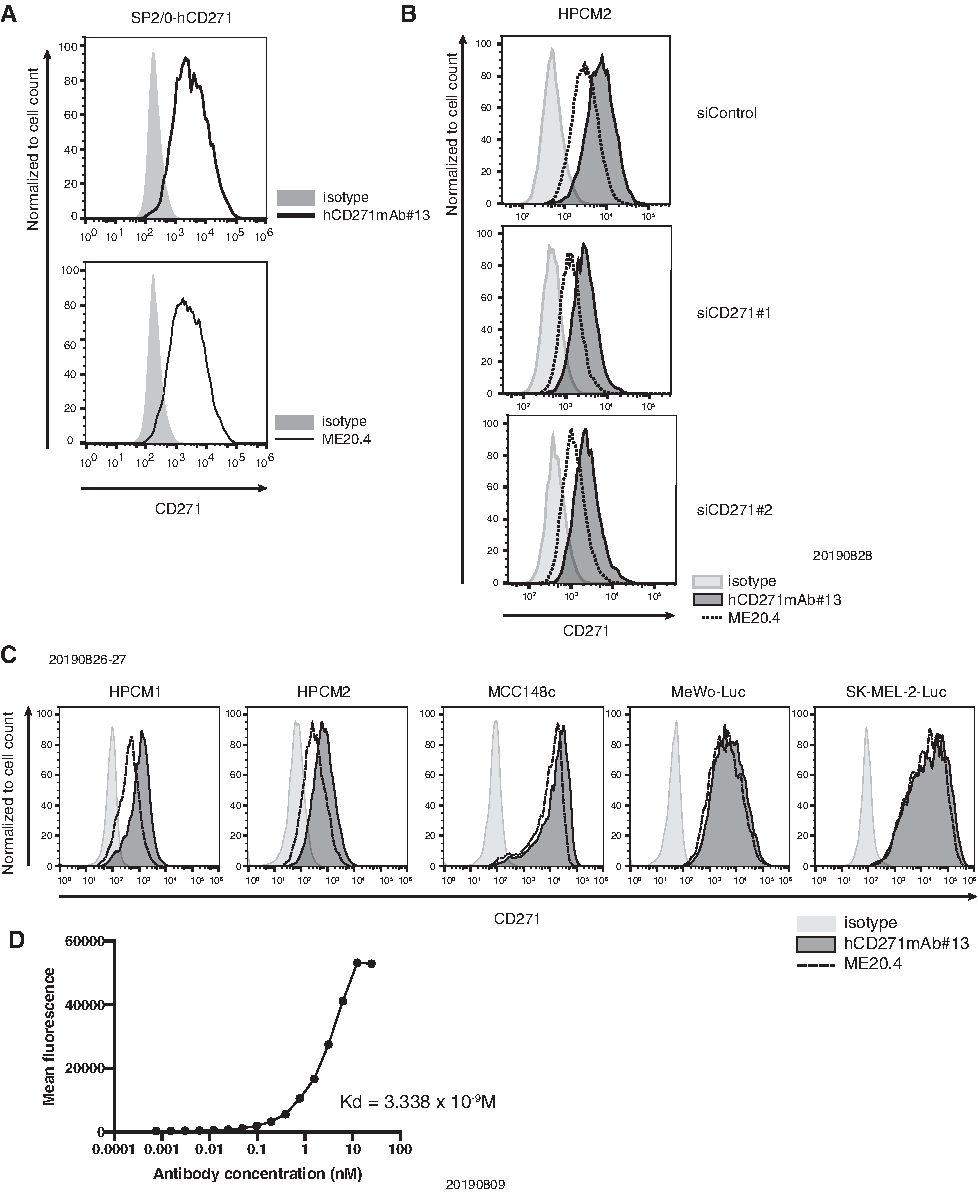
Next, we examined the affinity of hCD271mAb#13 for human CD271 using cell lines by flow cytometry. In human CD271-overexpressing SP2/0 cells, hCD271mAb#13 bound to SP2/0-CD271 cells (Fig. 2A). We also evaluated the affinity of hCD271mAb#13 in CD271 knockdown HPCM2 cells using flow cytometry. The intensity detected by flow cytometry was clearly diminished in CD271 knockdown HPCM2 cells compared with control cells (Fig. 2B). Thus, we concluded that hCD271mAb#13 specifically detects CD271. We also evaluated the affinity of hCD271mAb#13 for endogenously expressed CD271 in human cancer cell lines (hypopharyngeal cancer, melanoma, and lung squamous cell carcinoma). hCD271mAb#13 bound endogenous CD271, as did a commercially available antibody ME20.4, in several types of human cancer cell lines (Fig. 2C). Moreover, we analyzed the kinetics of hCD271mAb#13 binding to SK-OV3-CD271 cells using flow cytometry (Fig. 2D). The dissociation constant (KD) of hCD271mAb#13 was estimated to be 3.338 × 10−9 M, similar to that of antibodies used in clinical trials.(9) We also assessed the application of this antibody for laboratory techniques. In Western blotting, hCD271mAb#13 detected CD271 expressed by SK-OV-3-CD271 cells, as did a D4B3 clone, which is used as a commercially available antibody.
In immunocytochemistry experiments, hCD271mAb#13 detected overexpressed CD271, which was dominantly expressed on the cellular membrane (Fig. 3B).

We also determined hCD271mAb#13 applicability for immunohistochemistry. First, we stained blocks of SK-OV-3 and SK-OV-3-hCD271 cells and observed that hCD271mAb#13 detected CD271 in both cell types compared with an isotype control (Fig. 3C). Next, we stained surgical specimens of hypopharyngeal cancer using hCD271mAb#13 and found that it detected CD271-positive cancer cells, as did a commercially available antibody (C40-1457) (Fig. 3D). CD271 staining with hCD271mAb#13 showed protein localization near the stroma, consistent with our previous findings.(6)
Taken together, our findings demonstrate successful production of a monoclonal antibody specific for CD271, hCD271mAb#13. hCD271mAb#13 enabled the detection of the CRD1 domain of human CD271 using flow cytometry, Western blotting, immunocytochemistry, and immunohistochemistry. Application of hCD271mAb#13 for CD271 labeling could facilitate the study of cancer cell regulation and other biological processes.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported, in part, by Japan Society for the Promotion of Science (JSPS) KAKENHI Grant Nos. JP: 19K08430, 17K10716, 17K11166, 16K07132, 18K16432, and 19K09903 and Practical Research for Innovative Cancer Control from Japan Agency for Medical Research and Development (AMED) by Grant No. JP17ck0106197, The Cooperative Research Project Program of Joint Usage/Research Center at The Institute of Development, Aging and Cancer, Tohoku University, and Takeda Medical Foundation.